不同來源堆肥腐殖質還原菌異化鐵還原能力評估與調控
2018-10-29 04:22:40趙昕宇范鈺瑩席北斗檀文炳何小松
中國環境科學 2018年10期
趙昕宇,范鈺瑩,3,席北斗*,檀文炳,何小松,張 慧,李 丹
?
不同來源堆肥腐殖質還原菌異化鐵還原能力評估與調控
趙昕宇1,2,范鈺瑩1,2,3,席北斗1,2*,檀文炳1,2,何小松1,2,張 慧1,2,李 丹1,2
(1.中國環境科學研究院環境基準與風險評估國家重點實驗室,北京100012;2.中國環境科學研究院國家環境保護地下水污染模擬與控制重點實驗室,北京 100012;3.東北農業大學,黑龍江哈爾濱 150030)
通過富集不同來源堆肥過程中的腐殖質還原菌,并分析比較其異化鐵還原能力差異,發現其電子轉移能力從大到小依次為:蛋白類>纖維素類>木質素類.相關性分析表明,和是極顯著影響異化鐵還原的腐殖質還原菌屬.利用冗余分析探究關鍵腐殖質還原菌與堆肥過程微環境因子的響應關系,結果發現可溶性有機氮是影響這些關鍵腐殖質還原菌變化的主要微環境因素.在此基礎上,基于堆肥微環境因子與關鍵腐殖質還原菌菌群結構之間的響應關系,提出一種促進異化鐵還原相關的腐殖質還原菌生長的調控方法.本研究可以深入了解堆肥中影響腐殖質還原菌群落的關鍵因素,而且對于環境中污染物生物地球化學循環也具有重要的生態學意義.
腐殖質還原菌;16S rDNA;異化鐵還原;冗余分析;堆肥
微生物可通過調節特定的生物化學途徑,降解或還原有機、無機污染物[1-2].這在污染物原位修復、污水處理以及生物燃料提取等方面呈現出不可替代的優越性和重要的應用前景,電子轉移是新陳代謝的基礎,地球所有能量都來源于氧化還原過程.腐殖質呼吸是近年來新發現的可參與環境修復生物降解過程的能量代謝方式,能夠進行腐殖質呼吸的微生物稱之為腐殖質還原菌[3],腐殖質還原菌作為一個龐大且復雜的生物類群,對重金屬及有機污染物的降解起到重要作用,腐殖質還原菌是以AQDS或腐殖質作電子受體時進行胞外電子傳遞的一類微生物,腐殖質和腐殖質還原菌之間的這種電子穿梭被認為是促進重金屬與有機污染物降解轉化的主要原因[4-6].腐殖質還原菌還原腐殖質的生物化學途徑與腐殖質還原菌還原Fe(III)的生化機制很相似[7-8],并且大多數的腐殖質還原菌都具有還原鐵的功能[9-11].
腐殖質還原菌普遍存在于自然環境中,學者們已經在土壤、泥炭地、污泥、湖泊沉積物、河流沉積物、海洋沉積物以及水體等環境介質中分離富集出多種具有腐殖質呼吸功能的微生物[12].并且在許多極端環境下,也分離出多種具有電子傳遞能力的微生物,例如嗜熱菌、嗜酸菌和嗜堿菌等[13].按照對氧氣的需求不同,腐殖質還原菌又可以分為兼性厭氧菌和嚴格厭氧菌,主要集中在以下3個門: Proteobacteria、Acidobacteria與Firmicutes[12].堆肥是一個以微生物為驅動力,大量合成腐殖質的生物化學過程.腐殖質的形成可能誘導堆肥過程中腐殖質還原菌的生長,并且該過程中微環境因子的變化可能會影響腐殖質還原菌的群落組成與活性[14-15].然而,對于不同來源物料在堆肥過程中腐殖質還原菌的鐵還原能力差異及其影響因素卻鮮有報道.并且,現如今從自然環境中分離的腐殖質還原菌僅占可培養微生物的0.1%.
本研究主要目的為:探究不同物料堆肥過程腐殖質還原菌對異化鐵還原電子轉移能力差異;篩選具有高效異化鐵還原能力的腐殖質還原菌;探究其關鍵影響因素,并提出一種促進篩選出的腐殖質還原菌生長的調控方法.
本研究主要探究6種物料:雞糞(CM)、牛糞(DCM)、果蔬(FVW)、雜草(WW)、秸稈(SW)及枯枝(GW)在堆肥過程中腐殖質還原菌的電子轉移能力及其影響因素,并構建一種可促進腐殖質還原菌生長的微環境調控方法.為堆肥物料中的污染物降解或應用富含腐殖質堆肥產品修復污染土壤提供參考.
1 材料與方法
1.1 主要儀器及實驗設計
實驗樣品采集、堆肥實驗搭建詳見前期文章[11].采集階段為升溫期(CM1、DCM1、FVW1、WW1、SW1和GW1)、高溫期(CM2、DCM2、FVW2、WW2、SW2和GW2)和腐熟期(CM3、DCM3、FVW3、WW3、SW3和GW3).
有機質(organic matter, OM)采用灼燒法測定;含水率采用馬弗爐測燒失量法測定,在105℃下灼燒6h后測量燒失量;可溶性有機碳(Dissolved organic carbon, DOC)、可溶性有機氮(Dissolved organic nitrogen, DON)均采用島津TOC-VCPH有機碳分析儀測定;總氮采用凱氏消煮法測定;NH4+-N采用納氏比色法測定;NO3--N采用分光光光度計法測定.以上指標的測定結果曾發表于先前研究中,詳見文章[16].
1.2 腐殖質還原菌培養及異化鐵還原試驗
腐殖質還原菌培養采用富集培養基(BAS培養基),具體成分詳見前期文章[16].
6種物料經5mm篩,用去離子水沖洗,沉降30min后棄上層混濁液,取下層泥漿,重復多次再使用.取5mL堆肥泥漿于裝有50mL富集培養基的西林瓶中,室溫下80% N2和20% CO2充20min,蓋橡膠塞.為了使富集培養基環境與原始物料環境盡可能保持一致,調節CM1、CM2、CM3、DCM1、DCM2、DCM3、FVW1、FVW2、FVW3、WW1、WW2、WW3、SW1、SW2、SW3、GW1、GW2及GW3的pH值為8.0、8.8、8.6、8.0、8.9、8.4、8.1、8.7、8.2、6.7、8.9、8.4、8.0、8.8、8.6、6.2、7.7及8.2,分不同堆肥階段放置于不同溫度的厭氧培養箱(升溫期40℃,高溫期58℃及腐熟期43℃)中培養.3d后將其開蓋至上清液橙紅色褪去,8h內使AQDS還原率達到80%可視為腐殖質還原菌馴化成功.
分別將MR-1菌株[12](對照)及堆肥腐殖質還原菌在LB液體培養基中培養至對數生長期(約18h),培養溫度為30℃,然后將其在4℃下離心10min(8000r/min),傾去上清液.用已滅菌的BAS培養液洗滌菌體,再次離心,重復多次,最終用等體積BAS培養液重新制成菌懸液,以無菌液BAS培養基作空白對照,在紫外吸收波長為600nm下,將所有菌體濃度均調至0.18,以保證一致性.在100mL西林瓶中進行反應,配制20mmol/L Fe (III)-critrate溶液作電子受體,以5mmol/L乳酸鈉作電子供體,加入200 μL菌液,最終反應體系為50mL.室溫下用80% N2和20% CO2充20min,pH值調至7.4.將樣品搖勻后,用注射器取2mL,過0.22μm纖維素濾膜,采用鄰菲啰啉分光光度法于510nm測定濾液吸光度,計算Fe(II)濃度,每24h測定一次直到最終達到穩定.利用一級動力學方程進行擬合[17],求得不同堆肥中腐殖質還原菌還原Fe(III)的常數.方程的表達式:
Fe= Fe0[1-exp (-×)]
式中:Fe為時刻Fe(II)的生成量, mmol/L;Fe0為Fe(II)的潛在生成量, mmol/L;為培養時間,h;為反應速率常數,h-1.
1.3 DNA 提取及16SrDNA測序
使用E.Z.N.A.TM土壤DNA提取試劑盒(Omega Bio-tek,廣州)提取六種物料的DNA,使用Illumina MiSeq測序儀測序,采用合成測序法,對最終獲得clean數據進行OTU聚類分析和物種分類學分析.在物種分類學上,對于序列進行精細至分類學上屬的劃分.
1.4 數據分析
運用SPSS 19.0、Origin 8.0、Canoco for windows 4.5進行數據分析.
2 結果與討論
2.1 腐殖質還原菌異化鐵還原特征

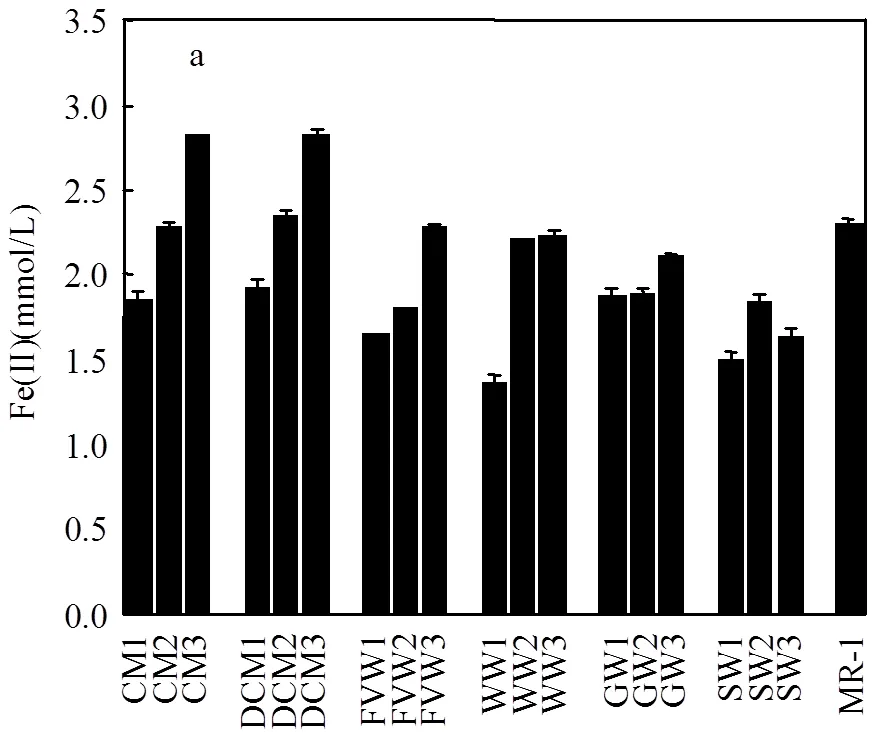
圖1 不同物料堆肥中腐殖質還原菌與S.oneidensis MR-1對Fe(III)還原的影響
為反映腐殖質還原菌還原腐殖質的能力,利用腐殖質還原菌與Fe(III)-citrate進行直接接觸反應,絕大部分腐殖質還原菌具有還原Fe(III)的功能[18], Fe(III)逐漸被腐殖質還原菌還原為Fe(II),以Fe(III)還原量間接代表腐殖質還原菌還原腐殖質的能力.如圖1所示,不同物料類型堆肥過程中腐殖質還原菌的還原能力呈現不同的變化趨勢.與相應對照組比較,不同來源堆肥過程中的腐殖質還原菌均能明顯還原Fe(III).
一級動力學方程可以較好的描述Fe(III)-citrate還原動態變化(表1),方程擬合的相關系數均達到顯著水平(<0.1).不同物料堆肥的3個階段的值變化存在差異,而在堆肥的升溫期,纖維素類物料(FVW、WW)反應速率顯著高于其它物料(<0.05).而蛋白類物料(CM、DCM)在高溫期的反應速率達到最高,至腐熟期反應速率仍保持較高水平.這說明不同物料的物質組成及結構對腐殖質還原菌還原異化鐵能力有不同的影響.堆肥初期腐殖質還原菌利用纖維素分解的單糖等營養物質進行生長代謝[19],異化鐵電子受體利用量增多,其還原能力隨之增強.而蛋白類物料中的營養物質豐富,且在高溫期的蛋白質大量降解[20],造成腐殖質還原菌代謝活動反應劇烈,反應速率隨之升高.
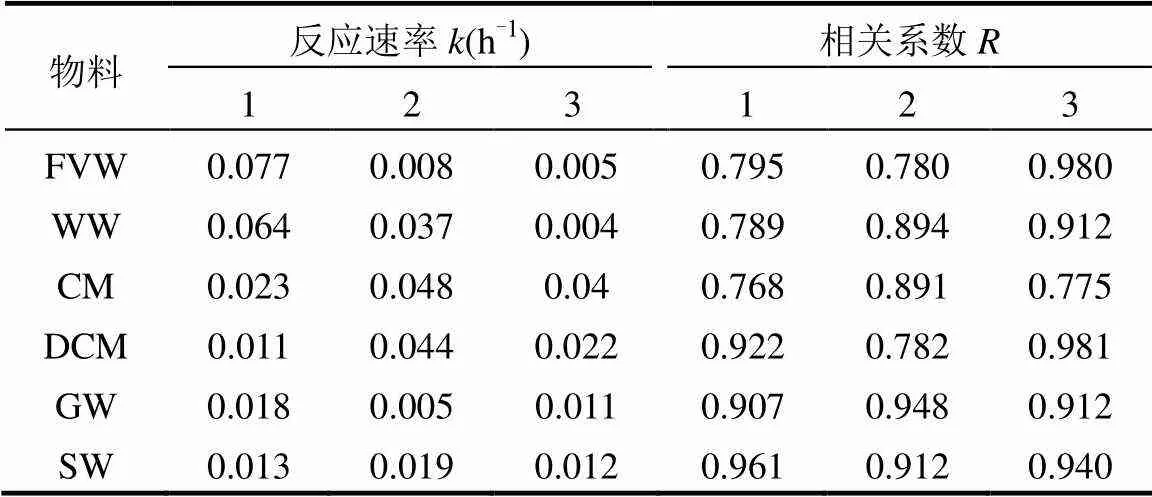
表1 Fe(III)還原能力的一級動力學方程擬合結果
與MR-1的還原能力相比(圖2a),堆肥腐熟期CM3、DCM3、FVW3、WW3、GW3及SW3中腐殖質還原菌的還原能力分別是模式菌株MR-1還原能力的1.22、1.23、0.99、0.97、0.92及0.71倍.由此可以說明,經過堆肥后,蛋白類物料(CM、DCM)中腐殖質還原菌電子轉移能力相比其他物料堆肥更強.CM、DCM、FVW及WW中腐殖質還原菌的還原能力隨堆肥過程呈明顯增加趨勢.這可能是由于蛋白類與纖維素類物料在初始階段較易降解,可相對快速地形成氧化還原功能基團,并影響腐殖質還原菌的電子轉移力.而與之相比,木質素類物料(GW、SW)中腐殖質還原菌的電子轉移能力相對較低,且隨堆肥過程變化不顯著.這主要是由于木質素類物料在堆肥過程中較難降解,腐殖質還原菌獲得營養物質相對較少,堆肥過程中形成氧化還原功能基團相對緩慢,導致期腐殖質還原菌電子轉移能力相對較弱.總之,不同堆肥中形成腐殖質的分子量及結構不同,腐殖質還原菌通過堆肥過程中形成的腐殖質做電子受體從而獲得自身生長的能力不同[21],進而影響堆肥中腐殖質還原菌的還原能力.圖2(b)可更為清晰的分析不同物料堆肥過程中腐殖質還原菌的電子轉移能力差異.從總體上看,腐殖質還原菌的電子轉移能力的變化量從大到小依次為CM>DCM>WW>FVW>GW>SW.因此,也可進一步證明,堆肥過程對蛋白類物料中腐殖質還原菌的提升能力最強,其次為纖維素類與木質素類.
2.2 腐殖質還原菌與異化鐵還原能力相關性分析
由于本研究中培養為富集培養,培養后為腐殖質還原菌菌群,因此哪種腐殖質還原菌顯著影響Fe(III)的還原尚不清晰,且還原的機制也尚不明確,因此,本研究采用相關性分析進一步篩選哪一種或幾種腐殖質還原菌對Fe(III)還原具有顯著作用.其中,不同物料堆肥過程中腐殖質還原菌的群落結構發表于前期研究[16]中.相關性分析結果如圖3,共有100個腐殖質還原菌菌屬與Fe(III)的還原呈正相關關系.由圖3中可以看出,腐殖質還原菌歸屬于4個菌門:Actinobacteria、Bacteroidetes、Firmicutes和Proteobacteria,大部分還原Fe(III)的腐殖質還原菌集中于Proteobacteria,其次為Firmicutes,而Actinobacteria和Bacteroidetes相對較少.在6種不同物料堆肥中,纖維素類物料FVW和WW中促進Fe(III)還原的相關菌屬分別為37個和39個,高于蛋白類物料CM(23個)和DCM(31個),而木質素類物料SW和GW中含有促進還原能力的腐殖質還原菌較少,分別為16個和8個.這可能由于纖維素類物料在堆肥前期不易分解,堆肥后期逐漸被纖維素菌分解為小分子物質,該類營養物質易被腐殖質還原菌吸收利用[19],微生物代謝加快,可加速堆肥過程中腐殖質還原菌電子轉移能力的馴化.堆肥腐熟期大量的腐殖質類物質累積,從而豐富了纖維素類物料中腐殖質還原菌的多樣性.相比于纖維素類物料,蛋白質類物料中腐殖質還原菌與異化鐵還原呈正相關的菌種略少,然而蛋白類物質的平均還原能力在6種物料中最強,這可能是由于蛋白類物質利于分解,腐殖質形成速度相對較快,腐殖質還原菌的氧化還原能力也相對較強[20].因此蛋白類物料經過堆肥馴化后,盡管促進Fe(III)還原相關菌屬的多樣性較低于纖維素類物料,但其還原異化鐵能力卻高于纖維素類物料.

菌屬序號按照圖中排列順序進行標號
*<0.05; **<0.01
圖3中可以看出,Actinobacteria門的(序號94)與Firmicutes門的(序號59)和(序號73)都與還原Fe(III)呈極顯著正相關(<0.01).這3種腐殖質還原菌屬可能具有較強的電子傳遞能力,在氧化還原過程中起主導作用.有研究表明,具有將Cr(VI)還原為Cr(III)能力,該菌屬在遇到高電位的金屬離子時可能會展現出較強的還原能力[22].因此,推測當與Fe(III)在厭氧環境中共存時也較容易將Fe(III)還原為Fe(II).菌屬是分泌脲酶的主要菌屬,常用于工業廢水處理.和在FVW中與還原Fe(III)能力顯著相關(圖3),而也僅在WW中與還原Fe(III)能力顯著相關.因此,腐殖質還原菌的還原能力可能受物料環境的條件約束.在不同的物料中腐殖質還原菌存在共同還原機制,腐殖質還原菌由于復雜的生長代謝環境,不能單獨發揮還原能力,可能需要與群落中相關的菌屬相互影響來達到還原Fe(III)的作用.另外,本研究中發現,有19個菌屬在還原過程中菌種豐度逐漸降低(圖4),且該些菌種在不同物料中都呈現相同的豐度降低趨勢,在反應過程中與Fe(III)還原量呈負相關關系.這說明該些菌屬可能由于不適應復雜的微生物環境及其它微生物對其生長條件的約束,在微生物環境中逐漸失去生長優勢.

圖4 不同物料堆肥中與Fe(III)還原能力呈顯著負相關關系的腐殖質還原菌識別
*<0.05; **<0.01
2.3 腐殖質還原菌群與微環境因子的響應關系
異化鐵還原是參與天然有機污染層降解有機污染物的重要過程[23],而本研究發現堆肥過程的腐殖質還原菌具有較強的異化鐵還原能力.因此通過一定的過程調控技術手段,增加腐殖質還原菌的數量,理論上可以提升堆肥或土壤中污染物的生物修復效率,具有重要的環境意義.但由于添加外源微生物會對堆肥微生物環境造成未知影響,且腐殖質還原菌須在厭氧環境下進行篩選,對接種環境條件限制較為嚴格.基于上述理論,通過腐殖質還原菌與微環境的響應關系分析[24],不添加外源微生物的條件下構建一種促進腐殖質還原菌生長的堆肥微環境調控方法是本文主要的研究方向.
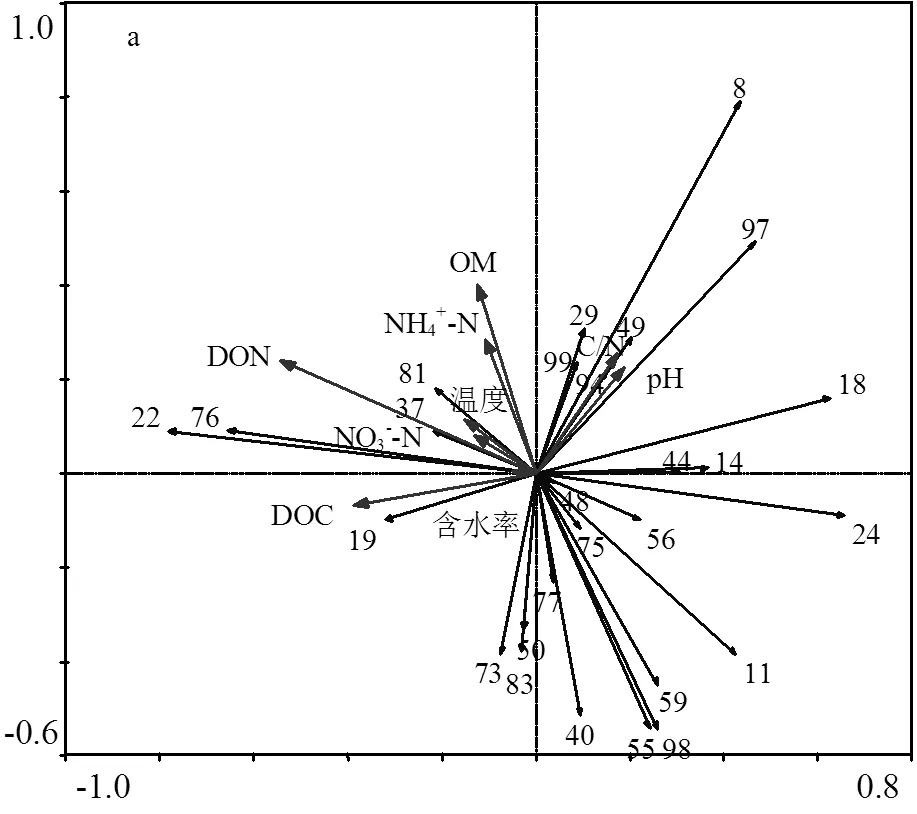
于100個與異化鐵相關菌屬中選擇普遍存在于6種物料的27個菌屬.對這些腐殖質還原菌的相對豐度采用降趨對應分析(即DCA分析),第一排序軸最大梯度為2.771.因此,選擇可以更好反應腐殖質還原菌菌群結構與微環境因子之間的響應關系的RDA來分析堆肥過程中微環境因子對腐殖質還原菌菌群結構變化的影響,以明確該27個菌屬與理化因子之間的響應關系.其中,第一排序軸與第二排序軸分別解釋了22.03%和40.64%的物種變化量.DON、DOC、pH、C/N、OM、NH4+-N、溫度、NO3--N及含水率分別單獨解釋了總變量的16.0%、8.2%、3.0%、2.8%、1.7%、1.5%、1.3%及0.6%.說明溫度、NO3--N與含水率對腐殖質還原菌的菌群分布影響較小.采用偏相關分析進一步表明,DON顯著影響腐殖質還原菌菌群結構的變化(< 0.05),說明DON是影響27屬腐殖質還原菌的重要微環境因子.然而,堆肥過程中腐殖質還原菌的新陳代謝與多種微環境因子的緊密相聯,因此,其他微環境因子對腐殖質還原菌群落也具有至關重要的影響[25].
從排序圖可進一步探究堆肥過程微環境因子對腐殖質還原菌的作用(圖5a和圖5b),其中,序號19、22、37、76及81與DON、DOC、NO3--N、SW1及SW3呈正相關關系.說明此類腐殖質還原菌的生長與氮相關指標具有密切關系,在秸稈類物料中適當添加有機碳源與氮源不僅能夠促進此類腐殖質還原菌的生長,還可能對堆肥過程中硝化作用具有一定貢獻;腐殖質還原菌序號8、29、49、94、97及99主要存在于蛋白類物料(CM、DCM)中,其中序號94與異化鐵還原能力呈極顯著正相關關系(圖3),鑒于該菌屬有較強還原金屬離子的能力,因此在蛋白質物料中主要對序號94的菌屬進行調控從而提升堆肥中重金屬污染物的修復率.從圖5a中可以看出這些腐殖質還原菌與C/N、pH呈正相關關系.說明在蛋白類物料中升高pH(£8.93)與C/N(£22.05)會在一定程度上提高此類腐殖質還原菌的豐度,這一結果也為在弱堿條件下篩選腐殖質還原菌提供可能性.序號為11、24、40、55、59、73及98的的腐殖質還原菌存在于多種類型物料中,其中與異化鐵還原能力呈極顯著正相關關系的59號菌屬在生物量分解中起主導作用[26].這些腐殖質還原菌與NH4+-N、OM和DON呈一定的負相關關系.由此可以推測,此類微生物能夠以DON與NH4+-N作為營養源進行新陳代謝.然而,若NH4+-N與DON濃度過高,也可能會抑制此類腐殖質還原菌的活性.因此還有更多的研究工作需要繼續開展,為進一步通過調控微環境促進腐殖質還原菌生長提供技術支持.
3 結論
3.1 不同物料中的腐殖質還原菌還原異化鐵能力有所不同.隨著堆肥過程進行,CM、DCM、FVW和WW中腐殖質還原菌還原能力增強.堆肥過程對GW和SW中腐殖質還原菌的還原能力無顯著影響.不同物料還原能力依次為蛋白質類物料(CM、DCM)>纖維素類物料(WW、FVW)>木質素類物料(GW、SW).
3.2 篩選了不同堆肥過程中與異化鐵還原能力極顯著相關的腐殖質還原菌,分別為Actinobacteria門的、Firmicutes門的和.具有異化鐵還原能力的腐殖質還原菌在纖維素類物料中有豐富的多樣性.
3.3 基于微環境因子與腐殖質還原菌的響應關系,篩選了堆肥過程中腐殖質還原菌的關鍵影響因素,DON能顯著影響腐殖質還原菌菌群分布.在此基礎上,構建了一種通過調節堆肥物料微環境因子,促進腐殖質還原菌生長的調控方法.
[1] Cheng D, He J Z. Isolation and characterization of "Dehalococcoides" sp. strain MB, which dechlorinates tetrachloroethene to trans-1,2- dichloroethene [J]. Applied and Environmental Microbiology, 2009, 75(18):5910-5918.
[2] Futagami T, Goto M, Furukawa K. Biochemical and genetic bases of dehalorespiration. Chemical Record, 2008,8(1):1-12.
[3] Xi B, Zhao X, He X, Huang C, et al. Successions and diversity of humic-reducing microorganisms and their association with physical- chemical parameters during composting [J]. Bioresource Technology, 2016,219:204-211.
[4] Wu C Y, Chen N, Li H, Li Q F. Kocuria rosea HN01, a newly alkaliphilic humus-reducing bacterium isolated from cassava dreg compost [J]. Journal of Soils & Sediments, 2014,14(2):423-431.
[5] Wolf M, Kappler A, Jie J, Meckenstock R U. Effects of humic substances and quinones at low concentrations on ferrihydrite reduction by Geobacter metallireducens [J]. Environmental Science & Technology, 2009,43(15):5679-5685.
[6] Klüpfel L, Piepenbrock A, Kappler A, Sander M. Humic substances as fully regenerable electron acceptors in recurrently anoxic environments [J]. Nature Geoscience, 2014,7(3):195-200.
[7] Bird L J, Bonnefoy V, Newman D K. Bioenergetic challenges of microbial iron metabolisms [J]. Trends in Microbiology, 2011,19(7): 330-340.
[8] 荊國華,李 偉,施 耀,等. Fe~(+3)(EDTA)還原菌的分離及其性能[J]. 中國環境科學, 2004,24(4):447-451.
[9] Lovley D R. Dissimilatory Fe(III) and Mn(IV) reduction [J]. Advances in Microbial Physiology, 1991,55(2):259-287.
[10] Francis A J, Dodge C J, Rose A W, Ramirez A J. Aerobic and anaerobic microbial dissolution of toxic metals from coal wastes: mechanism of action [J]. Environmental Science & Technology, 1989, 23(4):435-441.
[11] Weber K, Achenbach L, Coates J. Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction [J]. Nature Reviews Microbiology, 2006,4(10):752-764.
[12] Martinez C M, Alvarez L H, Celis L B, Cervantes F J. Humus- reducing microorganisms and their valuable contribution in environmental processes [J]. Applied Microbiology & Biotechnology, 2013,97(24):10293-10308.
[13] Lovley D R, Nevin K P. Lack of production of electron-shuttling compounds or solubilization of Fe(III) during reduction of insoluble Fe(III) oxide of Geobacter Metallireducens [J]. Applied and Environmental Microbiology, 2000,66(5):2248-2251.
[14] Jiang J, Kappler A. Kinetics of microbial and chemical reduction of humic substances: implications for electron shuttling [J]. Environmental Science & Technology, 2008,42(10):3563-3569.
[15] Lovley D R, Coates J D, Bluntharris E L, Phillips E J, et al. Humic substances as electron acceptors for microbial respiration [J]. Nature, 1996,382(6590):445-448.
[16] Zhao X, He X, Xi B, Gao R, et al. Response of humic-reducing microorganisms to the redox properties of humic substance during composting [J]. Waste Management, 2017,70:37-44.
[17] 汪明霞,王 娟,司友斌. Shewanella oneidensis MR-1異化還原Fe(Ⅲ)介導的As(Ⅲ)氧化轉化[J]. 中國環境科學, 2014,34(9):2368-2373.
[18] Roelcke M, Han Y, Cai Z, Richter J. Nitrogen mineralization in paddy soils of the Chinese Taihu Region under aerobic conditions [J]. Nutrient Cycling in Agroecosystems, 2002,63(2/3):255-266.
[19] Ait B G, Cegarra J, Merlina G, Revel J C, et al. Qualitative and quantitative evolution of polyphenolic compounds during composting of an olive-mill waste-wheat straw mixture [J]. Journal of Hazardous Materials, 2009,165(1-3):1119-1123.
[20] Said-Pullicino D, Erriquens F G, Gigliotti G. Changes in the chemical characteristics of water-extractable organic matter during composting and their influence on compost stability and maturity [J]. Bioresource Technology, 2007,98(9):1822-1831.
[21] Zhang J, Zeng G, Chen Y, Yu M, et al. Effects of physico-chemical parameters on the bacterial and fungal communities during agricultural waste composting [J]. Bioresource Technology, 2011,102(3):2950-2956.
[22] Zhu W, Yang Z, Ma Z, Chai L. Reduction of high concentrations of chromate bysp. CRB1isolated from Changsha, China [J]. World Journal of Microbiology & Biotechnology, 2008,24(7):991-996.
[23] Lovley D R, Woodward J C. Mechanisms for chelator stimulation of microbial Fe(III)-oxide reduction [J]. Chemical Geology, 1996, 132(1-4):19-24.
[24] Wei Y, Zhao Y, Fan Y, Lu Q, et al. Impact of phosphate-solubilizing bacteria inoculation methods on phosphorus transformation and long-term utilization in composting [J]. Bioresource Technology, 2017, 241:134-141.
[25] Wang X, Cui H, Shi J, Zhao X, et al. Relationship between bacterial diversity and environmental parameters during composting of different raw materials [J]. Bioresource Technology, 2015,198:395-402.
[26] Zhao D, Cao X, Huang R, Zeng J, et al. Variation of bacterial communities in water and sediments during the decomposition of Microcystis biomass [J]. Plos One, 2017,12(4):e0176397.
A regulating method for humic-reducing microorganisms and assessment of the reduction of dissimilatory Fe(III)in composting.
ZHAO Xin-yu1,2, FAN Yu-ying1,2,3, XI Bei-dou1,2*, TAN Wen-bing1,2, HE Xiao-song1,2, ZHANG-Hui1,2, LI-Dan1,2
(1.State Key Laboratory of Environmental Criteria and Risk Assessment, Chinese Research Academy of Environmental Sciences, Beijing 100012, China;2.State Environmental Protection Key Laboratory of Simulation and Control of Groudwater Pollution, Chinese Research Academy of Environmental Sciences, Beijing 100012, China;3.Northeast Agricultural University, Harbin 150030, China)., 2018,38(10):3815~3822
Reduction of dissimilatory Fe(III) by humic-reducing microorganisms (HRMs) from different composts was conducted. Results showed that the capacity for reduction of dissimilatory Fe(III) by HRMs was ranked in the order protein-rich compost > cellulose-rich compost > lignin-rich compost. The result of correlation showed that、andwere significantly associated with the Fe(III) reduction. It was indicated that dissolved organic nitrogen was the primary micro-enviromental factors significantly driving the variation of these key HRMs by Redundancy analyses (RDA). Finally, based on the relationship between the mico-enviromental factors and HRMs, this work proposed a regulating method to enhance the growth of the key HRMs during composting. This study not only investigated the influencial factors of HRMs in compost deeply, but also had ecologicalsignificant in the study of waste biogeochemical process.
humic-reducing microorganism;16S rDNA;reduction of dissimilatoryFe(III);RDA;compost
X705
A
1000-6923(2018)10-3815-08
趙昕宇(1989-),女,黑龍江雙鴨山人,中國環境科學研究院博士后,主要從事有機固體廢棄物處置與資源化研究.發表論文11篇.
2018-06-25
國家自然科學基金資助項目(41501242)
* 責任作者, 研究員, xibeidou@yeah.net
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
