代謝工程改造畢赤酵母生產葡萄糖二酸
2018-11-06 11:15:14堵國成華兆哲
食品與生物技術學報 2018年9期
關鍵詞:產量
劉 葉 , 鞏 旭 , 康 振 , 堵國成 , 陳 堅 , 華兆哲 *
(1.江南大學 工業生物技術教育部重點實驗室,江蘇 無錫214122;2.江南大學 生物工程學院,江蘇 無錫214122)
葡萄糖二酸(Glucaric acid)是一種自然界存在的高附加值有機酸,在十字花科蔬菜、櫻桃和柑橘類水果中含量豐富,少量存在于包括人在內的哺乳動物中[1]。葡萄糖二酸及其衍生物具有廣泛的應用價值,它可以降低膽固醇[1]、治療糖尿病[2]、預防癌癥[3]等,同時,在合成生物可降解的聚合物、羥基化尼龍等新型材料方面也有大量的應用[4-5],曾被美國能源部確定為“最具價值的生物煉制產品”之一[6]。
目前,葡萄糖二酸的制備方法主要為化學法,如硝酸氧化、 以 2,2,6,6-四甲基哌啶氧化物(TEMPO)為催化劑的共催化氧化等[7]。此方法投入高、得率低,同時產生的廢氣廢液對環境造成一定的污染。由于化學法存在的局限性,生物法制備葡萄糖二酸越來越受到研究者的關注。葡萄糖二酸與抗壞血酸是哺乳動物中葡萄糖醛酸代謝途徑的終產物,然而,從葡萄糖到葡萄糖二酸,至少需要10步反應[8]。Prather等通過在大腸桿菌中異源表達釀酒酵母(Saccharomyces cerevisiae)來源的肌醇-1-磷酸合成酶基因(Ino1)、小鼠來源的肌醇氧化酶基因(MIOX)以及丁香假單胞菌(Pseudomonas syringae)來源的醛酸脫氫酶基因(udh),成功構建葡萄糖二酸合成途徑,實現了葡萄糖二酸的生物合成[9-11]。通過優化途徑、提高MIOX的穩定性及前體物質肌醇的轉運能力,在補加肌醇的條件下,葡萄糖二酸的產量達到了4.85 g/L。但是,大腸桿菌耐酸能力弱,易受噬菌體感染,并具有潛在的致病性,因此在工業上難以推廣應用。
畢赤酵母是一種單細胞真核生物,具有營養要求低、生長快、培養基廉價、可進行高密度發酵等優點[12]。相比大腸桿菌,畢赤酵母具有真核生物表達時對蛋白質的加工和修飾等功能[13],同時,畢赤酵母耐酸能力較強(pH 3~7)[14],因此是一種可用于生產功能性有機酸的優良宿主。
在本研究中,我們通過異源表達小鼠來源的MIOX基因及Pseudomonas putidaKT2440來源的udh基因,成功在畢赤酵母內構建了葡萄糖二酸合成途徑,通過對碳源、初始pH、接種齡及接種量進行搖瓶水平的優化,進一步提高了葡萄糖二酸的產量。最后,我們按照搖瓶優化的條件,對重組菌在3 L發酵罐上進行了補料分批培養的研究。
1 材料與方法
1.1 材料
1.1.1 菌種和質粒Pichia pastorisGS115:本研究中代謝工程改造的宿主菌株;Pseudomonas putidaKT2440:用于 udh基因的克隆;Escherichia coliJM109、E.coliTOP10:用于構建質粒的宿主菌;質粒pUC57-mM:含小鼠來源的MIOX基因,由南京金斯瑞生物科技有限公司合成,并按照畢赤酵母密碼子偏好性進行了優化;質粒pPIC9K、pGAPZB:用于重組質粒的構建。
1.1.2 培養基LB培養基(g/L):胰蛋白胨10,酵母粉 5,NaCl 10。制備固體培養基時添加2%的瓊脂。
YPD 培養基(g/L):蛋白胨 20,酵母粉 10,葡萄糖20。
YPDM 培養基(g/L):蛋白胨 20,酵母粉 10,葡萄糖 20,肌醇 10。
YPDS 培養基(g/L):蛋白胨 20,酵母粉 10,葡萄糖 20,山梨醇 182.2。
MD 固體培養基(g/L):葡萄糖 20,YNB 13.4,生物素 4×10-4,瓊脂 20。
改良 BSM 培養基 (g/L):85%磷酸 26.7 mL,CaSO40.93,K2SO418.2,MgSO47.275,KOH 4.125,PTM1離子液4.35 mL,葡萄糖60,肌醇13。
PTM1 離子液 (g/L):CuSO4·5H2O 6,KI 0.08,MnSO4·H2O 3,H3BO30.02,MoNa2O4·H2O 0.2,CoCl20.5,ZnCl220,FeSO4·7H2O 65,生物素 0.2;濃 H2SO45.0 mL。
1.1.3 酶、引物、DNA Marker及相關試劑盒Primer STAR DNA聚合酶,DNA Marker,T4 DNA連接酶,大腸桿菌感受態制備試劑盒:購自TaKaRa(大連);各種限制性內切酶:購自Thermo公司;PCR引物(見表1):由上海生工生物有限公司合成;質粒小量抽提試劑盒、細菌總DNA提取試劑盒:購自上海生工生物有限公司;酵母基因組提取試劑盒:購自天根生化科技(北京)有限公司;葡萄糖二酸鉀色譜級標準品:購自Sigma-Aldrich公司。
1.2 方法
1.2.1 重組表達載體的構建以pGAPZB質粒為模板,TU-1-F、TU-1-R為引物擴增畢赤酵母組成型啟動子TEF,同時以惡臭假單胞菌 (P.putidaKT2440)基因組為模板,TU-2-F、TU-2-R為引物擴增udh基因。使用融合PCR方法,將TEF啟動子與udh基因融合。使用限制性內切酶SacI和NotI對得到的融合片段雙酶切,連接至具有相應切口的表達載體pPIC9K上,轉化至E.coliJM109中,在確保閱讀框正確的前提下鑒定出重組表達質粒pPIC9KTEF-U,經DNA測序比對,重組序列正確。以pUC57-mM質粒為模板,mM-F、mM-R為引物擴增小鼠來源的MIOX基因,使用XhoI和XbaI連接至具有相應切口的表達載體pGAPZB上,轉化E.coliTOP10中,在確保閱讀框正確的前提下,鑒定出重組表達質粒pGAPZB-M,經測序比對,序列正確。

表1 本文所用的引物Table 1 Primers used in this study
1.2.2 畢赤酵母轉化方法及轉化子篩選將質粒pPIC9KTEF-U經SalI線性化之后,采用電轉化方式轉化至P.pastorisGS115感受態細胞中,在MD培養基中進行篩選,30℃恒溫倒置培養3~4 d。挑選MD平板上生長良好的單菌落分別接種至含有1.0、2.0、3.0、4.0 mg/mL G418的YPD平板中,篩選抗高質量濃度G418(4.0 mg/mL)的重組畢赤酵母,以重組子基因組DNA為模板進行PCR驗證,獲得重組菌株。將質粒pGAPZB-M經BspHI線性化,轉化上述重組菌感受態中,涂布于含100 μg/mL博萊霉素的YPDS平板上,30℃恒溫倒置培養3~4 d。挑選生長良好的單菌落接種至分別含有500、1 000、1 500、2 000 μg/mL博萊霉素的YPDS平板上,篩選抗高濃度博萊霉素(2 000 μg/mL)的重組子,以重組子基因組DNA為模板進行PCR驗證,獲得重組菌株。
1.2.3 重組畢赤酵母菌株培養條件
1)搖瓶培養:將重組菌單菌落接種至25 mL(搖瓶容量為250 mL)種子培養基YPD中,30℃、220 r/min培養24 h,將種子培養液按10%接種體積分數轉接至50 mL(搖瓶容量為500 mL)的發酵培養基YPDM中發酵,培養60 h。
2)3 L發酵罐培養:將重組菌單菌落接種至50 mL(搖瓶容量為 500 mL)YPD 培養基中,30℃、220 r/min培養24 h,將種子培養液按25%的接種體積分數轉接至含1 L改良BSM發酵培養基的3 L發酵罐中。使用80%氨水控制pH為5.5,溫度30℃,攪拌轉速為800 r/min,通氣量為4 vvm。當發酵液中葡萄糖質量濃度低于5 g/L時,流加葡萄糖維持質量濃度在0~5 g/L之間。
1.2.4 葡萄糖二酸的檢測
1)定性檢測:發酵結束后,取1 mL發酵液在12 000 r/min下離心5 min,取上清液經0.22 μm濾膜過濾,用于島津離子阱飛行時間質譜儀(LCMSIT-TOF)分析[15]。流動相:A:1 mmol/L 甲酸銨+0.01%甲酸+水;B:1 mmol/L 甲酸銨+乙腈;洗脫程序:0~12 min 30%B;12~30 min 30%~65%B;30~31 min 65%~95%B;31~35 min 95%B;35~36 min 95%~30%B;36~40 min 30%B; 色譜柱:Shim-pack VPODS(150 L×2.0);流速:0.15 mL/min 進樣量:5 μL;離子化模式:電噴霧負離子模式;霧化氣流速:1.5 L/min;CDL溫度:200℃,HB 溫度:200℃;掃描范圍:MS1,m/z:150~300。
2)定量檢測:取樣品發酵液上清液,經Affi-Gel凝膠處理[16]后,用高效液相色譜儀(HPLC)分析。流動相:5 mmol/L稀硫酸;色譜柱:Aminex HPX-87H;流速 0.5 mL/min;柱溫:55 ℃;進樣量:10 μL;檢測器:UV(檢測波長:210 nm)。
1.2.5 MIOX及Udh的活性測定發酵結束后,將發酵液在12 000 r/min、4℃下離心5 min,棄上清液。將菌體用Tris-HCl緩沖液(10 mmol/L,pH 8.0)清洗兩遍后,重懸在清洗緩沖液中。在重懸菌體加入1.0 mm硅珠,采用FastPrep-24均質破碎儀破碎,破壁后離心,上清液用來檢測途徑酶活性。MIOX及Udh的活性檢測方法見文獻[10]。
MIOX單位酶活定義:1 min內生成1 nmol/L醛酸所需要的酶量;Udh單位酶活定義:1 min內生成1 μmol/L NADH所需要的酶量。
2 結果與討論
2.1 重組表達載體的鑒定及重組菌的構建
將重組表達質粒pPIC9KTEF-U用SacI和NotI雙酶切,得到約8.3 kb與1.2 kb的兩條可見特異性DNA條帶,見圖1(a),分別對應于切除AOX啟動子的線性化pPIC9K和融合片段TEF-Udh。重組表達質粒pGAPZB-M經XhoI、XbaI雙酶切,得到約2.9 kb與 858 bp的兩條可見特異性DNA條帶,見圖1(b),分別對應于線性化的pGAPZB和目的基因MIOX。上述結果表明,重組表達質粒pPIC9KTEF-U及pGAPZB-M均構建成功。
將重組質粒pPIC9KTEF-U采用電轉的方式轉化P.pastorisGS115感受態細胞中,獲得重組菌,命名為P.pastorisGS115-TU,進一步將重組質粒pGAPZB-M轉化P.pastorisGS115-TU感受態細胞中,獲得相應的重組菌,命名為P.pastorisGS115-TUM。

圖1 雙酶切驗證重組質粒Fig.1 Confirmation of recombinant plasmids by digestion
2.2 重組畢赤酵母發酵生產葡萄糖二酸
將重組菌P.pastorisGS115-TUM及對照菌P.pastorisGS115在搖瓶中培養60 h后,取1 mL上清液進行LCMS-IT-TOF質譜分析,結果見圖2。在P.pastorisGS115-TUM發酵上清液中成功檢測到葡萄糖二酸(m/z=209),而對照菌P.pastorisGS115并未檢測到葡萄糖二酸。該結果表明,通過引入MIOX基因及udh基因,在重組菌P.pastorisGS115-TUM中已成功實現葡萄糖二酸的合成,而原始菌株并不具有生成葡萄糖二酸的能力。為了進一步分析途經,我們對途經酶MIOX及Udh的活性進行了檢測,結果見圖3。MIOX并未檢測到明顯的活性,推測原因為表達量較低或不穩定,但Udh酶活性較高,后面的研究工作將進一步分析并提高MIOX的活性。
經高效液相色譜(HPLC)定量檢測,重組菌P.pastorisGS115-TUM在500 mL三角搖瓶中培養60 h 后,葡萄糖二酸產量為(566.36±16.98)mg/L。

圖2 發酵液中葡萄糖二酸的質譜檢測Fig.2 Mass spectrum of glucaric acid in the culture supernatant

圖3MIOX及Udh酶活性檢測Fig.3 Specific activities of MIOX and Udh
2.3 搖瓶水平優化培養條件提高葡萄糖二酸產量
2.3.1 不同碳源對葡萄糖二酸產量的影響選擇甘油和葡萄糖作為碳源,分別以不同初始質量濃度(20、40、60、80、100 g/L)在 500 mL 搖瓶中培養重組菌P.pastorisGS115-TUM,結果見圖4。不同的碳源對菌體細胞生長及葡萄糖二酸的產量影響較大,與底物甘油相比,雖然以葡萄糖為碳源獲得的菌體量較低,但是葡萄糖二酸產量明顯高于甘油。由圖4可以看出,初始葡萄糖質量濃度為60 g/L時葡萄糖二酸產量最高,為(830.08±32.60) mg/L,因此選擇葡萄糖作為后期培養的碳源。

圖4 碳源對菌體生長及產量的影響Fig.4 Effect of carbon sources on recombinant strain and glucaric acid production
2.3.2 不同初始pH對葡萄糖二酸產量的影響配制 初 始 pH 分 別 為 4.0、4.5、5.0、5.5、6.0、6.5 的YPDM發酵培養基,初始葡萄糖質量濃度為60 g/L,將培養24 h的種子培養液以10%的接種體積分數轉接到發酵培養基中,培養60 h,結果見圖5。不同初始pH對菌體量影響不大。pH值為4.0~5.5時,葡萄糖二酸的產量逐漸增大;當pH大于5.5時,葡萄糖二酸產量略有下降。因此,選擇培養基最適初始pH為5.5。

圖5 初始pH對菌體生長及產量的影響Fig.5 Effect of initial pH on recombinant strain and glucaric acid production
2.3.3 不同接種齡對葡萄糖二酸產量的影響分別取培養 12、16、20、24、28、32 h 的種子培養液以10%的接種體積分數轉接到初始pH為5.5的發酵培養基中,發酵培養60 h,見圖6。在24 h之前,隨著接種齡的增加,葡萄糖二酸產量及菌體量逐漸增加,在24 h之后,菌體量變化不大,但是葡萄糖二酸產量有所下降。因此,選擇最適接種齡為24 h。
2.3.4 不同接種體積分數對葡萄糖二酸產量的影響取培養24 h的種子培養液分別以5%、10%、15%、20%、25%、30%的接種體積分數轉接至初始pH為5.5的發酵培養基中,發酵培養60 h,見圖7。在5%~25%范圍內,菌體量、葡萄糖二酸產量與接種體積分數成正相關關系;當大于25%時,菌體量、葡萄糖二酸產量均略有下降。因此,最優接種體積分數確定為25%。

圖6 接種齡對菌體生長及產量的影響Fig.6 Effect of seed time on recombinant strain and glucaric acid production

圖7 接種體積分數對菌體生長及產量的影響Fig.7 Effect of inoculation amount on recombinant strain and glucaric acid production
2.3.5 不同初始肌醇質量濃度對葡萄糖二酸產量的影響配制初始肌醇質量濃度分別為2、4、6、8、10 g/L和12 g/L的發酵培養基,初始葡萄糖質量濃度60 g/L,初始pH為5.5,將培養24 h的種子培養液以25%的接種體積分數轉接到發酵培養基中,培養60 h,見圖8。不同初始肌醇質量濃度對菌體量的影響不大。在肌醇質量濃度2~10 g/L范圍內,葡萄糖二酸產量隨初始肌醇質量濃度的增加而增加,當大于10 g/L時,葡萄糖二酸產量則趨于平穩。因此,初始肌醇質量濃度確定為10 g/L。
通過對碳源、初始pH、接種齡、接種體積分數及初始肌醇質量濃度搖瓶水平的優化,葡萄糖二酸的產量達到了(967.60±3.90)mg/L,相比優化前的產量(566.36±16.98) mg/L,提高了 70.85%。
2.4 發酵罐補料分批培養的研究
依據搖瓶優化的結果,將重組菌P.pastoris GS115-TUM在3 L發酵罐中進行放大實驗。以改良BSM為發酵培養基,初始葡萄糖質量濃度60 g/L,肌醇質量濃度13 g/L。發酵培養過程中,為保證碳源的充足,實現菌體高密度培養,當殘糖質量濃度低于5 g/L時,流加800 g/L葡萄糖母液 (含12 mL/L PTM1),維持殘糖質量濃度在0~5 g/L之間,結果見圖9。葡萄糖二酸的生成與菌體的生長呈正相關關系,因此可以判斷為生長偶聯型。在發酵60 h之前,肌醇的質量濃度下降明顯,此時葡萄糖二酸快速積累。發酵60 h之后,肌醇的質量濃度不再有明顯下降,而葡萄糖二酸的產量也趨于穩定。由此可以看出,培養基中添加肌醇有助于葡萄糖二酸的生成,且葡萄糖二酸的積累與肌醇的消耗成正相關關系。發酵培養96 h后,葡萄糖二酸的產量達到了(2.60±0.04) g/L。

圖8 初始肌醇質量濃度對菌體生長及產量的影響Fig.8 Effect of initial concentration of myo-inositol on recombinant strain and glucaric acid production
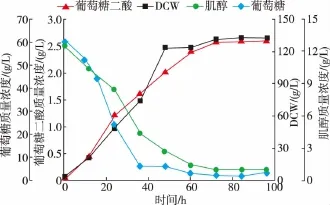
圖9 重組菌3 L發酵罐補料分批發酵過程曲線Fig.9 Fed-batch fermentation of recombinant strain in a3 L fermenter
3 結語
葡萄糖二酸作為一種自然界存在的高附加值有機酸,具有重要的生物功能。作者通過在畢赤酵母內異源表達小鼠來源的MIOX基因及Pseudomonas putida KT2440來源的udh基因,成功構建葡萄糖二酸合成途徑,在添加肌醇的搖瓶培養基中,葡萄糖二酸產量為(566.36±16.98) mg/L。通過對碳源、初始pH、接種齡及接種體積分數的優化,葡萄糖二酸產量達到(967.60±3.90)mg/L。按照搖瓶優化的條件,我們將重組菌在3 L發酵罐上進行補料分批培養,在補加葡萄糖及肌醇的情況下,葡萄糖二酸的產量達到(2.60±0.04) g/L。
本研究提出的重組畢赤酵母生產葡萄糖二酸的方法,具有廣闊的發展前景,為生物法合成葡萄糖二酸提供了新的思路。今后工作將通過對畢赤酵母內葡萄糖二酸代謝途徑進行分子層面的改造及優化,以期進一步提高葡萄糖二酸的產量。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
