小鼠早期胚胎CSD基因及牛磺酸的表達研究*
2018-10-31 07:50:18范晶晶呂釗旭馮永珍鄭秋闿
重慶醫(yī)學 2018年30期
關鍵詞:小鼠
范晶晶,李 煒,呂釗旭,馮永珍,鄭秋闿
(1.濰坊學院生物與農(nóng)業(yè)工程學院,山東濰坊 261061;2.山東省高校生物化學與分子生物學重點實驗室,山東濰坊 261061;3.濰坊護理職業(yè)學院內(nèi)科教研室,山東濰坊261041;4.山東省濰坊市婦幼保健院新生兒科 261000;5.濰坊學院校醫(yī)院護理部,山東濰坊 261061;6.濰坊學院化學化工與環(huán)境工程學院,山東濰坊 261061)
半胱亞磺酸脫羧酶(cysteine sulfinate decarboxylase,CSD)對底物半胱亞磺酸具有較高親和性,是牛磺酸合成的限速酶[1]。在哺乳動物體內(nèi)牛磺酸合成的第一步是由蛋氨酸和半胱氨酸經(jīng)代謝產(chǎn)生半胱亞磺酸,然后半胱亞磺酸經(jīng)過CSD的脫羧作用形成亞牛磺酸,再經(jīng)氧化最終生成牛磺酸[2]。研究發(fā)現(xiàn)牛磺酸是輸卵管液中水平最高的游離氨基酸,在小鼠輸卵管液中可達到6.64 μmol/L,能占總游離氨基酸的59%[3],在妊娠第二天大鼠輸卵管液中濃度達101.8 μmol/L[4]。輸卵管是胚胎早期發(fā)育的主要場所,許多研究結(jié)果已證實牛磺酸確實參與調(diào)節(jié)胚胎的發(fā)育和胚胎的著床。在培養(yǎng)液中添加一定量的牛磺酸能促進小鼠、大鼠、兔、牛和人等多種動物胚胎的體外發(fā)育[4-8]。但是目前早期胚胎是否表達CSD,胚胎能否通過CSD的作用來自身合成牛磺酸滿足發(fā)育的需要,至今仍無詳細的研究。因此,本實驗采用反轉(zhuǎn)錄PCR(RT-PCR)、免疫熒光染色的方法,研究分析牛磺酸、CSD基因及蛋白在小鼠早期胚胎中的表達情況,進而為CSD和牛磺酸在胚胎早期發(fā)育功能研究奠定基礎。
1 材料與方法
1.1動物與試劑 10周齡雌、雄昆明(KM)品系小鼠(購自青島市實驗動物中心,SCXK(魯)20130010),自由飲水采食,溫度控制在22~26 ℃,14 h光照,10 h黑暗。小鼠超數(shù)排卵和早期各發(fā)育階段胚胎的收集方法按照范晶晶[9]的描述進行。Trizol RNA提取試劑盒(美國Invitrogen公司),M-MLV 反轉(zhuǎn)錄試劑盒(美國Promega公司),孕馬血清激素(PMSG)、人絨毛膜促性腺激素(hCG,寧波第二激素廠),F(xiàn)ITC標記鏈霉親和素(SP-FITC,美國Southern Biotech公司),生物素標記羊抗兔IgG(GARB)、FITC標記山羊抗兔IgG(GAR-FITC,Zymed公司),牛磺酸一抗(德國Merck公司),CSD一抗是由法國TAPPAZ教授(Directeur de Recherche CNRS,INSERM U 433,法國)贈送。其余試劑購自美國Sigma公司。主要儀器有高速冷凍離心機(德國Eppendorf公司),凝膠成像系統(tǒng)(美國Alpha Imager公司),TE2000-E激光共聚焦掃描顯微鏡(日本Nikon公司)。
1.2RT-PCR 分別收集各發(fā)育時期100枚胚胎于500 μL的Trizol RNA裂解液中,按照說明書步驟提取總RNA,進行反轉(zhuǎn)錄。取3 μL反轉(zhuǎn)錄產(chǎn)物加至PCR反應體系中,引物各1 μL,10×buffer 5 μL,dNTP 4 μL,Taq酶1 μL,H2O 35 μL,反應總體積為50 μL。PCR反應條件為:94 ℃ 5 min;94 ℃ 30 s,52 ℃ 20 s,72 ℃ 30 s,35個循環(huán);循環(huán)結(jié)束后72 ℃ 10 min。CSD上游引物為5′-GTT TGG GAT TGT TGT AGA TGA-3′,下游引物為5′-GTG TAC TGG CTA GTG TTG AGG-3′,預計擴增片段長度為279 bp(登錄號 NM_144942)。β-肌動蛋白(β-actin)作為內(nèi)參,上游引物為5′-ATG AGG TAG TCT GTC AGG T-3′,下游引物為5′-ATG GAT GAC GAT ATC GCT-3′,預計擴增片段長度為569 bp(登錄號X03672)。擴增PCR產(chǎn)物用0.8%瓊脂糖凝膠電泳分離,Alpha Imager凝膠成像系統(tǒng)拍照分析,Alpha Imager 2200 軟件分析PCR電泳條帶的光密度值。
1.3免疫熒光染色 取2-細胞、4-細胞、8-細胞、桑椹胚、囊胚期胚胎用磷酸鹽緩沖液(PBS)沖洗,然后轉(zhuǎn)移到載玻片上,晾干后迅速用-20 ℃預冷的固定液(甲醇∶丙酮=1∶1)固定30 min。封閉液(0.2%Triton X-100、10%正常羊血清的PBS)封閉2 h,加入CSD一抗(1∶1 000)在4 ℃下孵育過夜。次日PBS洗3次,每次5 min,加入GARB(1∶200)在常溫下育孵2 h,然后用PBS洗滌,加入SP-FITC(1∶50)常溫下孵育2 h,再用上述方法進行洗滌,用Hoechst33342復染2 min。另外一部分固定好的胚胎封閉后加入牛磺酸一抗(1∶1 000),4 ℃下孵育過夜,加入GAR-FITC(1∶200)在常溫下育孵2 h,加入碘化丙啶(PI)(1∶200)復染2 min,用1,4-二疊氮雙環(huán)[2.2.2]辛烷(DABCO)封片,激光共聚焦顯微鏡觀察。

2 結(jié) 果
2.1RT-PCR結(jié)果 CSD mRNA在2-細胞、4-細胞、8-細胞、桑葚胚、囊胚期的RT-PCR檢測結(jié)果顯示,著床前不同時期小鼠胚胎都有CSD mRNA的表達,擴增的PCR產(chǎn)物與預期的大小一致,約279 bp,β-actin約569 bp。CSD mRNA在小鼠著床前胚胎中的相對表達強度顯示,胚胎從2-細胞期開始到囊胚期CSD mRNA表達量呈逐漸升高趨勢,桑葚胚表達量急劇升高,囊胚期達到最高值。見圖1。
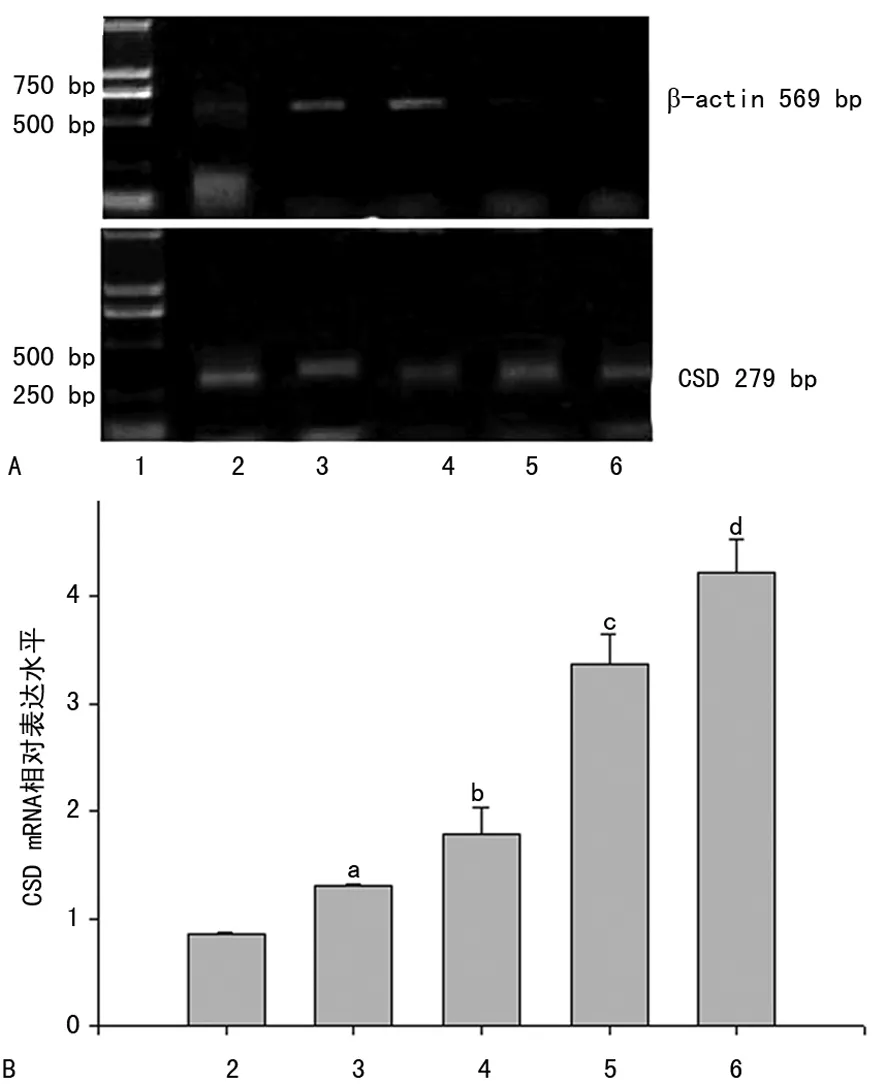
A:PCR凝膠電泳圖;B:PCR分析圖。1:DNA分子標記物;2:2-細胞期;3:4-細胞期;4:8-細胞期;5:桑葚胚期;6:囊胚期;a:P<0.95,與2-細胞期比較;b:P<0.05,與4-細胞期比較;c:P<0.05,與8-細胞期比較;d:P<0.05,與桑葚胚期比較
圖1 RT-PCR檢測CSD mRNA在早期胚胎
發(fā)育各階段的表達
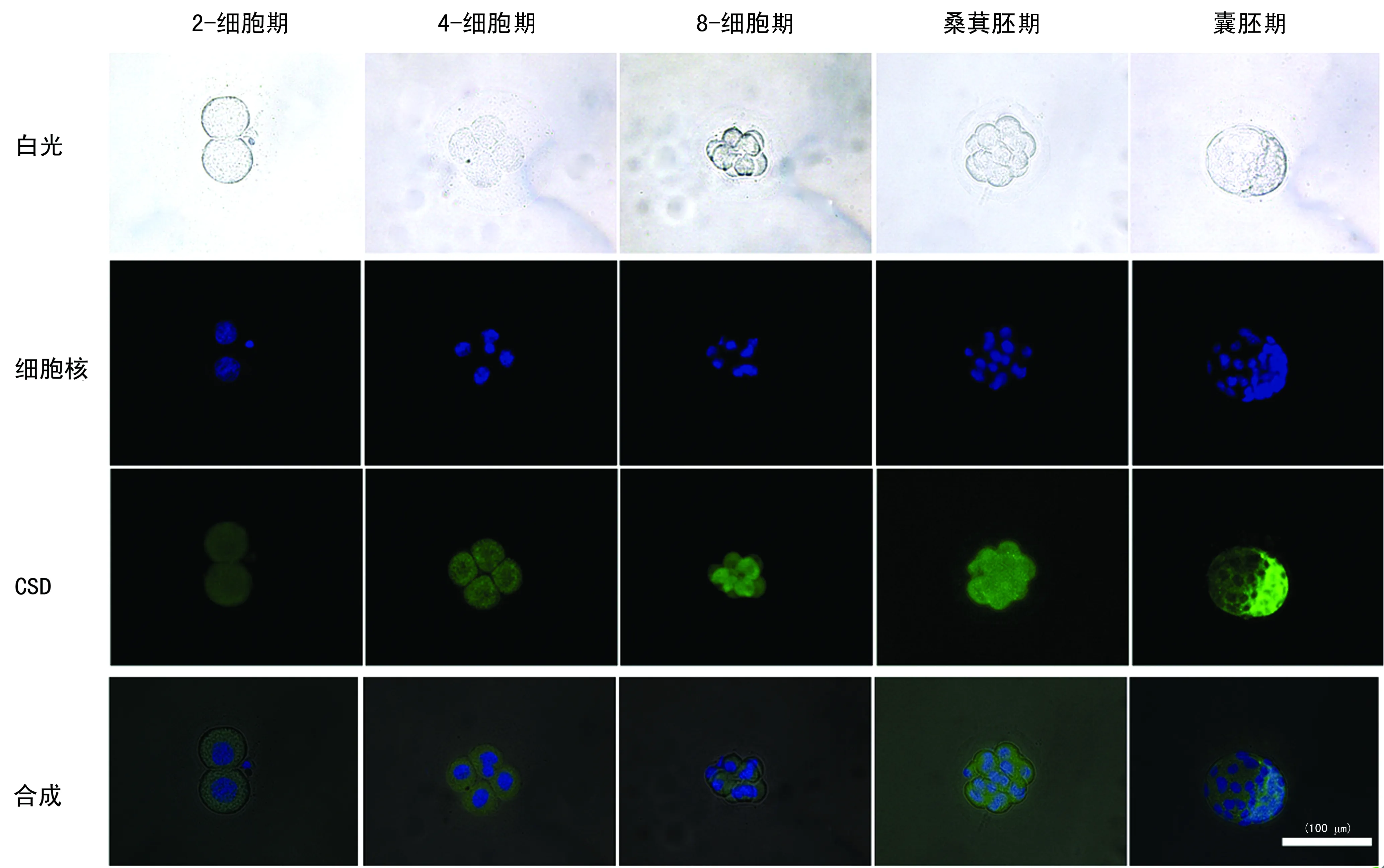
圖2 CSD蛋白在小鼠早期胚胎發(fā)育各階段的表達(免疫熒光染色)
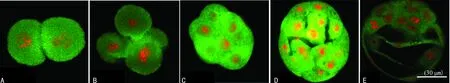
A:2-細胞期;B:4-細胞期;C:8-細胞期;D:桑葚胚期;E:囊胚期
圖3牛磺酸在小鼠早期胚胎發(fā)育各階段的表達(免疫熒光染色)
2.2免疫熒光染色結(jié)果 CSD蛋白免疫熒光染色結(jié)果顯示,從2-細胞至囊胚期均呈陽性綠色熒光,熒光亮度逐漸增強。CSD均勻分布在卵裂球細胞質(zhì)中,晚期囊胚的內(nèi)細胞團和滋養(yǎng)層細胞都為陽性著色,透明帶無著色,見圖2。牛磺酸免疫熒光染色結(jié)果顯示,從2-細胞期至囊胚期的整個發(fā)育過程細胞質(zhì)均呈陽性綠色熒光,透明帶無陽性,見圖3。
3 討 論
小鼠妊娠期短、繁殖力強多被用來研究胚胎的發(fā)育。哺乳動物早期胚胎發(fā)育是一個復雜有序的調(diào)控過程,任何因素異常都有可能導致發(fā)育失敗[10]。牛磺酸是調(diào)節(jié)機體多個系統(tǒng)功能的重要活性物質(zhì)[11-12]。近年研究發(fā)現(xiàn)在雌性動物生殖系統(tǒng)含量豐富,對胚胎的生長發(fā)育具有重要的促進作用[13]。例如可提高受精卵的發(fā)育速度[4],顯著提高胚胎2-細胞率、囊胚率、囊胚總細胞數(shù)[5,14]。牛磺酸促進胚胎發(fā)育的機制主要有:(1)調(diào)節(jié)滲透壓穩(wěn)定細胞膜[15];(2)可以抑制氧化性毒素物質(zhì)的產(chǎn)生,并通過調(diào)節(jié)線粒體活性起到抗氧化作用[16];(3)有類胰島素樣作用,能促進細胞增殖[17];(4)可以抑制Na+,K+-ATP酶活性,避免細胞內(nèi)K+水平異常升高所造成胚胎的損傷等[18]。
CSD在嚙齒類動物體內(nèi)的活性較高,其自身合成的牛磺酸足以滿足生理需要。CSD基因敲除后,第二代(G2)小鼠大部分在出生后24 h死亡,給G2代鼠飲水中飼喂牛磺酸可以使G3代鼠的成活率從15%提高到92%[19]。李建華[20]報道在小鼠輸卵管上皮細胞中有CSD的表達,并且在輸卵管上皮細胞也檢測到了牛磺酸[21],說明輸卵管上皮細胞可以通過CSD途徑合成并分泌牛磺酸到輸卵管液中,從而調(diào)節(jié)早期胚胎的發(fā)育。KIM等[12]也報道,在ICR品系小鼠4.5、10.5、15.5、18.5 d胚胎中都能檢測到CSD mRNA的表達。另外,對小鼠卵巢進行CSD免疫組織化學染色,結(jié)果發(fā)現(xiàn)從初級卵泡到三級卵泡的卵母細胞都有CSD蛋白表達[20]。本研究結(jié)果發(fā)現(xiàn)從小鼠2-細胞開始一直到囊胚的整個發(fā)育過程都能檢測到CSD的表達,說明胚胎自身也可以通過CSD來合成牛磺酸。結(jié)果提示著床前后胚胎中牛磺酸不僅可以由輸卵管液提供,也可以通過自身來合成一部分牛磺酸。同時早期胚胎也一直有牛磺酸表達,說明牛磺酸確實參與了胚胎的卵裂、細胞的發(fā)育和分化過程。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
