蘋果矮化砧木1年生枝解剖結構
2018-11-01 01:19:34李春燕楊廷楨王淑婷蔡華成高敬東杜學梅弓桂花杜海燕
農學學報 2018年10期
李春燕,楊廷楨,王淑婷,王 騫,蔡華成,高敬東,杜學梅,弓桂花,杜海燕
(山西省農業科學院果樹研究所/果樹種質創制和利用山西省重點實驗室,太原030031)
0 引言
蘋果(Malus pumilaMill.)矮化砧在全球蘋果生產中已被廣泛應用,但致矮機理還不清楚。國內外眾多學者分別從形態解剖結構、水勢、物質運輸、酶活性、激素調節等方面對蘋果砧木致矮機理進行了研究[1-7]。生物學原理告訴人們,結構決定功能,因此,從解剖結構研究蘋果砧木致矮機理很有意義。生產上利用蘋果矮化砧木栽培,一般選蘋果矮化砧木的1年生枝條作為嫁接砧段,所以弄清蘋果矮化砧木1年生枝的解剖結構是從解剖結構研究蘋果矮化砧木致矮機理的基礎。
前人主要從材皮比、導管、篩管、髓部對蘋果砧木1年生枝的解剖結構與矮化性的關系進行了研究[1-7],研究結論大多一致,表明,蘋果砧木枝條的材皮比,導管數量、導管密度、導管占木質部的比率和樹體生長勢呈正相關[1-5],篩管、髓部面積的大小與樹體生長勢沒有顯著相關性[1-2]。
但除了導管、篩管外,管胞和伴胞也是物質運輸的主要通道,在柑橘、梨上的研究表明,髓部、木栓形成層也可能是砧木致矮的因素[1,8-9],且植物莖的各組織在生長發育及物質運輸代謝過程中密切相關。因此,為了更加深入精確地研究蘋果矮化砧木1年生枝解剖結構與矮化性的關系,對蘋果砧木1年生枝的解剖結構進行辨析很有必要。目前,有關蘋果1年生枝組織解剖結構詳細深入的研究未見報道。本研究以Y-1蘋果矮化砧木為試材,運用常規石蠟切片法制片,通過顯微鏡觀察辨析其1年生枝條各組織的解剖結構,以期為更加深入精確地研究蘋果矮化砧木1年生枝解剖結構與矮化性的關系提供理論依據與參考。
1 試材與方法
1.1 試驗時間、地點
采樣于2015年10月下旬在山西省農科院果樹研究所砧木課題組砧木資源圃進行,石蠟切片制作于同年11月初在山西省農科院果樹研究所實驗室進行。
1.2 試驗材料
選擇生長健壯、無病蟲害的Y-1蘋果矮化砧木植株為試材。
1.3 試驗方法
在供試植株營養生長停止,頂芽形成,組織成熟后,取樹體北方位、中間部位、由剪口芽萌發的1年生發育枝,在中部取1 cm的枝段,然后以FAA固定液固定。枝樣用常規石蠟切片法制片,分橫切面和縱切面,用型號BX510 Olympus顯微鏡配置的型號DP71相機進行顯微攝影。
2 結果與分析
2.1 大體結構
對石蠟切片顯微觀察發現,蘋果矮化砧木Y-1 1年生枝的大體結構(圖1A)和大多雙子葉植物莖結構一樣,包括周皮(圖1A a)、皮層(圖1A b)、維管系統、髓(圖1A f、圖1B a)和髓射線(圖1B e),其中木質部和韌皮部有初生結構和次生結構,由外到內依次為:木栓層(圖1C a、圖1D a)、木栓形成層(圖1C b、圖1D b)、栓內層(圖1C c、圖1D c)、皮層(圖1E a、圖1F a)、初生韌皮部(圖1G d、圖1H d)、次生韌皮部(圖1G b、圖1H b)、形成層(圖1G a、圖1H a)、次生木質部(圖1B b)、初生木質部(圖1B c)、髓部,韌皮射線(圖1G i)、木射線(圖1B d)、髓射線貫穿其中。木栓層、木栓形成層和栓內層共同組成周皮,次生韌皮部、初生韌皮部、形成層、次生木質部、初生木質部及其貫穿其中的射線共同組成維管系統[10-16]。Y-11年生枝橫斷面各組織厚度依次為周皮0.0667 mm,皮層0.0426 mm,木質部1.4879 mm,韌皮部0.1879 mm,髓部半徑0.2845 mm,枝條半徑2.0696 mm。
2.2 詳細結構
2.2.1 周皮(圖1C、圖1D)周皮是一種次生保護組織,來源于初生結構的表皮,隨著枝的加粗生長,周皮逐漸代替了表皮,起保護作用;周皮通常在莖生長的第1年即出現,以后可繼續加厚[17];因此,蘋果矮化砧木Y-1 1年生枝應該已形成周皮。構成周皮的木栓層和栓內層來源于木栓形成層,木栓形成層進行平周分裂,向外衍生的細胞分化成木栓層,向內形成栓內層。木栓層細胞為木栓化的死細胞,呈磚形,排列整齊,缺乏細胞間隙,蘋果矮化砧木Y-1 1年生枝的木栓層有2~3層細胞,被番紅染成深紅色或棕色。木栓形成層(圖1C b、圖1D b)的細胞類似維管形成層,呈現高度液泡化,細胞間隙較小[17]。栓內層細胞(圖1C c、圖1D c)為生活細胞,由常含葉綠體的薄壁細胞組成,細胞間隙比木栓形成層大[17-18]。
2.2.2 皮層(圖1E、圖1F)由圖1E和圖1F可看出,蘋果矮化砧木Y-1 1年生枝皮層(圖1E a、圖版1F a)位于周皮和維管束之間,主要由多層薄壁細胞構成,具有明顯的細胞間隙,靠近周皮處有數層厚角組織(圖1E b、圖1F b),內含葉綠體[17];和栓內層相比,皮層細胞的形狀與排列明顯不同,細胞較大且排列疏松。
2.2.3 維管柱(圖1G、圖1H、圖1I、圖1J)皮層以內的部分稱為維管柱,它包含維管束、髓和髓射線。維管組織呈束狀,且各束在皮層的內側呈環狀排列。每個維管束由木質部、束內形成層(圖1G a、圖1H a)、韌皮部組成,次生木質部和次生韌皮部均來源于束內形成層,束內形成層進行切向分裂,向外產生次生韌皮部,向內產生次生木質部,同時還產生徑向排列的次生韌皮射線(圖1G i)和次生木射線(圖1G h)。維管束與維管束之間的薄壁細胞是髓射線,來源于束間形成層,它內連髓部,外通皮層,在橫斷面上呈輻射狀,有貯藏養料和橫向運輸的功能。
(1)韌皮部。莖韌皮部包含初生韌皮部和次生韌皮部,二者的組成成分基本相同,均由篩管、伴胞、韌皮薄壁細胞和韌皮纖維組成,不同的是,次生韌皮部中具有初生韌皮部中沒有的薄壁細胞——韌皮射線。次生木質部的韌皮射線細胞(圖1G i)成束排列,容易分辨,而初生韌皮部中的韌皮薄壁細胞在顯微條件下較難分辨。篩管和伴胞是植物輸導組織的組成分子,起運輸同化產物的作用。篩管分子在橫切面上為多角形,但衰老的篩管及一些韌皮薄壁細胞常被擠毀[18](被擠毀的衰老的篩管見圖1G j),而產生新的篩管分子。伴胞是一種特殊類型的薄壁組織細胞,一個篩管分子常結合一個或幾個伴胞[15],且可以發生在篩管分子各個方向[19];在橫切面上,伴胞比篩管分子小,多呈三角形、方形或梯形[17,19-20];伴胞的原生質濃厚[13],和篩管相比染色較深(見圖G f)。初生韌皮部中的韌皮纖維在靠近皮層處成束分布形成一圈,其細胞是一種厚壁組織細胞,橫切面呈圓形、長圓形或多角形,細胞壁很厚[10]。次生韌皮部和初生韌皮部之間的界線是不太明顯的,一般說來,次生韌皮部排列整齊。在木本植物的次生韌皮部中,有功能的韌皮部通常限于當年產生的生長層[17],因此,研究砧木枝條篩管的解剖結構與致矮能力的關系需選用接近形成層次生木質部的篩管。
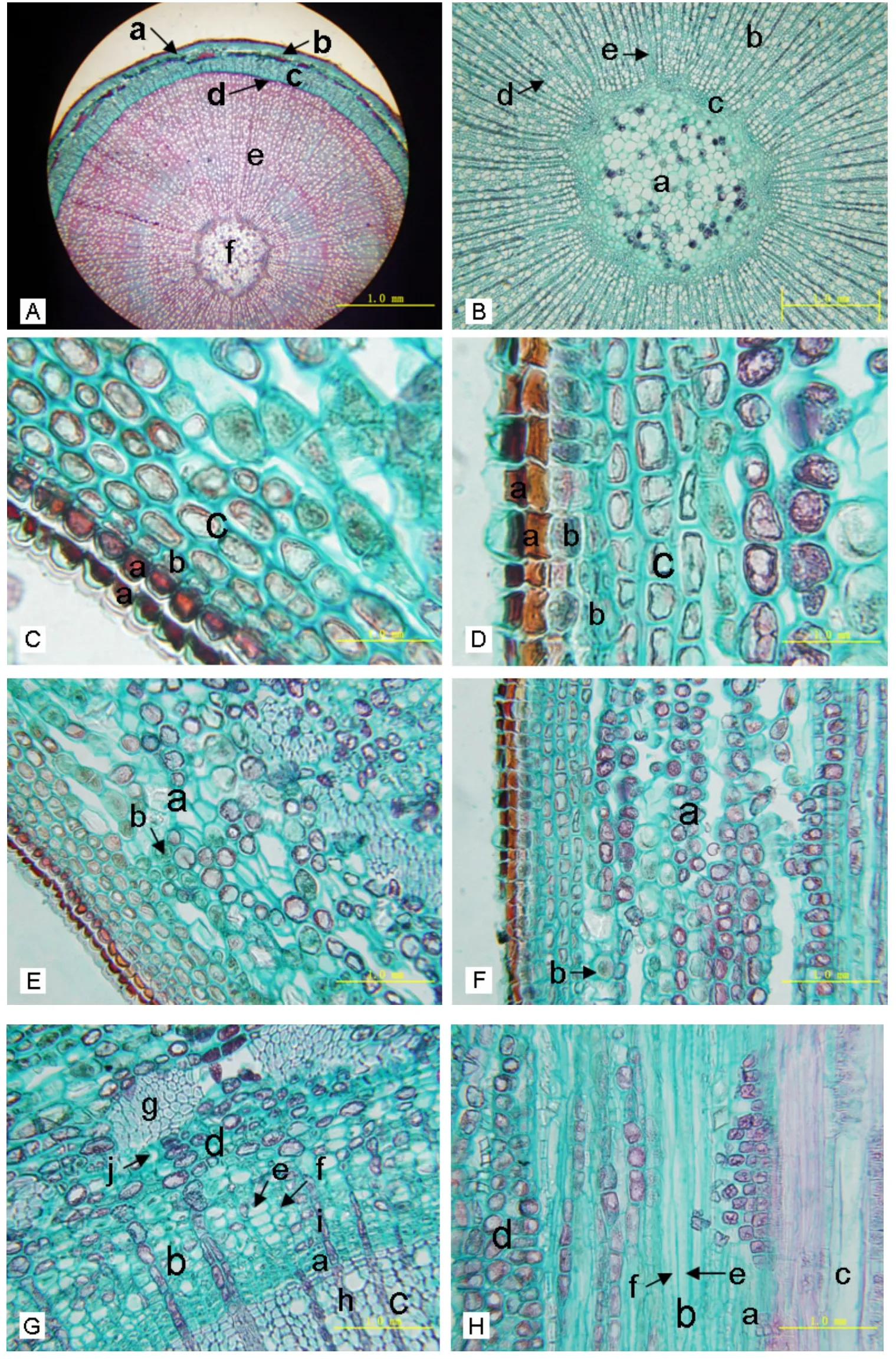

圖1 蘋果矮化砧Y-1 1年生枝解剖結構
(2)木質部(圖1I、圖1J)。形成層以內、髓部以外為木質部。蘋果1年生枝的木質部包含初生木質部和次生木質部。次生木質部和初生木質部均由導管(圖1I a、圖1J a)、管胞、木纖維和木薄壁細胞組成。蘋果矮化砧木Y-1 1年生枝的維管柱和大多木本植物一樣,木質部占大部分,其中的導管和管胞起向上運輸水分和無機鹽的作用。導管比較容易辨認,因為它孔徑比其他細胞明顯大。管胞呈紡錘形單細胞,細胞壁木質化,橫切面呈三角形、方形或多角形,常集合分布在木質部[20]。纖維在橫切面上呈圓形、長圓形或多角形,成群或束狀分布,且常比在同一木材中的管胞壁厚[21]。在木質部中還存在一種管胞和纖維中間狀態的細胞,即纖維管胞,纖維管胞、纖維、管胞這3種細胞在顯微結構下不易分辨[17,22],所以,和矮化關系密切的管胞在顯微結構下不易辨別。木薄壁組織細胞含有不同的后含物,常見的有淀粉和脂肪,具有貯藏營養的功能[17]。木射線(圖1I b)由射線原始細胞衍生,細胞壁常木質化[17-18]。
(3)髓部(圖1B)。蘋果砧木Y-1 1年生枝的蘋果莖的髓射線(圖1B e)很窄,只有1-2列細胞。髓位于莖中央,由薄壁細胞組成,具有細胞間隙,常貯有豐富養料。
3 結論與討論
3.1 大體結構
本研究表明,蘋果矮化砧木Y-1 1年生枝的大體結構包括周皮、皮層、維管系統、髓和髓射線,其中木質部和韌皮部有初生結構和次生結構,由外到內依次為:木栓層、木栓形成層、栓內層、皮層、初生韌皮部、次生韌皮部、形成層、次生木質部、初生木質部、髓部,韌皮射線、木射線、髓射線貫穿其中。初生韌皮部和次生韌皮部組織界線不明顯。
3.2 導管、管胞、篩管
蘋果1年生枝次生維管組織處在初生韌皮部和初生木質部之間,其向內向外的擴張使莖的內部產生相當大的壓力,這種壓力使得原生木質部和衰老的篩管及一些韌皮薄壁細胞常漸漸被擠毀,而這些初生木質部導管通常失去了輸導作用,衰老的篩管也被新生篩管分子所更新,且篩管的輸導作用常只能維持1~2年。所以,要研究具有輸導作用的莖組織器官應以接近形成層、當年產生的生長層為宜。另外,導管由于周圍的薄壁細胞從紋孔處擠入其腔內,形成侵填體而被堵塞,失去輸導功能[18];篩管原生質中往往含有P-蛋白,這種P-蛋白可以迅速而可逆地從凝縮狀態轉變為分散狀態,在此過程中它們堵塞篩管,從而控制篩管的輸導能力[17]。所以,結合本研究可以認為,在顯微結構下,與矮化關系密切的導管容易辨別,管胞難以辨別,篩管、伴胞可以辨別,但對具有輸導能力的導管、篩管需用更精細和先進的技術方法進行分辨研究。
3.3 枝條組織解剖結構與致矮
在蘋果接穗品種和砧木本身枝條上的研究表明,枝條材皮比越大,樹體生長勢越強[3,23];蘋果砧木1年生枝材皮比在2~3為矮化,>3為喬化,比值越大,樹體越喬化[1]。枝條導管的密度、總面積、孔徑越小,數量越少,占木質部比例越小,樹體生長勢就越弱[2]。在梨上的研究則認為,砧木致矮是由于砧段的木栓形成層細胞層數多,使皮部的篩管分子在增粗的過程中承受更大的壓力,從而被擠壓變形、萎縮、破壞或消失,阻礙了碳水化合物向基部的運輸,導致樹體矮化[9]。另外,在蘋果上的研究認為,髓部及篩管的面積與砧木植株的體積無相關性[1],而在柑橘上的研究與之相反,得出柑橘樹體從矮化到喬化,髓部的面積呈增大趨勢[8]。
筆者分別對M系與Y系蘋果矮化砧木解剖結構進行了研究,表明:蘋果砧木1年生枝木質部從近皮部到近髓部,導管密度大多呈增大趨勢,導管面積大多呈減小趨勢;M系蘋果砧木1年生枝的周皮厚度、皮層厚度、髓部半徑、枝條半徑,周皮、皮層、髓部、枝條的面積,篩管面積均顯著小于喬化砧木,而木質部占橫斷面積的百分比、近皮部導管密度、導管平均密度、近皮部導管面積/篩管面積、導管平均面積/篩管面積均顯著大于喬化砧木。得出,M系蘋果矮化砧木致矮的關鍵部位可能是韌皮部(主要是篩管)。Y系蘋果矮化砧木1年生枝髓部半徑及面積、近皮部導管面積、導管平均面積、木質部厚度/韌皮部厚度、材皮比均顯著小于喬化砧木,而近皮部導管密度、韌皮部厚度/枝條半徑、韌皮部占橫斷面積的百分比均顯著大于喬化砧木;Y系蘋果矮化砧木嫁接品種1年生枝髓部半徑、近皮部導管面積/篩管面積、中部導管面積/篩管面積、近髓部導管面積/篩管面積、導管平均面積/篩管面積顯著小于喬化砧木。得出:Y系蘋果矮化砧可使接穗品種髓部變大,導管面積/篩管面積變小,Y系蘋果矮化砧木致矮關鍵部位可能是木質部(主要是導管)。從M系和Y系的研究結論可以看出,矮化砧限制了髓部的生長;砧木不同,可能致矮機理也不同。
今后應運用更先進的技術對蘋果矮化砧木1年生枝的各組織解剖結構與喬化砧木進行更準確精細的比較,以找出更多與矮化有關的組織特征,從枝條組織解剖結構對蘋果矮砧致矮機理進行更深入的探討。
