葉面噴施鉀鎂鈣肥對桃果實著色、品質及著色相關基因表達的影響
2018-11-05 04:02:16何平李林光王海波常源升
山東農業科學 2018年10期
關鍵詞:差異
何平, 李林光, 王海波, 常源升
(山東省果樹研究所, 山東 泰安 271000)
桃(Prunuspersica) 屬于薔薇科(Rosaceae)李屬(Prunus)。中國是世界桃的第一生產大國,面積和產量均居世界首位。 桃是我國為數不多的具有明顯國際競爭力的果品之一。桃生產中, 存在病蟲害嚴重、著色不良、外觀品質差等問題, 降低了果實的商品性。為了促進桃果實著色, 改善果實品質, 生產上常通過加強肥水管理、減少負載量等手段以促進果實著色。前人已在多種園藝作物上開展了葉面噴施肥料促進果實著色的研究。關軍鋒等[1]研究表明, 在甜櫻桃果實成熟前施用Ca肥可顯著提高果實中花色苷含量。周開兵[2]、高丹[3]等一致認為, 在荔枝果實膨大期噴施P、K和Ca肥可促進果皮著色。于忠范等[4]在蘋果上噴施Ca肥, 明顯改善果實色澤;而金會翠[5]、姜學玲[6]等在蘋果上噴施K肥,增加了果皮花色苷含量, 促進果實著色。另有研究發現, 在葡萄[7,8]和溫州蜜柑[9]上葉面施用Mg肥, 可提高果實花色素含量。由些可見, 施用K、Mg、Ca肥可促進果實著色, 改善果實品質, 但目前有關K、Mg和Ca元素營養對桃果皮著色的機理研究較少。本試驗以生產上廣泛栽培的‘春雪’桃為材料, 在果實轉色期進行葉面噴施K、Mg、Ca肥, 研究其對果實色澤、品質及著色相關基因表達的影響, 以期為科學、合理使用葉面肥料改善桃果實品質提供理論依據。
1 材料與方法
1.1 試驗材料
試驗于2017年在山東省果樹研究所天平湖實驗基地進行。果園土質為砂壤土, 管理水平較好, 株行距為2 m×4 m, 主干型整枝。以6年生‘春雪’桃為材料, 選擇生長、結果、干周、冠徑及樹勢等基本一致健壯樹進行。
1.2 試驗設計及方法
選擇長勢基本一致的樹體作為試材, 于5月15日(轉色期)、5月20日分別用K、Mg、Ca肥整株噴施。參照高丹等[3]的方案, 設置7個肥料處理: Mg(1.5% MgCl2溶液)、K(0.5% KCl溶液)、Ca(0.3% CaCl2溶液)、K+Mg(0.5% KCl溶液與1.5%MgCl2溶液按7∶4混合)、K+Ca(0.5% KCl溶液與0.3%CaCl2溶液按7∶1混合)、Mg+Ca(1.5% MgCl2溶液與0.3% CaCl2溶液按4∶1混合)和K+Mg+Ca(0.5% KCl溶液、1.5% MgCl2溶液與0.3% CaCl2溶液按7∶1∶ 4混合), 以葉面噴清水為對照(CK)。單株小區,重復3次。第二次處理后, 每隔5 d取一次樣, 直至果實成熟。分別于5月25日(S1)、5月30日(S2)、6月5日(S3)、6月10日(S4)和6月15日(S5)沿樹冠外圍距地面1.5~2.0 m處隨機采收10個果實,重復3次。采后帶回實驗室, 檢測各項果皮色澤、果實品質指標。果皮用液氮迅速冷凍, -80℃保存備用。
1.3 測定指標及方法
果皮色澤參照高丹等[3]的方法采用全自動色差計測定,根據分布均勻的測定點果皮a、b值換算出色度角[h=tan-1(b/a)],h值越小果皮綜合色澤越紅。花色苷含量參照李合生[10]的比色法測定。葉綠素含量參照Arnon等[11]的方法測定。類黃酮含量參考 Meyers等[12]的提取方法, 采用 NaNO2-Al(NO3)3比色法測定。
果肉可溶性固形物含量用蒽酮比色法于620 nm波長下測定, 根據蔗糖的標準曲線計算可溶性固形物含量。
單果質量檢測: 每次采果后隨機選取30個果, 用游標卡尺(0.02 mm)測量果實的縱、橫、側徑, 用天平(0.01 g)稱重, 計算平均單果重。
1.4 樣品RNA提取、cDNA合成及熒光定量PCR分析
采用天根Trizol Universal總RNA提取試劑(DP424)提取桃果實表皮的RNA, 步驟參考其說明書。采用TransScript Ⅱ All-in-One First-Strand cDNA Synthesis SuperMix for qPCR(北京全式金)試劑盒對提取的RNA進行反轉錄; 用SYBRGreen染料進行熒光定量PCR(qRT-PCR)分析, 以2-ΔΔCt法計算基因的差異倍數。每個樣品重復3次。選用PpN1(ppa009483m)為內參基因, 引物序列如表1 所示。

表1 用于qRT-PCR分析的基因及其引物
1.5 數據分析
試驗數據用SPSS軟件統計分析, 每個數據分析重復5次, 用Microsoft Excel軟件制作圖表。
2 結果與分析
2.1 葉面噴肥處理對果皮著色的影響
由表2看出, 隨著桃果實成熟,著色增加,果皮色度角h 值呈下降趨勢。不同噴肥處理在果實轉色期對色度角h 值產生明顯的影響。在S1期, 不同噴肥處理的h值與CK無顯著差異; 在S2期, K、K+Mg和K+Ca處理的h值與CK差異不顯著, 其余噴肥處理的h值均顯著低于CK; 在S3期, 各噴肥處理的h值均顯著低于CK, 其中Mg、Ca和Mg+Ca處理的h值降低幅度較大; 在S4期, Mg、Ca、Mg+Ca和K+Mg+Ca處理的h值顯著低于CK, 其余處理的h值與CK無顯著差異; 在S5期, Ca處理的h值顯著低于CK, 其余處理的h值與CK差異不顯著。可見, 各處理在果實轉色期對果皮著色均有一定影響, 但Mg+Ca處理可以顯著地促進果皮提前著色。
2.2 葉面噴肥處理對果皮花色苷、葉綠素含量的影響
由表3看出, 隨著果實成熟, 果皮中花色苷含量均逐漸升高, 不同噴肥處理對春雪桃果皮花色苷含量有明顯影響。在S1、S2期, 各噴肥處理的果皮花色苷含量與同期CK差異不顯著; 在S3期, Ca、Mg+Ca、K+Mg+Ca處理的果皮花色苷含量顯著高于CK, 其余各噴肥處理的果皮花色苷含量與CK差異不顯著; 在S4期,除K、Mg處理外,其它處理均顯著高于對照;S5期, 各噴肥處理的果皮花色苷含量均顯著高于CK。可見, 不同噴肥處理可促進果皮花色苷積累。
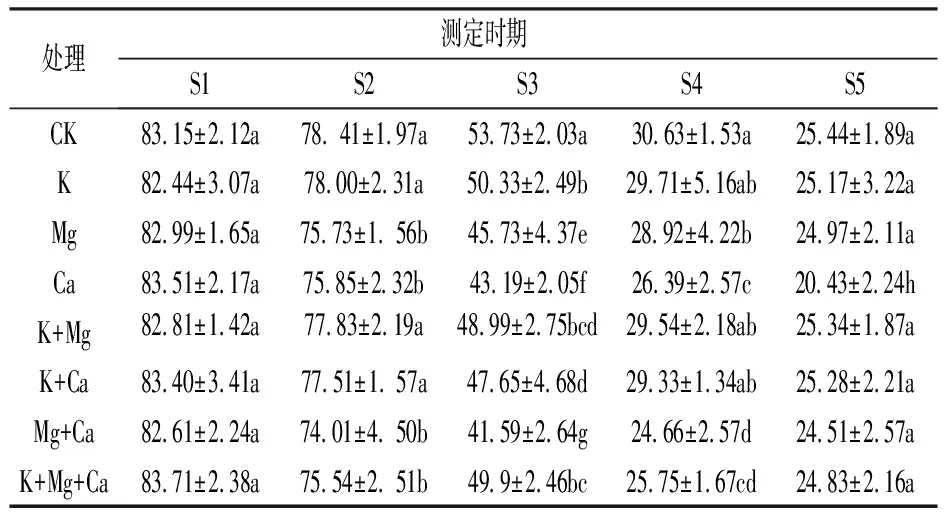
表2 不同噴肥處理對桃果皮色度角h值的影響
注: 同列數據后不同小寫字母表示差異達5%顯著水平。下同。
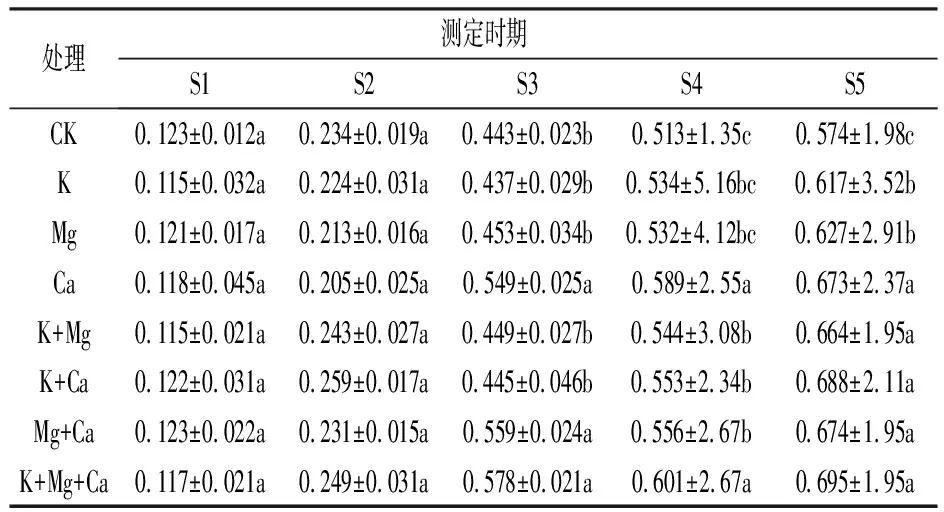
表3 不同噴肥處理對桃果皮花色苷含量的影響
如圖1所示, 在果實發育過程中, 果皮中葉綠素含量總體呈現下降趨勢。在S1和S2期, 各噴肥處理與同期對照差異不顯著; S3期Mg處理和S4期Mg+Ca處理的果皮中葉綠素含量顯著低于CK, 其余各處理差異不顯著; 在S5期, Ca、Mg+Ca處理的果皮葉綠素含量顯著低于CK, 其余各處理與CK差異不顯著。Ca與Mg+Ca處理在果實采收期(S5期),果皮中葉綠素含量分別為對照果實的43.3%與48.1%。
2.3 葉面噴肥處理對果實可溶性固形物、類黃酮含量的影響
如圖2所示, 隨著春雪桃果實成熟, 可溶性固形物含量逐漸升高, 不同處理對果肉可溶性固形物含量產生一定影響。在S1期, 不同噴肥處理與CK差異不顯著; 在S2期, K處理優于Ca處理, 且兩個處理均顯著高于CK; 在S3期, Ca處理效果最好, K處理次之, K+Mg+Ca處理略好于CK, 其余各處理低于CK; 在S4期, Ca處理效果最佳, Mg+Ca處理次之, 隨后為Mg處理和K處理, 且均顯著高于CK; 在S5期, Ca處理效果最佳, K處理次之, 隨后為Mg+Ca、K+Mg+Ca處理, 且均顯著高于CK, 其余處理與CK差異不顯著。在果實采收期, Ca處理果實中可溶性固形物含量比對照高29.3%。
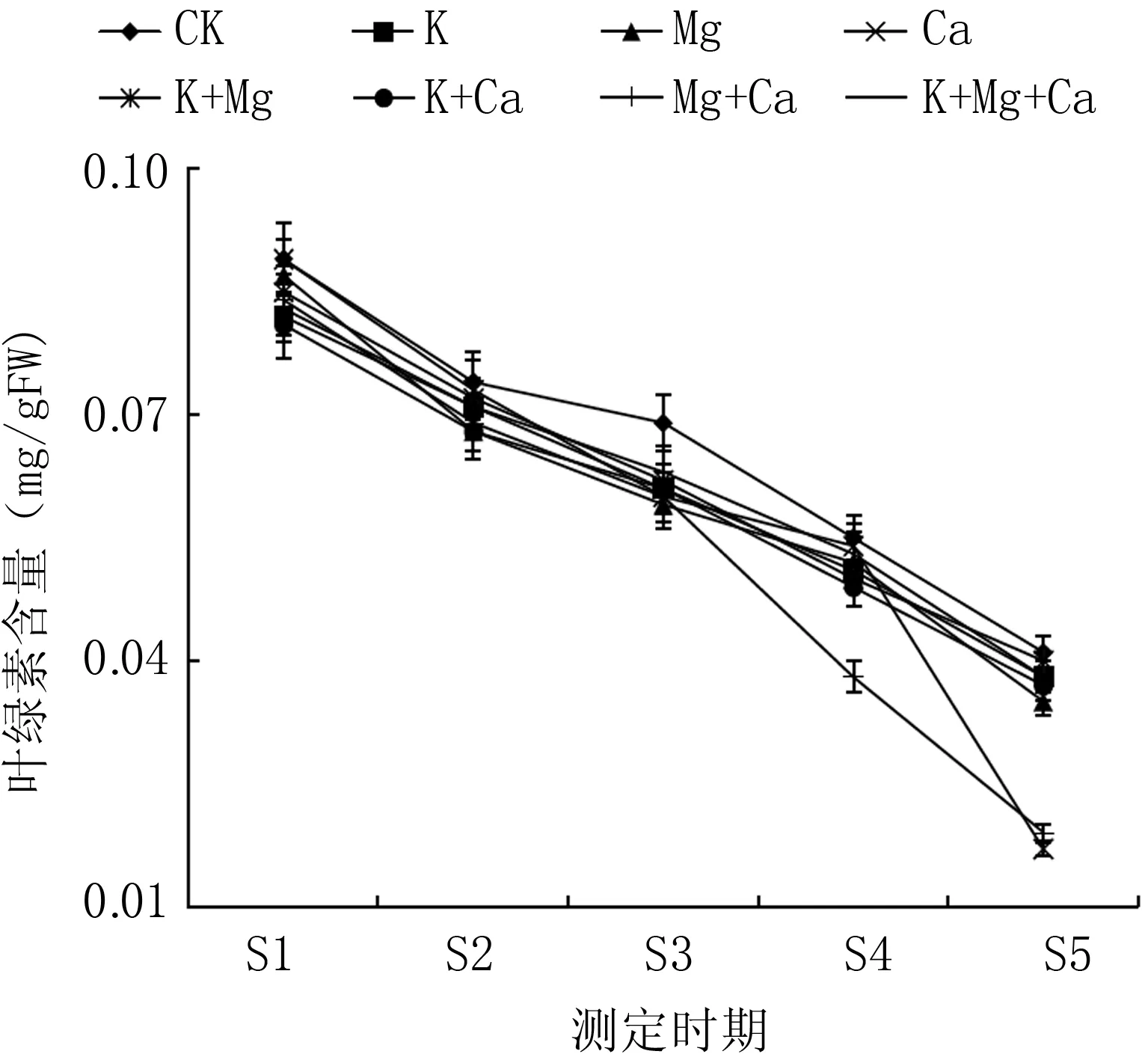
圖1 不同噴肥處理對桃果皮葉綠素含量的影響
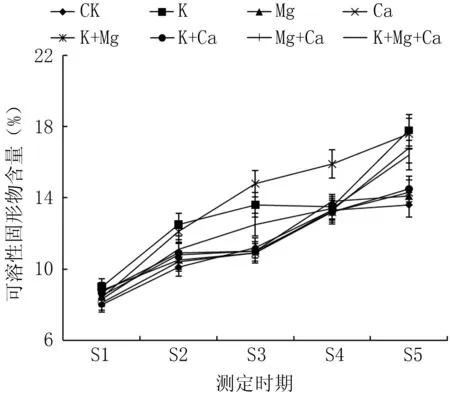
圖2 不同噴肥處理對春雪桃果實可溶性固形物含量的影響
如圖3所示, 春雪桃果實成熟過程中, 果皮中類黃酮含量整體呈現下降趨勢; 不同噴肥處理不同程度地降低春雪桃果皮中類黃酮含量。在S1、S2期, 各噴肥處理的果皮類黃酮含量與同期CK差異不顯著; 在S3期,各噴肥處理的類黃酮含量均低于對照,除Mg、K+Mg外均與CK差異顯著; 在S4、S5期, 各噴肥處理的果皮類黃酮含量顯著低于CK。在果實采收期(S5期), Mg+Ca和K+Mg+Ca處理春雪桃果皮中類黃酮含量分別比對照低30.8%和23.2%。
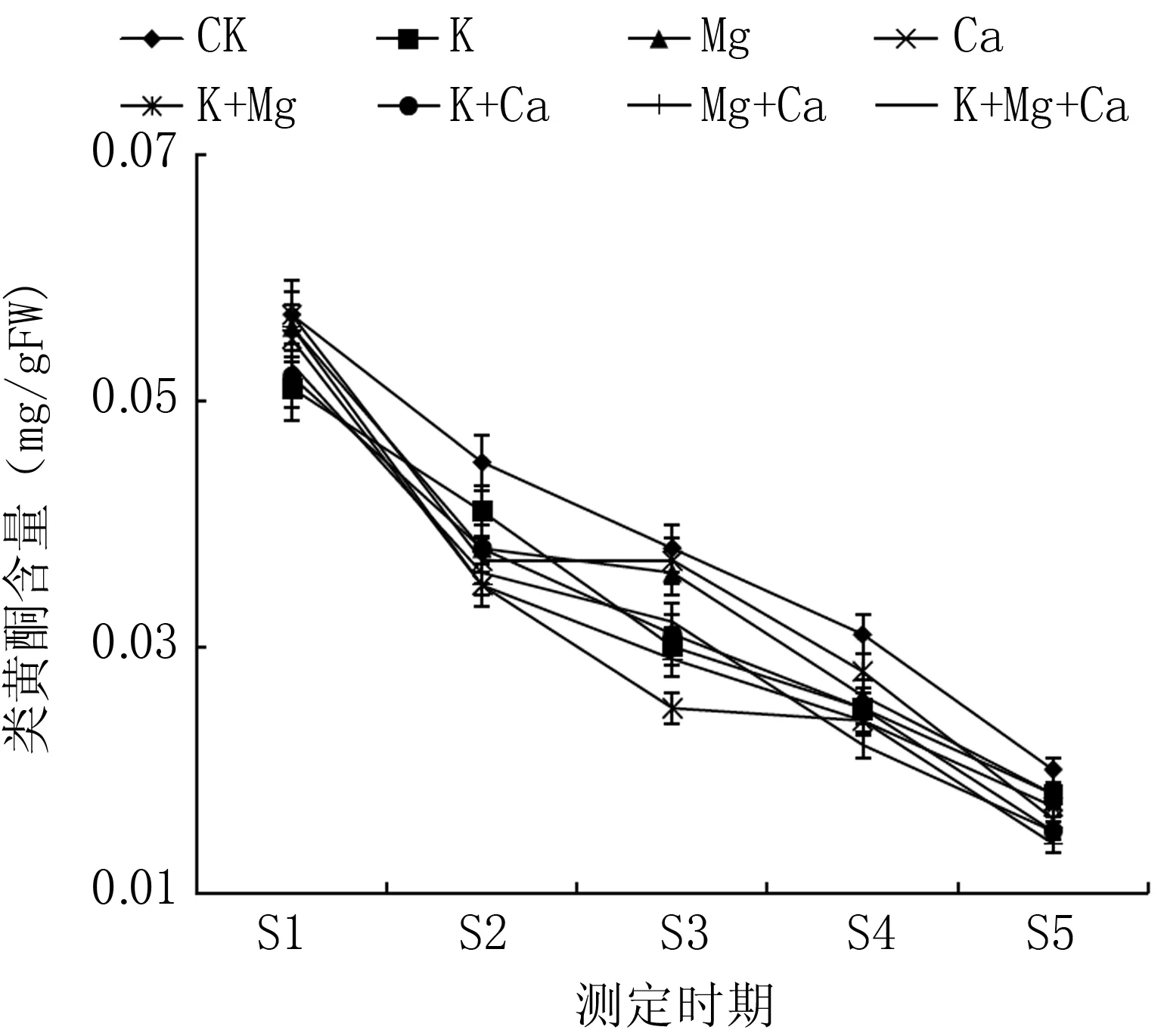
圖3 不同噴肥處理對桃果皮類黃酮含量的影響
2.4 葉面噴肥處理對果實大小及形狀的影響
如表4所示, 在果實采收期(S5期)不同噴肥處理對春雪桃果實單果重沒有顯著影響, 各處理與對照在果實縱、橫、側徑及果形指數上也不存在顯著差異。由此可知, 葉面噴肥對春雪桃產量影響不大。
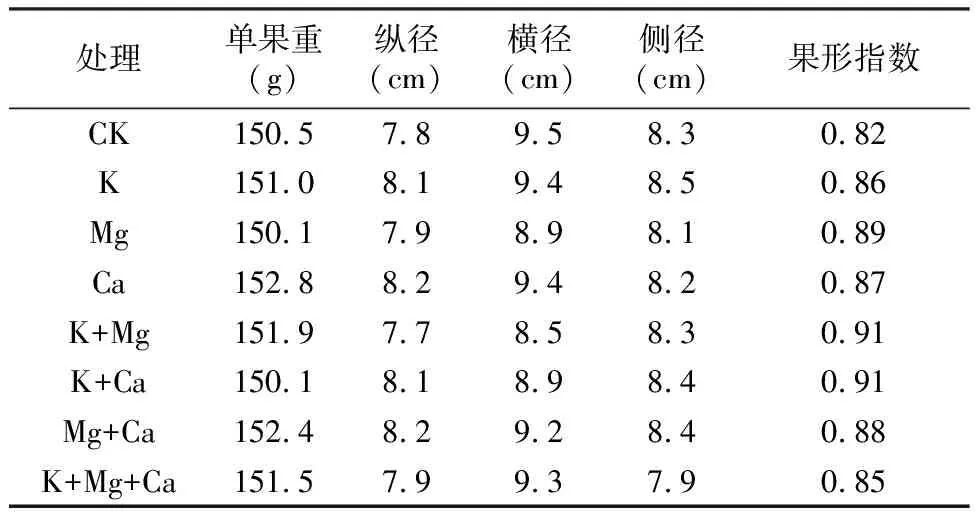
表4 不同噴肥處理對桃單果重、果實縱橫側徑的影響
2.5 葉面噴肥處理對花色苷代謝相關基因表達的影響
二氫黃酮醇還原酶(DFR)、無色花色素雙加氧酶(LDOX)、類黃酮糖基轉移酶(UFGT)是花色苷生物合成的關鍵酶。如圖4所示, 轉色初期, 基因DFR、LDOX、UFGT表達量較低, 隨著春雪桃果實成熟, 基因表達量迅速升高, 均在果實采收前期(S4期)達到峰值, 其后表達量降低。在果實采收期(S5期), Ca與Mg+Ca處理果實著色相關基因DFR、LDOX、UFGT的表達量均高于對照,兩處理UFGT基因的表達量分別為對照的2.54倍與2.18倍。
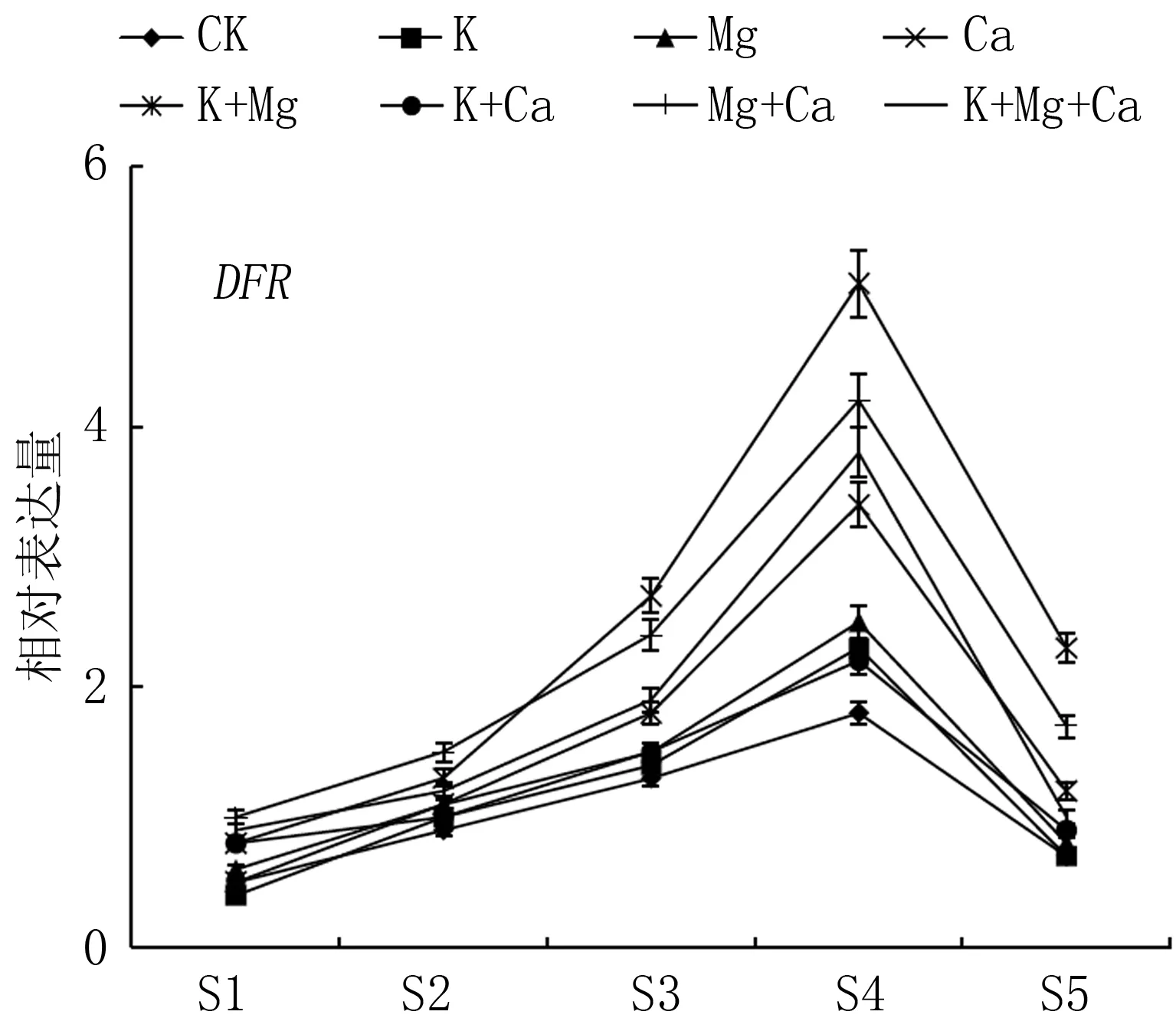
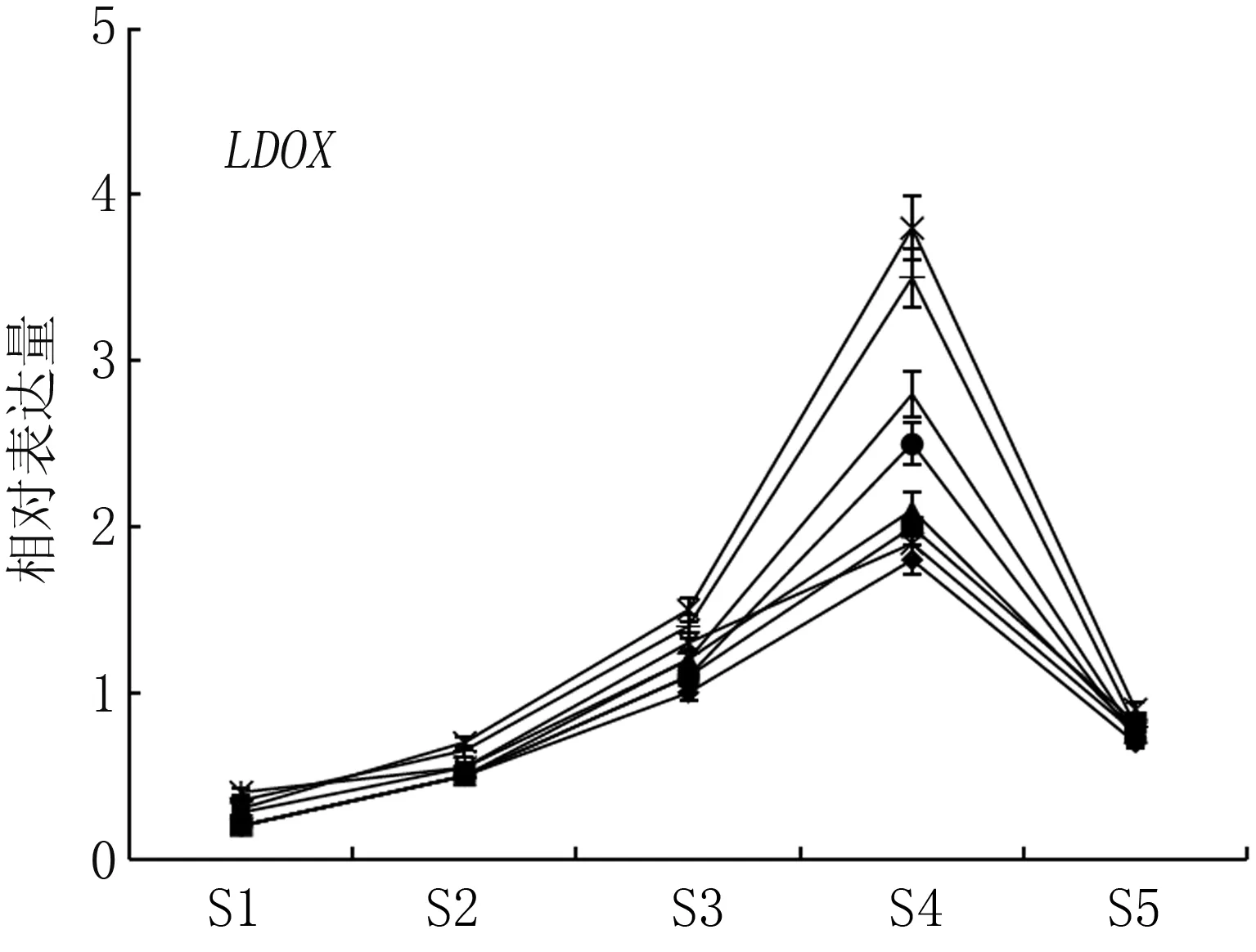
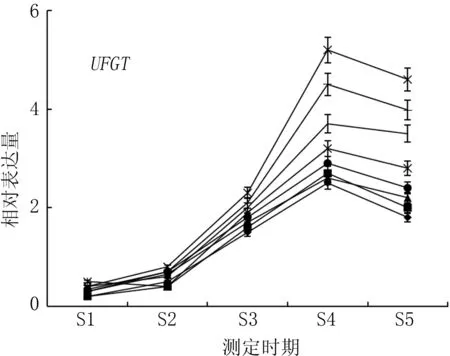
圖4 不同噴肥處理對春雪桃基因DFR、LDOX、UFGT表達的影響
3 討論
果品色澤是果實商品價值的重要體現, 也是果實外觀品質的重要指標, 其形成是由于葉綠素降解, 同時形成顯現類胡蘿卜素或花色苷的結果[13]。研究表明, Ca肥能夠有效改善果皮著色[3], 果實成熟前施Ca肥, 可以顯著提高花青素含量; Mg肥可以提高果實可溶性糖含量[14], 降低酸度[15], 從而改善果實品質; 陳建等[16]研究得出, Mg2+可以與花色素螯合, 對花色苷具有穩定和保護作用。本試驗結果表明, K、Mg和Ca素處理提高了春雪桃果皮中花色苷含量, 降低了葉綠素含量, 改善了果實色澤;對果面色澤參數分析發現, 各處理在果實轉色期對果皮色度角h值均有一定影響, 果面色澤得到改善, 且Mg+Ca處理可以顯著促進果皮提前著色。這一結果與前人在荔枝[3]、葡萄[7,8]、蘋果[4]上的研究結果相一致。
花色苷是在糖代謝基礎上形成的, 類黃酮糖基轉移酶將糖基從活化的供體分子轉移到花色素上, 使不穩定的花色素轉變為穩定的花色苷[17], 在葡萄果實中, 只有糖分達到一定濃度時果實方能開始著色[18]; 利用蔗糖處理, 山楂果實花色苷合成酶基因上調表達, 促進花色苷的積累[19]。本研究結果表明, 隨著果實可溶性固形物含量的增加, 果皮中花色苷不斷積累; 不同噴肥處理可能通過促進果實中糖類積累, 增強類黃酮糖基轉移酶活性進而促進花色苷的生物合成, 改善果實色澤。二氫黃酮醇還原酶、無色花色素雙加氧酶、類黃酮糖基轉移酶在果實著色過程中發揮重要作用。結構基因DFR、LDOX的上調表達能夠為花色苷生物合成提供更多的底物物質, 與花色苷含量的增加密切相關[20]。Jeong等[21]研究表明, 花色苷合成途徑中關鍵結構基因VvUFGT的表達與葡萄果皮中總花色苷的積累呈現高度正相關性。本試驗中不同噴肥處理上調了春雪桃果實花色苷合成結構基因DFR、LDOX、UFGT的表達, 果皮中花色苷含量增加, Ca與Mg+Ca處理果實著色關鍵基因UFGT的表達量顯著高于對照。這一結果為K、Ca和Mg素營養提高桃果皮著色提供理論支持。
4 結論
葉面噴施K、Ca、Mg肥料改善了春雪桃果實著色,提高了果實品質, 促進了著色相關基因的表達, 其中以Ca與Mg+Ca處理效果最佳。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
