塔里木盆地5個生態小區稀有放線菌分離及合成抗生素基因分布
2018-10-26 02:12:32朱榮貴關統偉姜秀娟
生物技術通報 2018年9期
關鍵詞:檢測
朱榮貴 關統偉 姜秀娟
(1. 四川理工學院生物工程學院,自貢 643000;2. 西華大學食品與生物工程學院,成都 610039)
隨著人類社會的發展,新型人類疾病不斷出現、病原微生物對臨床藥物耐藥性不斷增加以及動植物病害嚴重發生等問題日益嚴重,促使人們呼喚新結構新活性抗生素的發現,目前,新抗生素中50%-78%由放線菌產生[1-2]。稀有放線菌是一類出現頻率比鏈霉菌低得多的放線菌類群,有著產生新的天然活性物質的極大潛力[3-4],其研究一直備受關注[5]。
塔里木盆地是我國最大內陸盆地,氣候極端干旱,植被稀少,土壤荒漠化和鹽堿化嚴重,環境較為特殊,可能存在獨特的稀有放線菌[6-7]。微生物的數量和組成與其生存環境(水、熱、pH、植被、土壤等生態條件)密切相關[8-9],為了給研究者有目標的分離獲得該地區稀有放線菌資源提供參考,本研究根據倪健等[10]對塔里木盆地生態小區的劃分,選擇5個代表性生態小區,使用選擇性培養基分離稀有放線菌,分析不同生態小區稀有放線菌分布情況。
為提高產活性物質潛力菌的篩選效率,本項目組還對抗生素合成相關基因進行了篩選,主要包括I型PKS、II型PKS、NRPS、APH和HMG-CoA。它們是相應抗生素類代謝產物產生的關鍵催化物,其中聚酮合酶(Polyketide synthase,PKS)能夠催化小分子羧酸類物質合成具有重要藥用價值的次級代謝產物——聚酮類化合物。放線菌主要含有I型PKS和II型PKS,I型PKS催化合成的抗生素主要包括大環內脂類、聚醚類、多烯類和安莎類;II型PKS催化合成的抗生素主要包括蒽環類和四環類。非核糖體多肽合成酶(Nonribosomal Peptide -synthetase,NRPS)是一些重要多肽類抗生素的合成系統中起關鍵作用的酶,能識別特定的氨基酸并將其直接相連形成多肽鏈。氨基糖苷磷酸轉移酶(Aminoglycoside Phosphotransferases,APH)與氨基糖苷類抗生素合成密切相關[11],氨基糖苷類抗生素主要包括鏈霉素、新霉素等。3-羥基-3-甲基戊二酰輔酶A(3-Hydroxy-3-Methylglutaryl Coenzyme A,HMG-CoA)還原酶與類異戊二烯的生物合成密切相關,后者有極大的臨床應用價值[12]。這些基因是篩選生產相應抗生素放線菌的重要依據。本研究根據基因篩選評估稀有放線菌的抗生素類活性物質的產生潛力,旨在為進一步開發利用稀有放線菌提供菌種基礎。
1 材料與方法
1.1 材料
試驗選擇塔里木盆地具代表性的5個生態小區,根據各小區的面積,每小區選擇2-4個點進行采樣,采樣取土下5-20 cm深度的土壤,裝入滅菌袋,編號注明采集信息,4℃低溫保藏帶回實驗室后,將各小區樣品以四分法均勻混合并檢測理化性質,以5份混合土樣為材料進行分離。各樣品理化性質見表1。
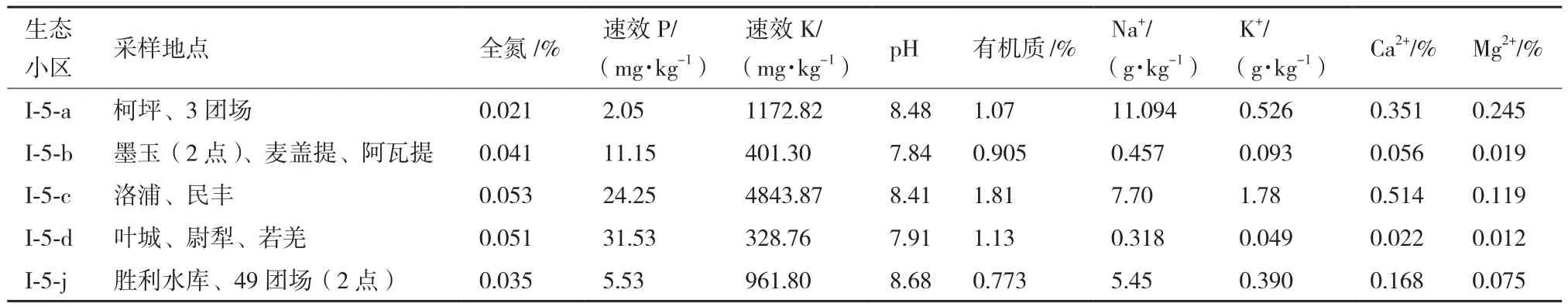
表1 塔里木盆地5個生態小區混合土樣理化特性
1.2 方法
1.2.1 稀有放線菌分離 (1)預處理。干熱處理:土樣風干,研磨過篩,120℃干熱處理1 h,稱取1g樣品到10 mL的無菌水中,120 r/min振蕩10 min。苯酚處理:取1 g樣品加入到1.5% 苯酚的無菌水中,30℃水浴30 min。SDS處理:稱取1 g土樣,用0.05%的SDS(5 mmol/l磷酸鹽緩沖液配制),加入6%的酵母膏,40℃下 200 r/min 振蕩 20 min[13]。
(2)分離培養:采用稀釋法涂板分離,使用改良HV培養基;ATCC172培養基[14];海藻糖-脯氨酸培養基;改良脯氨酸培養基;改良高氏二號培養基[15];葡萄糖-天門冬酰氨培養基[16]和YIM-3培養基[17]等7種培養基進行復合篩選。培養基加入終濃度為60 μg/mL的重鉻酸鉀、50 μg/mL的放線菌酮和25 μg/mL的制霉菌素來抑制細菌和霉菌的生長。以預處理和培養基為因素,設置21個處理進行分離,28℃培養 2 周[18]。
(3)純化:根據菌落形態挑菌純化,純化培養基為ISP4培養基。ISP 4培養基:淀粉10 g,(NH4)2SO44g,CaCO32 g,MgSO42 g,K2HPO42 g,瓊脂16-18 g,水1 000 mL,pH調至7.0-7.5。
1.2.2 菌種鑒定 采用16S rRNA基因序列分析方法鑒定菌種。以酶小量法提取待測菌株基因組DNA[16],利用通用引物(正向 PA :5′-CAGA GTTTGATCCTGGCT-3′和反向 PB :5′-AGGAGGTGA TCCAGCCGCA -3′)進行16S rDNA基因的擴增,PCR擴增產物送至上海生工生物工程技術服務有限公司進行序列測定。獲得的16S rRNA基因序列在NCBI上采用Blast Search法進行相似性檢索,從GenBank和EMBL/DDBJ數據庫中搜集到相關放線菌的16S rRNA基因序列進行比較鑒定。最后使用顯微鏡觀察形態特征并與分子鑒定結果比照進行綜合鑒定。
1.2.3 生態分布及多樣性分析 分析各生態小區菌種分布及多樣性,計算不同小區的物種均勻度(Evenness)、豐富度(Shannon)和多樣性(Simpson)指數,其中Simpson指數用如下公式計算:

其中,N為個體總數,S屬數,Ni為i屬的個體數。
1.2.4 抗生素合成相關基因擴增 采用5對引物擴增菌株的功能基因。引物K1F(5′-TSAAGTCSAACA TCGGBCA-3′) 和 M6R(5′-CGCAGGTTSCSGTAC CAGTA-3′)用來擴增 PKS-I基因[19],引物 KSa(5′-TSGRCTACRTCAACGGSCAC GG-3′)和 KSβ(5′-TACSAGTCSWTCGCCTG GTTC-3′)用來擴增 PKS-II基因[20],引物 A3F(5′-GCSTACSYS ATSTACACSTCSGG-3′) 和 A7R(5′-SASGTCVCCSGTSCGG TAS-3′)用來擴增NRPS基因,引物STR-F(5′-CGGCTGCTCGACCACGAC-3′) 和 STR-R(5′-GTCCTCGATGTCCCACAG-3′)用來擴增 APH 基因[21],引物 HMGF(5′-GGGCATCGCCGCGACCCTCGTCGACGAGCG-3′)和 HMGR(5′-GCGATGACGGCG AGGC GGCGGGCGTTCTC-3′)用來擴增 HMG-CoA 基因[12]。擴增產物進行電泳檢測,目的條帶進行割膠回收純化,克隆測序,通過Blast比對進行驗證。
1.2.5 拮抗活性初篩 采用抑菌圈法檢測分離到的放線菌對金黃色葡萄球菌(Staphylococcus aureus,菌種編號ATCC 25923)、大腸桿菌(Escherichia coli,菌種編號ATCC 25922)的拮抗活性(本部分實驗病原菌均購于中國農業菌種保存中心)。使用血球計數板將病原菌調制成108個/ mL濃度的菌懸液,將金黃色葡萄球菌、大腸桿菌菌懸液取100 μL涂布到LB培養基上,再在平板上接種放線菌的菌餅(瓊脂塊直徑0.6 cm)。LB培養平板37℃培養24-48 h。采用平板對峙培養法檢測分離菌對給新疆主要經濟作物造成危害的棉花枯萎病原菌(Fusarium oxysperiumsp.Vasinfectum,菌種編號ACCC 31038)、棉花黃萎病原菌(Verticillium dahliae,菌種編號ACCC 36211)和辣椒疫霉病原菌(Phytophthoracapsici,菌種編號ACCC 36278)的拮抗活性,將棉花枯、黃萎病原菌和辣椒疫霉病原菌菌懸液取100 μL涂布到PDA培養基上,接種放線菌(懸液濃度及方法同上)PDA培養平板28℃培養5 d。測量以上兩組實驗的抑菌圈直徑(病原菌的菌落/絲與放線菌之間形成的隔離帶外圈的直徑),每個菌種做3組重復,取3組重復平均值,判斷放線菌對病原菌的拮抗活性。
2 結果
2.1 生態小區稀有放線菌分布分析
2.1.1 生態小區稀有放線菌組成狀況 對分離到的放線菌菌株先觀察其菌落形態特征,選擇形態差異明顯的稀有放線菌菌株,并在ISP 4培養基上插片培養1周,取片進行顯微觀測,記錄其菌體、基絲形態,比照《放線菌的分類和鑒定》(閻遜初,1992),并結合16S rRNA基因序列在Blast上的比對結果進行菌屬鑒定。結果(表2)顯示分離得18株稀有放線菌,屬于放線菌的10個屬。
不同生態小區稀有放線菌分布種類由多到少依次為:塔里木河河岸林生態小區(I-5-j)10株,分別屬于8屬;塔克拉瑪干沙漠和田河西部生態小區(I-5-b)9株,分別屬于8屬;塔里木北部壤質荒漠生態小區(I-5-a)8株,分別屬于8個屬;塔克拉瑪干沙漠中部生態小區(I-5-c)7株,分別屬于7屬;塔克拉瑪干沙漠塔里木河東部生態小區(I-5-d)4株,分別屬于4個屬。
從稀有放線菌在生態小區的分布情況看,三項指數豐富度、均勻度、多樣性均為壤質土壤小區較高(表3),反映出壤質土壤較為適宜微生物生長,其種類也很豐富。此外,從總體上看,不同植被土樣放線菌分布,以耕作土多樣性較為豐富,這一點與其它研究者結論基本相符,原因可能為耕作土水分充足、施肥較多、熟化程度高,利于稀有放線菌繁衍。
2.1.2 多樣性分析 依據每個生態小區稀有放線菌組成、菌數(CFU計,數據未列出),計算不同小區的物種均勻度(Evenness)、豐富度(Shannon)和多樣性(Simpson)指數。結果(表3)顯示豐富度指數以塔里木北部壤質荒漠生態小區最大,為2.382 1,該小區物種所涵蓋屬的數目最多(屬數相同情況下,各屬的個體數目分配均勻性最高小區多樣性指數最高);物種均勻度指數以塔克拉瑪干沙漠塔里木河東部生態小區最大,為0.871 8,說明該小區各個菌種的個體數目分配均勻性最高;Simpson指數是綜合均勻度和豐富度的一項多樣性指數,以塔里木北部壤質荒漠生態小區最大,為0.779 9,該小區的稀有放線菌屬數多且各屬的種數較為均勻,多樣性最好。
2.2 抗生素合成相關基因及菌株的拮抗效果檢測
利用5對引物通過PCR擴增菌株抗生素合成相關基因,測得序列通過Blast比對進行驗證。驗證序列與相關功能基因相符為“陽性(+)”,不符為“陰性(-)”。對分離菌對病原菌的對峙培養平板進行檢測,抑菌圈直徑<0.2 cm為無拮抗性(陰性“-”),0.2 cm≤抑菌圈直徑≤0.4 cm為弱拮抗性(陽性“+”),抑菌圈直徑>0.4 cm為強拮抗性(++)、對分離菌的5種功能基因的分布及拮抗檢測結果見表4。
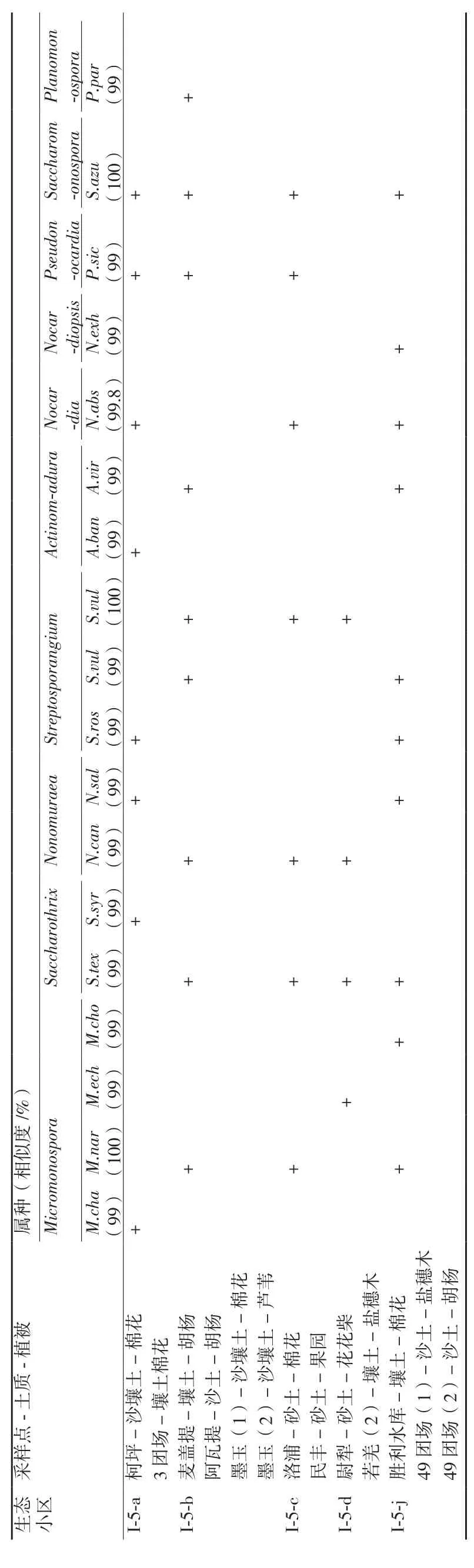
表2 不同生態小區土樣放線菌組成
18株稀有放線菌中,有14株菌含有1種以上的功能基因,其中的9株菌含有I型PKS基因,4株菌含有II型PKS基因,3株菌含有NRPS基因,4株菌含有APH基因,沒有檢測到HMG-COA基因。有一株鏈孢囊菌含有4種功能基因,有合成多種抗生素的潛力。I型PKS在稀有放線菌中的分布較為普遍,在這18株菌能占到有50%的比例,有產生大環內酯類和聚醚類等抗生素的潛力。HMG-COA基因在這18株菌中沒有檢測到,可能不具有產生異戊二烯化合物的潛力,通過檢測,可以為有針對性的篩選產次級代謝產物稀有放線菌提供指導。
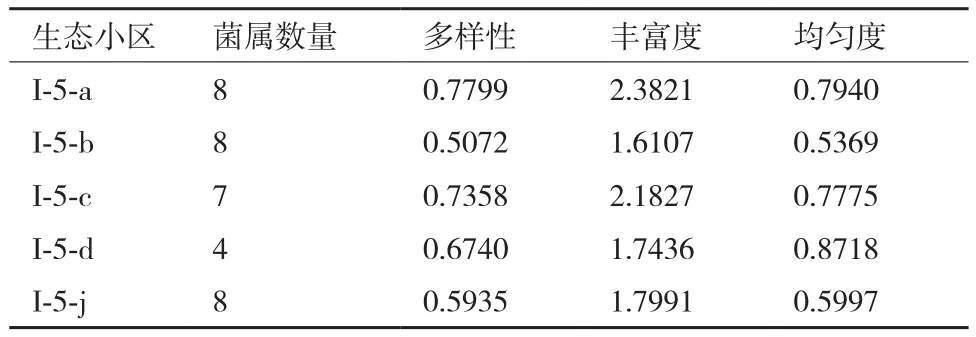
表3 不同生態小區物種的多樣性
觀察放線菌的拮抗效果[22]。菌株47017、47029、47035和47037對金黃色葡萄球菌有強的拮抗活性,菌株47004和47011對金黃色葡萄球菌有弱拮抗活性。18株菌對大腸桿菌沒有拮抗活性。菌株47017對棉花黃萎病原菌有拮抗活性,菌株47029對棉花枯萎病原菌和棉花黃萎病原菌都有拮抗活性,其它菌株沒有拮抗3種植物病原真菌的活性。以含APH基因的47029菌株拮抗種類最多,表現出了廣譜抗菌活性。有4株含有PKSⅠ型基因的菌對金黃色葡萄球菌表現出拮抗性,菌株所含基因種類與其對病原菌拮抗性效果見表4。
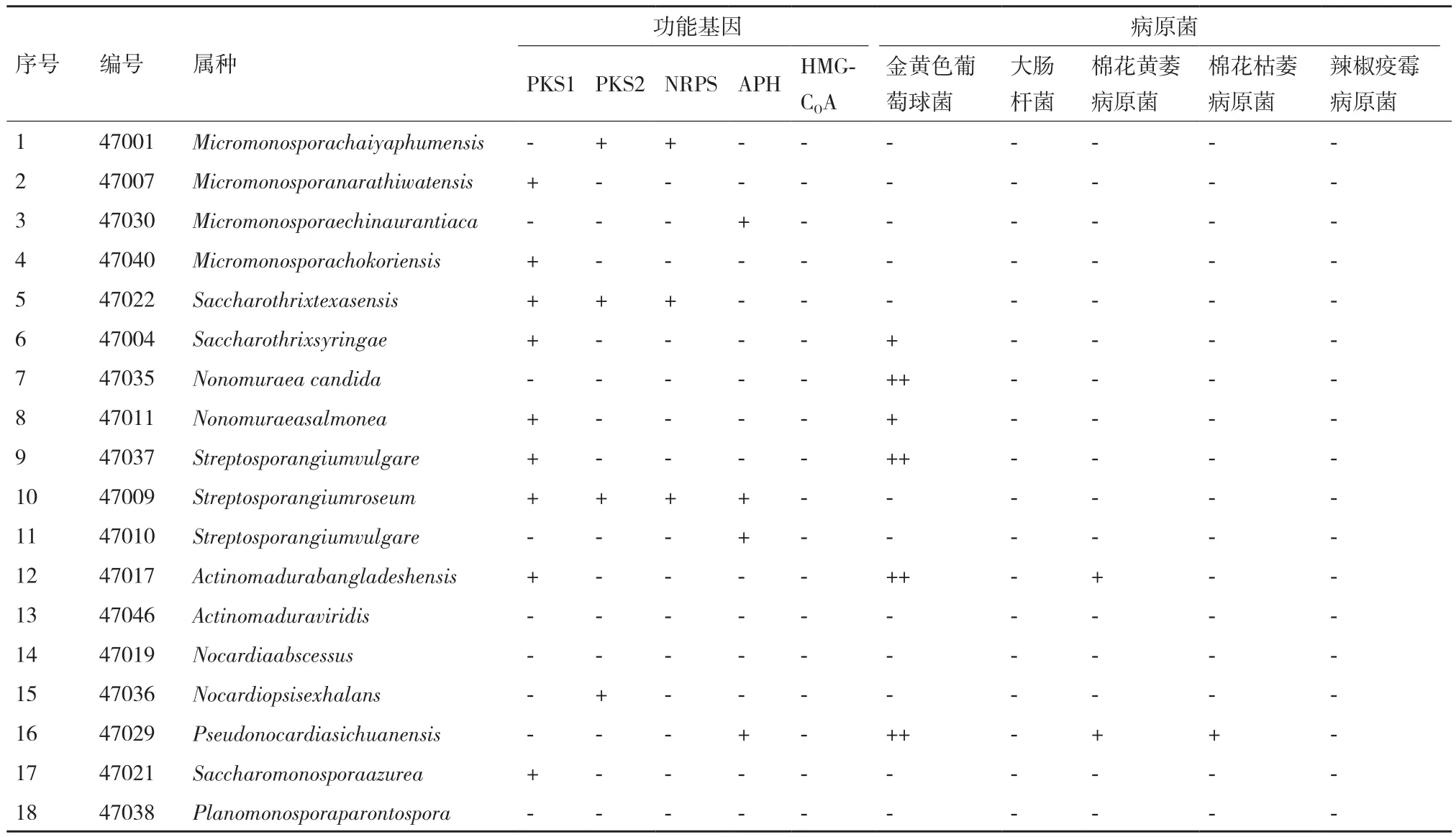
表4 五種功能基因在18株稀有放線菌中的分布及菌株的拮抗效果檢測
3 討論
從分離結果來看,幾個樣點的稀有放線菌種類較多。一些產生物活性物質的重要菌源,如小單孢菌、馬杜拉放線菌、諾卡氏菌等都能分離到。但從分離的角度講,由于本研究采樣點集中于塔里木河沿線地區及塔中地區,涉及生態區域僅占整個盆地1/3左右(共計13個生態小區)。加之實驗雖選用了一些分離效率比較高的培養基,但實際上很多稀有放線菌培養條件特殊[23]。因此,本實驗可能無法分離到一些特殊的微生物。研究者仍需要不斷拓展分離源和創新分離手段來開發這些資源。
本次分離菌株含有較為豐富PKS、NRPS及APH基因,基因分布以PKS I型最多,未檢出HMGCoA還原酶基因。PKS I型基因陽性率為50%,II型PKS為22.2%,NRPS為16.7%,APH為22.2%。除PKS I型基因外,其它基因檢出率與已報道鏈霉菌的檢出率相當[24]。有的菌株同時含有多種功能基因(如鏈孢囊菌),因而可能產生不同的抗菌或細胞毒活性物質,從這一層面講,本研究所分離的放線菌菌群具有更為復雜的多樣性。此外,兩株親緣關系極近的菌株47010、47037,含有不同的抗生素合成基因,這也說明,菌株的產活性物質能力具有菌株特異性而非菌種特異性,通過分子檢測的手段可以忽略菌株之間的相似性而快速的分析菌株的產抗潛力。因此,分子檢測不僅能檢驗菌株的產抗潛力,也是同一物種不同菌株間進行細致分類的研究方法之一。
本研究基因和活性之間的關系不能一一對應,如菌株47035雖然沒有抗生素合成基因,但具有拮抗金黃色葡萄球菌的活性。可能原因:一是,這些菌株產生其他種類的抗生素,而我們檢測的基因有限,出現“漏”篩。二是,PKS、NRPS等基因本身存在多樣性,同一種基因的表達也可能在結構和活性上有差別。
此外,18株放線菌中有14株具有抗生素合成相關基因,但只有6株具有拮抗病原菌的活性,其可能原因:一是所選取靶標種類較少,放線菌產生的抗生素對上述5種靶標菌不具有拮抗性(但可能對其它靶標菌具有作用),需要增加靶標菌檢測;二是野生型的菌株可能只產生痕量的次級代謝產物,或者合成基因沉默表達,從而對靶標菌不能表現出拮抗性;三是檢測培養基不合適。對于這些菌株,筆者認為不能簡單定義為“無用菌株”。因為它們往往含有次級代謝產物的合成基因簇。通過激活其次級代謝產物合成基因的表達,如摸索培養、發酵條件,找出抗生素合成基因表達的調控因素,或者通過基因操作如核糖體工程[25]等手段,激活潛在“有用”次級代謝產物相關基因表達,甚至通過誘變,使菌株突變產生新的化合物,都可能使其功能基因發揮作用。這也是分子手段篩選產抗生素菌的一項優勢所在,不僅提高了篩選效率,也能充分挖掘潛在的有用菌源。
4 結論
本實驗從塔里木盆地5個生態小區土樣中分離稀有放線菌,經16S rRNA序列及形態分析,鑒定分離得18種菌株,其中塔里木北部壤質荒漠生態小區菌種豐富度指數最大,為2.3821。通過擴增分離菌株產抗生素相關基因I型PKS、II型PKS、NRPS、APH及HMG-CoA,并進行電泳分析,有14株菌含有抗生素合成相關基因,其中4株含有多種基因。檢測分離菌株對5種主要動植物病原菌的抗菌活性,有6株具有拮抗活性。其中2株具有廣譜抗菌活性。本研究基因和活性之間關系并非一一對應。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48