柑橘采后病害生防木霉的篩選與鑒定
2018-11-05 01:55:54羅紅霞欒豐剛
生物災害科學 2018年3期
羅紅霞,欒豐剛,張 揚,李 冬*
?
柑橘采后病害生防木霉的篩選與鑒定
羅紅霞1,欒豐剛2,張 揚2,李 冬2*
(1. 江西省九江市林業有害生物防治檢疫局,江西 九江 332000;2. 江西農業大學 林學院,江西 南昌 330045)
從毛竹林土壤中分離得到9株木霉菌株,采用平板對峙法評估了9株木霉菌對4種柑橘產后病害病原菌的抑制效果,并利用ITS及2基因序列分析鑒定木霉菌株。結果發現:(1)9株木霉對柑橘青霉病菌具有較好的抑制效果,對峙培養7 d的抑制率均達到100%;9株木霉對其它3種柑橘產后病害的抑制率不同,其中菌株T2對柑橘黑腐病抑菌效果最好,抑制率達85.84%;菌株T3和T8對柑橘酸腐病菌抑制效果最好,抑制率達100%;菌株T3、T5和T6對柑橘綠霉病菌抑制效果最好,抑制率達80%。(2)共鑒定木霉菌3種,其中哈茨木霉()6株(T1、T3、T5、T7、T8和T9),為優勢種;綠木霉()2株(T2和T4);綠色木霉()1株(T6)。
生物防治;木霉菌;拮抗;柑橘采后病害
木霉屬是土壤微生物的重要類群,廣泛分布于世界各地。由于木霉可以產生纖維素酶及其它水解酶和多種次生代謝產物,具有重要的經濟價值。同時,木霉具有顯著的植物病原菌拮抗活性和生物防治潛力,國內外學者對其拮抗作用及機制作了較為系統深入的研究,并開發了多種商業化生防菌劑及生物菌肥用于農作物保護[1-4],該屬真菌已經成為研究和應用最多的植物病害生防菌。
自Persoon(1794)建立了木霉菌屬(Pers.)以來,關于該屬的分類經歷了較大的變化,如e的有性型為[5]、9個集合種(species aggregate)分類體系[6]、5個組(section)分類系統[7]以及將Saturnisporum組并入到Longibrachiatum組的4個組分類體系[8]。隨著分子系統學應用于木霉屬分類研究,為開展木霉屬的鑒定提供了有效的技術參考。如Druzhinina等[9]建立了木霉屬與肉座菌屬物種的DNA條形碼序列(ITS1和ITS2)鑒別系統OKey v. 1.0。Kopchinskiy等[10]建立了專門用于木霉屬與肉座菌屬物種序列檢索的BLAST數據庫。Zhu等[11]比較了基于、2和1基因的種內與種間序列差異及序列獲取難易程度,認為2最小種間差異大于最大種內差異,且PCR擴增與測序成功率較高(94.4%),可作為木霉屬的補充條形碼。
隨著木霉分子系統學研究加速,新的物種不斷被發現。截至2015年,木霉屬包括254個種及2個變種[11]。我國學者也在木霉屬資源與系統分類研究方面做了大量工作,迄今我國已報道的木霉屬真菌有118種[12-19],體現出了較高的物種多樣性。然而,我國目前對木霉菌屬的研究多集中在華東、西南和東北地區,而其他地區的研究相對較少。
柑橘是世界第一大果[20],也是我國南方栽培的主要水果之一,具有重要的經濟價值[21]。目前,我國柑橘仍以鮮果銷售為主,為滿足長期供應,柑橘需要在采收后經過較長時間的貯藏與運輸。由于柑橘果實含水率高、營養豐富,在采收、貯藏及運輸過程中極易受到各種病原真菌感染[22]。目前,常見且發生嚴重的柑橘產后病害主要有綠霉病()、青霉病()、酸腐病()[22]及黑腐病(),其中綠霉病與青霉病引起的損失占所有采后病害90%以上[23]。據統計,我國每年柑橘因采后病害而造成的損失一般為10%左右,個別年份高達30%以上[24]。因此,如何預防及降低柑橘采后病害的發生已成為國內外開發研究的熱點之一。
迄今為止,國內外控制柑橘采后病害主要措施仍是化學殺菌劑處理,常見藥劑有多菌靈、甲基托布津、苯萊特等。由于化學農藥的長期使用,除了農藥殘留給人們健康帶來隱患外,還導致病原菌對殺菌劑產生抗藥性,降低了化學殺菌劑的防治效果。因此,本研究擬重點解決以下問題:(1)分離篩選柑橘主要產后病害病原菌拮抗木霉;(2)明確拮抗木霉的分類地位。通過試驗研究,以期為木霉菌在柑橘采后病害生物防治中的應用提供菌種資源及參考依據。
1 材料與方法
1.1 供試菌株和材料
供試菌株:4種柑橘采后病害病原菌由江西農業大學農學院植物保護系向妙蓮老師惠贈;木霉菌(T1-T9)由江西農業大學園林與藝術學院森林保護實驗室分離和保存。
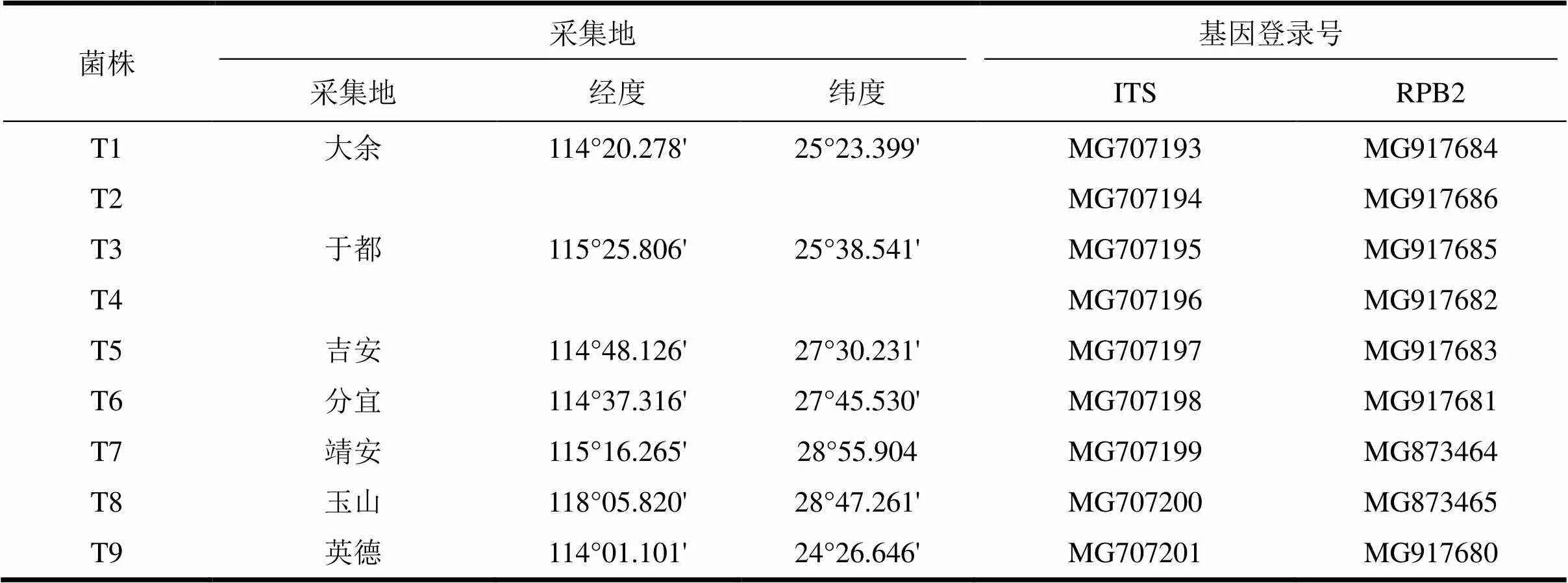
表1 供試木霉菌株信息
1.2 拮抗木霉菌的篩選
參考Dennis & Webster(1971)[25]的平板對峙法并稍作修改。將已初篩具有拮抗活性的木霉菌株與病原菌平板培養后,使用打孔器將病原菌和木霉切成直徑6 mm的菌絲塊,木霉接種至PDA平板中央,病原菌接種至距離平板邊緣約2 cm處,每皿放4塊病原菌,置放入25 ℃恒溫培養,每個菌株設置3個重復,以僅接種病原菌純培養為對照。168 h后觀察并測量病原菌的菌落半徑,按以下公式計算木霉菌的抑制率:
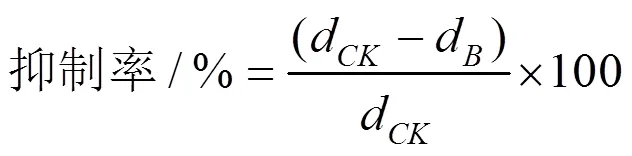
其中:d為對照組接種植物病原菌后168 h的直線生長距離;d為對峙培養組接種植物病原菌后168 h的直線生長距離。
1.3 拮抗木霉菌的鑒定
采用CTAB法提取木霉菌株總DNA,采用引物ITS4和ITS5對ITS序列進行擴增[26],用引物983F和2218R對RPB2序列進行擴增[27]。雙向測序后采用DNAstar中的Seqman 7.1.0軟件包[28]進行序列拼接,并采用人工校正個別位點的錯誤信息。將測得ITS及RPB2序列提交至GenBank獲得登錄號,并分別在ISTH(International Subcommission on Trichoderma and Hypocrea Taxonomy)的OKEY v. 2.0[8]和BLAST v. 1.0[9]進行Blast比對鑒定其分類地位。
2 結果與分析
2.1 拮抗菌對幾種植物病原菌的拮抗效果
由表2可看出:9株木霉菌對柑橘青霉病菌抑制效果較好,抑制率均為100%;對柑橘黑腐病拮抗效果最好的是菌株T2,菌株T6次之;對柑橘酸腐病菌拮抗效果最好的是菌株T3和T8,抑制率為100%;對柑橘綠霉病菌抑制效果較好的木霉菌株是T3、T5和T6,抑制率為80%。

圖1 不同木霉菌株對柑桔酸腐病菌的拮抗作用
從木霉菌株來看,除菌株T9對柑橘黑腐病抑制率(35.39%)較低外,其余8株木霉對4種柑橘產后病害病原菌的抑制率均超過了60%,即8株木霉對4種柑橘產后病害病原菌有一定的抑菌活性。
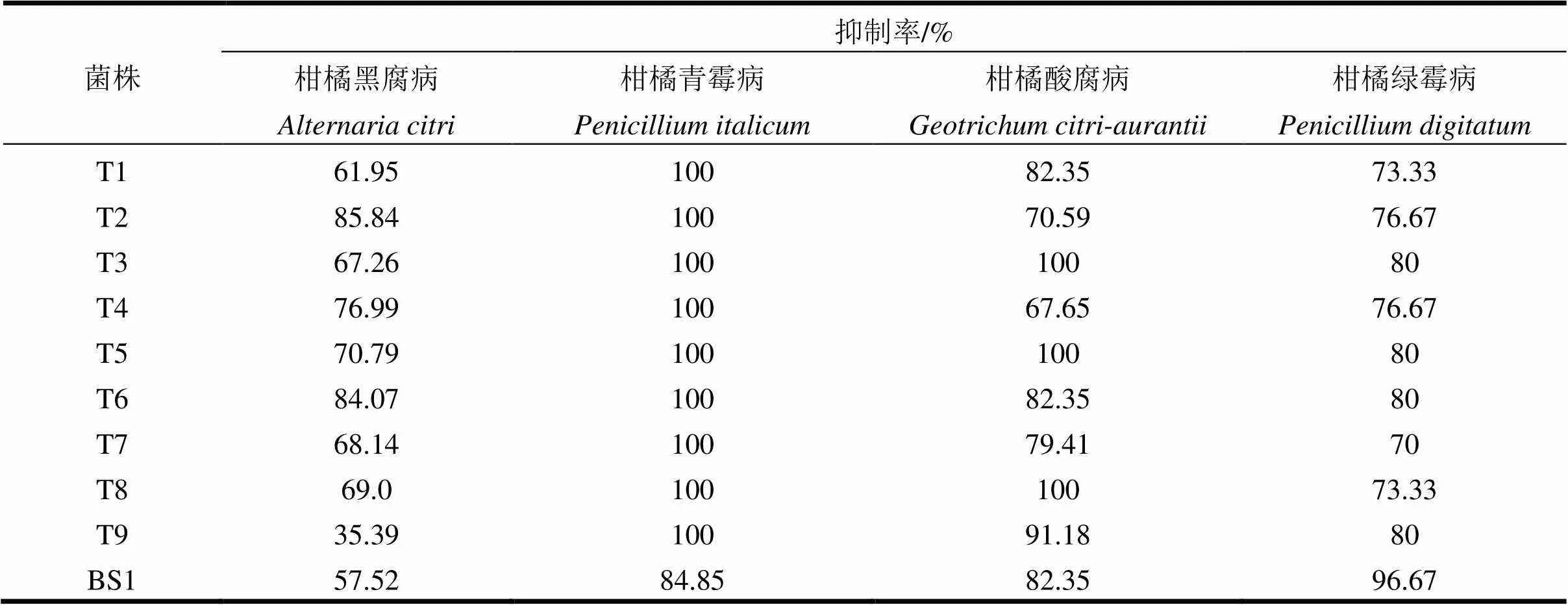
表2 木霉菌株對不同植物病原菌的抑制率
2.2 拮抗木霉的分子鑒定
基于ITS基因序列在OKEY v. 2.0比對結果可知:本研究的9株木霉菌以哈茨木霉()為主,其中6株木霉(T1、T3、T5、T7、T8和T9)在系統發育上與哈慈木霉親緣關系最近;菌株T6與綠木霉()親緣關系最近,而菌株T2和T4與擬康寧木霉()親緣關系最近。

表3 基于ITS序列的TrichOKEY v2.0比對結果
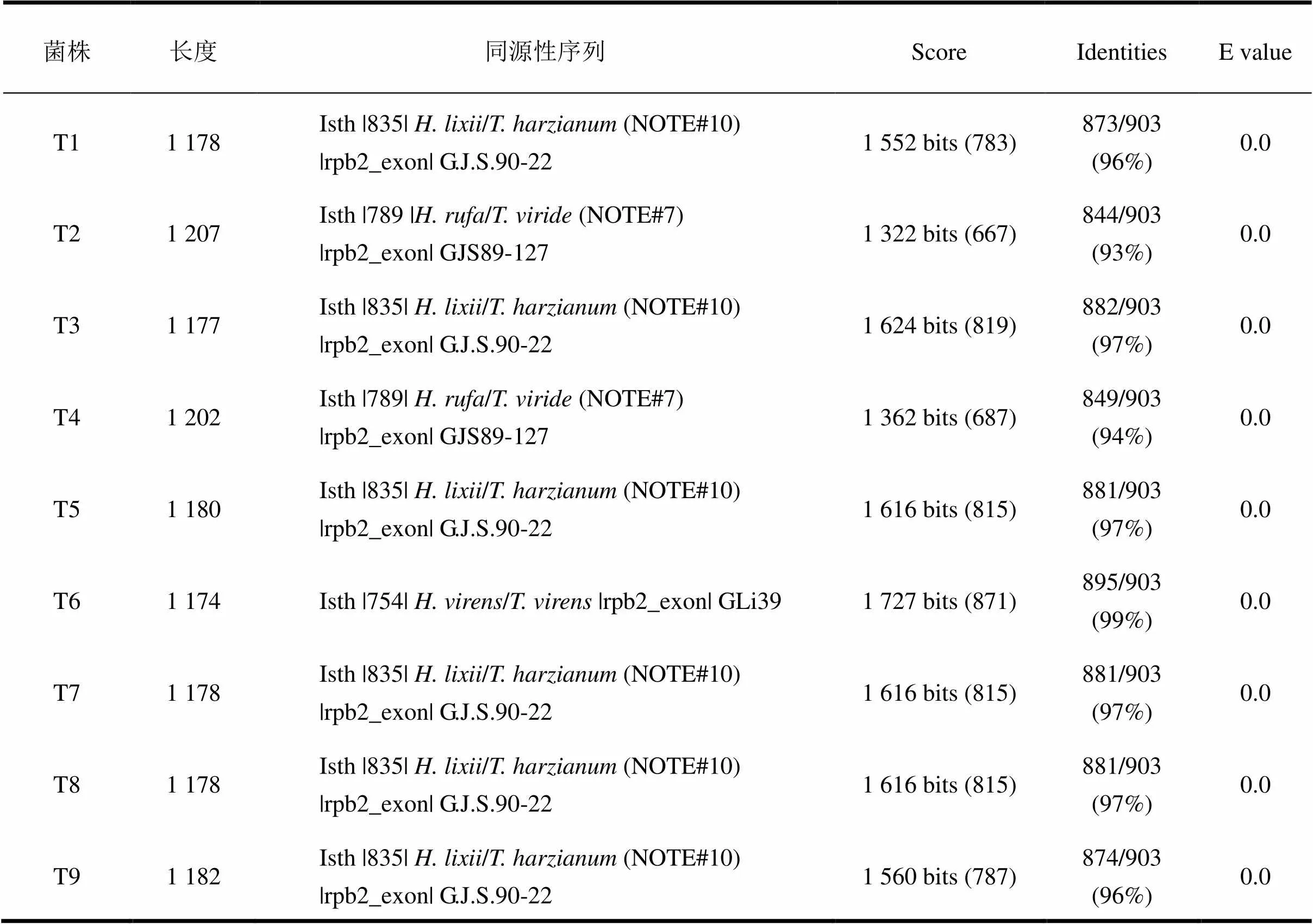
表4 基于rpb2基因的TrichBLAST v.1.0比對結果
基于2基因序列輸入BLAST v. 1.0比對結果可知:7株木霉(T1、T3、T5、T6、T7、T8和T9)與OKEY v. 2.0比對結果基本一致,而2株木霉(T2和T4)基于2基因序列blast結果為綠色木霉()。依據Zhu等[11]對于木霉菌ITS、2和1基因鑒定準確性的評價,2基因比ITS基因更適合作為木霉屬的補充條形碼,本研究據此認為菌株T2和T4鑒定為綠色木霉()更為合適。
3 討 論
隨著農業生產的發展以及人們對果蔬產品需求量的增加,柑橘作為世界主要水果之一,其種植面積及產量也不斷擴大,而其產后病害也日益突出,成為影響柑橘產業發展的制約因子之一。目前,化學防治仍然是柑橘產后病害的主要措施之一。然而,殺菌劑的廣泛使用帶了諸多問題,如農藥殘留、病原菌抗性增加、生態平衡破壞等,進而造成殺菌劑效果下降、用量增加,甚至防治無效等后果[29-30]。如在柑橘上長期適應苯并咪唑類殺菌劑,使得原來對其不敏感的柑橘黑腐病與柑橘酸腐病菌種上升成為主要病害病原菌[31]。因此,尋找安全、經濟、高效的生物防治成為研究的熱點。
迄今為止,利用拮抗真菌對植物病害進行生物防治的研究相當多,但多數研究多限于實驗室條件下,生產應用較為成功僅為少數,如木霉()、盾殼霉()、毛殼霉()等,其中以木霉作為生物載體的生防菌劑或菌肥開發應用最多,全球已注冊的產品超過50種[32],如以色列MakhteshimAgan公司以哈茨木霉為生物載體開發的產品Trichodex用于防治由灰葡萄孢引起的蔬菜和葡萄病害;新西蘭Agrimm Bilogicals公司以木霉(包括哈茨木霉和綠色木霉)為生物載體開發的產品Trichodwels用于防治各種土傳病害及由紫韌革菌引起的銀葉病;美國生物農藥公司BioWorks以木霉(包括哈茨木霉和綠色木霉)為生物載體開發的RootShield用于防治輸出及其它作物猝倒病。由此可見,木霉在植物病害生物防治方面展現出了巨大的應用潛力。
本研究的9株木霉對4種主要的柑橘產后病害均有一定的抑菌活性,除菌株T9對柑橘黑腐病菌抑菌效果較差外,其余8株木霉對4種柑橘產后病害均表現出一定的抑菌活性。9株木霉除了對柑橘青霉病菌抑制效果一致較好以外,而對其它3種柑橘產后病害表現出不同的抑制效果。從生長速度上看,木霉菌具有高于柑橘產后病害病原菌的生長速度,因此,木霉菌可覆蓋病原菌并通過競爭作用達到抑菌目的。由于柑橘產后病害防治對象為柑橘果實,直接以拮抗木霉開發菌劑(如粉劑、水劑等)進行生物防治可能存在諸多問題。課題組進行了木霉發酵液粗提物平板抑菌(圖2)初步試驗,結果發現木霉粗提物中含有抑菌物質,今后將重點獲得純化的抑菌活性成分,將對開發利用木霉防治柑橘產后病害更為有利。
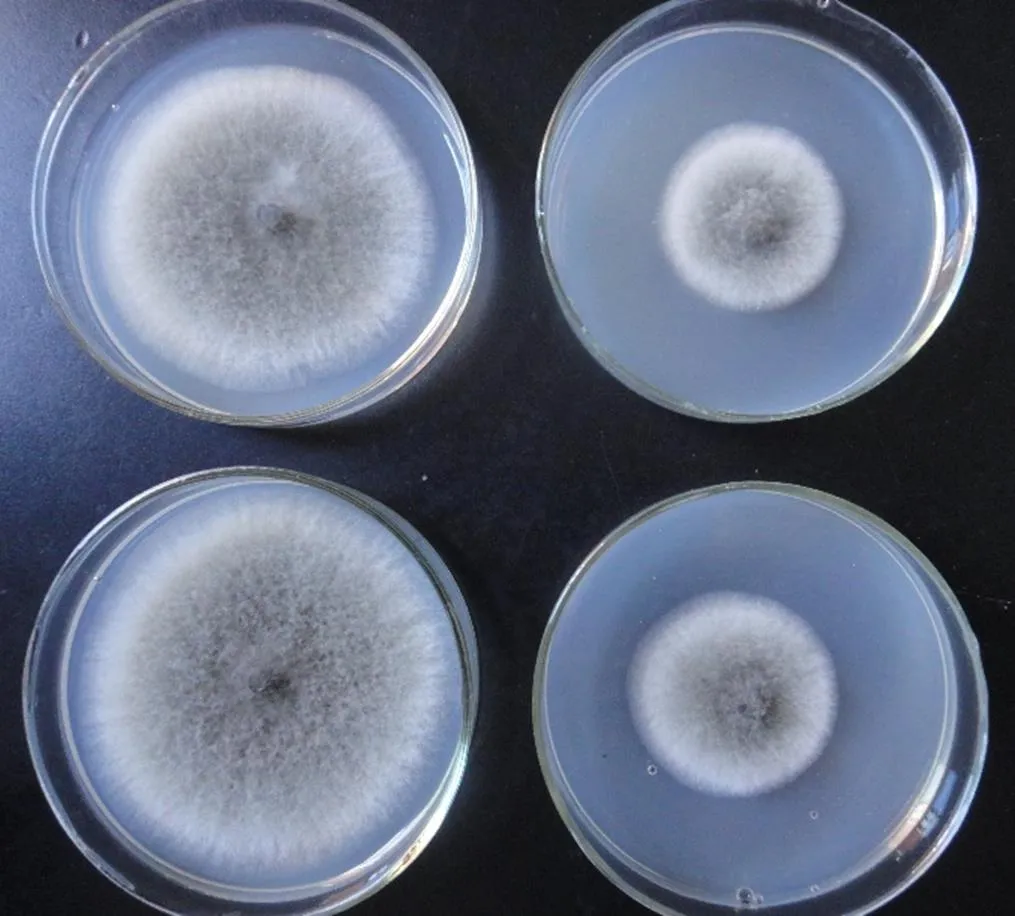
圖2 木霉菌株T9發酵液對柑桔酸腐病菌的抑制作用
本研究通過系統發育分析將供試9株木霉鑒定為3種:其中包括6株哈茨木霉(),2株為綠色木霉()及1株綠木霉(),表現出較高的物種多樣性。結合9株木霉對3種柑橘產后病害(柑橘青霉病除外)病原菌的抑菌效果來看:綠色木霉菌株T2對柑橘黑腐病抑制效果最好;哈茨木霉菌株T3和T8對柑橘酸腐病抑菌率最高;而哈茨木霉菌株T3與T5及綠木霉菌株T6則對柑橘綠霉病表現出較好的抑菌活性。然而,本試驗尚未就供試菌株的抑菌機理及實際防病效果進行研究,后續試驗將對上述問題做進一步研究。
[1] Tripathi P, Dubey N K. Exploitation of natural products as an alternative strategy to control postharvest fungal rotting of fruit and vegetables[J]. Postharvest Biology & Technology, 2004, 32(3): 235-245.
[2] Asad S A, Ali N, Hameed A, et al. Biocontrol efficacy of different isolates ofagainst soil borne pathogen[J]. Pol J Microbiol, 2014, 63(1): 95-103.
[3] Zafari D, Koushki M M, Bazgir E. Biocontrol evaluation of wheat take-all disease byscreened isolates[J]. African Journal of Biotechnology, 2008, 7(20): 3650-3656.
[4] Benítez T, Rincón A M, Limón M C, et al. Biocontrol mechanisms ofstrains[J]. International Microbiology the Official Journal of the Spanish Society for Microbiology, 2004, 7(4): 249.
[5] Lorito M, Woo S L, Harman G E, et al. Translational research on: from 'Omics to the field[J]. Annual Review of Phytopathology, 2010, 48(1): 395.
[6] Rifai M A. A revision of the genus[J]. Mycol Pap, 1969, 116: 1-54.
[7] Bissett J. A revision of the genusI. Section Longibrachiatum sect[J]. Canadian Journal of Botany, 1984, 62(5): 924-931.
[8] Gams W, Bissett J. Morphology and identification of[M]//Kubicek C P, Harman G E.and: Vol 1: Basic biology, taxonomy and genetics. London: Taylor and Francis Ltd. 1998: 3-34.
[9] Druzhinina I S, Kopchinskiy A G, Komoń M, et al. An oligonucleotide barcode for species identification in Trichoderma and Hypocrea[J]. Fungal Genetics & Biology, 2005, 42(10): 813-828.
[10] Kopchinskiy A, Komon M, Kubicek Cp, et al. TrichoBLAST: a multilocus database forandidentifications[J]. Mycological Research, 2005, 109(6): 658-660.
[11] Zhu Z X, Zeng Z Q, Zhuang W Y. Selection of a supplementary DNA barcode for the genus(Hypocreales, Ascomycota)[J]. Mycosystema, 2014, 33(6): 1253-1262.
[12] John B, Walter G, Walter J, et al. Acceptednames in the year 2015[J]. Ima Fungus, 2015, 6(2): 263-295.
[13] 張廣志, 楊合同, 張新建, 等. 木霉現有種類名錄[J]. 菌物學報, 2014(6): 1210-1230.
[14] 朱兆香, 莊文穎. 木霉屬研究概況[J]. 菌物學報, 2014(6): 1136-1153.
[15] 秦文韜, 陳凱, 莊文穎. 木霉屬 5 個中國新記錄種及 3 種木霉在中國的新分布[J]. 菌物學報, 2016, 35(10): 1-15.
[16] 朱兆香, 李玉. 木霉屬3個中國新記錄種[J]. 菌物學報, 2016, 35(11): 1008-1017.
[17] 陳凱, 莊文穎. 來自土壤的木霉屬4個中國新記錄種[J]. 菌物學報, 2017, 36(12): 1-10.
[18] 陳凱, 李紀順, 張廣志, 等. 中國新記錄種毛細木霉及功能評價[J]. 山東科學, 2016, 29(1): 93-97.
[19] 陳凱, 李紀順, 王貽蓮, 等. 洞庭湖濕地木霉多樣性及生防活性[J]. 微生物學通報, 2017, 44(10): 2307-2320.
[20] 張廣志, 張新建, 李成云, 等. 木霉屬7個中國新記錄種(英文)[J]. 菌物學報, 2016, 35(8): 984-993.
[21] 鄧秀新郭文武. 國際柑橘學會第九屆學術大會介紹[J]. 果樹學報, 2001(2): 124.
[22] 陳仕俏, 趙文紅, 白衛東. 我國柑橘的發展現狀與展望[J]. 農產品加工(學刊), 2008(3): 21-24.
[23] Zhang Z F, Zhu Z R, Ma Z H, et al. A molecular mechanism of azoxystrobin resistance inUV mutants and a PCR-based assay for detection of azoxystrobin-resistant strains in packing- or store-house isolates[J]. International Journal of Food Microbiology, 2009, 131(2/3): 157-161.
[24] 陸寧海, 徐瑞富, 吳利民, 等. 柑桔青霉菌侵染條件及致病性研究[J]. 東南園藝, 2005(1): 6-8.
[25] Dennis C, Webster J. Antagonistic properties of species-groups of: III. Hyphal interaction[J]. Transactions of the British Mycological Society, 1971, 57(57): 363-369.
[26] White T J, Bruns T, Lee S, et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[J]. PCR Protocols: a guide to methods and applications, 1990, 18: 315-322.
[27] Liu Y J, Whelen S, Hall B D. Phylogenetic relationships among ascomycetes: evidence from an RNA polymerse II subunit[J]. Molecular Biology & Evolution, 1999, 16(12): 1799-1808.
[28] Burland T G. DNASTAR’s Lasergene sequence analysis software[J]. Methods in Molecular Biology, 2000, 132: 71-91.
[29] Eckert J W, Ogawa J M. The chemical control of postharvest diseases: deciduous fruits, berries, vegetables and root/tuber crops[J]. Annual Review of Phytopathology, 1988, 26(1): 433-469.
[30] Zhu J W, Xie Q Y, Li H Y. Occurrence of imazalil-resistant biotype ofin China and the resistant molecular mechanism[J]. Journal of Zhejiang Universityence A, 2006, 7(2): 362-365.
[31] 李紅葉曹若彬. 果蔬產后病害生物防治研究進展[J]. 生物防治通報, 1993(4): 176-180.
[32] Woo S L, Scala F, Ruocco M, et al. The molecular biology of the interactions betweenspp., Phytopathogenic fungi, and plants[J]. Phytopathology, 2006, 96(2): 181.
Screening and Identification ofStrains in Biological Control of Post-harvest Diseases of Citrus
LUO Hong-xia1, LUAN Feng-gang2, ZHANG Yang2, LI Dong2*
(1. Forest Pest Control and Quarantine Bureau of Jiujiang, Jiujiang, Jiangxi 332000, China; 2. College of Forestry, Jiangxi Agricultural University, Nanchang 330045, China)
Nine strains ofspp. from soils in different bamboo forests in Jiangxi Province were evaluated, and their antagonism against,,, andwere got by flat dilution. And then strains ofspp. were identified with the sequences of ITS and2. The results showed that: (1) nine strains ofhad antagonism against, and the inhibition effects (IE) reached 100% in 7 days. There were different inhibition effect on other three pathogens of,, and. Thestrain T2 had the significant antifungal effects on, with the IE up to 85.84%. Thestrains T3 and T8 had powerful antagonism against, with the IE up to 100%. And the inhibitory effects ofstrains T3, T5 and T6 were the best, with the IE up to 80%. (2) Three species among the nine strains ofspp. were identified, of whichwas a dominant species, which included six strains (T1、T3、T5、T7、T8 and T9). Two strains ofspp. (T2 and T4) belonged to, while only onestrain (T6) belonged to.
biological control;; antagonist; post-harvest diseases of citrus
S436.66.1+9
A
2095-3704(2018)03-0192-06
2018-07-18
國家自然科學基金(31560203)、江西省自然科學基金項目(20171BAB204026)和江西省教育廳科技計劃項目(GJJ160363)
羅紅霞(1971—),女,工程師,主要從事林業微生物研究,LFG7869@126.com;
李冬,副教授,jxld2008@163.com。
羅紅霞, 欒豐剛, 張揚, 等. 柑橘采后病害生防木霉的篩選與鑒定[J]. 生物災害科學, 2018, 41(3): 192-197.
10.3969/j.issn.2095-3704.2018.03.42