鎘對梭魚草葉片保護酶活性、光合及熒光特性的影響
2018-10-19 05:23:34辛建攀李文明祁茜田如男
草業(yè)學(xué)報 2018年10期
關(guān)鍵詞:植物
辛建攀,李文明,祁茜,田如男
(南京林業(yè)大學(xué)風(fēng)景園林學(xué)院,江蘇 南京 210037)
鎘主要來源于農(nóng)藥使用、污水排放、采礦及工業(yè)生產(chǎn)等人類活動,是環(huán)境中生物毒性最強的重金屬元素之一,具有較強的移動性、化學(xué)活性和較長的生物半衰期,經(jīng)植物根系吸收后可迅速轉(zhuǎn)移至地上部分[1-2]。研究表明,植物能夠通過提高超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)等保護酶活性來忍耐一定程度的鎘脅迫[3-5]。但是,較高濃度的鎘會破壞植物的多種代謝活動,使細胞內(nèi)超氧陰離子自由基(O2-·)、過氧化氫(H2O2)等活性氧自由基大量積累,引起膜脂過氧化作用,抑制酶活性或破壞葉綠體結(jié)構(gòu),不利于植物光合作用及氣體交換等生理活動,從而抑制植物生長[6-7]。
梭魚草(Pontederiacordata)隸屬于雨久花科(Pontederiaceae)梭魚草屬(Pontederia),原產(chǎn)于新大陸美洲熱帶及溫帶地區(qū),主要分布于沼澤、濕地、水渠、湖泊、池塘及河流邊緣淺水區(qū)域等,為專性濕地植物,具有很高的園林觀賞和生態(tài)應(yīng)用價值,在景觀水體綠化美化、濕地恢復(fù)及重建過程中備受青睞[8-11]。從已有資料來看,有關(guān)梭魚草的研究多集中于繁殖生物學(xué)[12-13]、富營養(yǎng)化水體凈化[14-15]等方面,也有學(xué)者對梭魚草去除重金屬鎘的效率進行了比較分析[16],但鎘脅迫對梭魚草生理生化特性的影響尚未有相關(guān)報道。因此,本研究通過溶液培養(yǎng)法,初步分析了不同濃度鎘脅迫下梭魚草葉片保護酶活性、葉綠素含量、氣體交換參數(shù)及葉綠素?zé)晒馓匦缘淖兓瑸榱私馑篝~草的耐鎘能力和生理機制提供理論基礎(chǔ)。
1 材料與方法
1.1 供試材料
試驗于2017年4-7月在南京林業(yè)大學(xué)風(fēng)景園林學(xué)院園林教學(xué)中心溫室完成,供試材料為梭魚草。選取長勢一致、健康植株,將根系沖洗干凈后移至于盛有2 L 1/2 Hoagland營養(yǎng)液的塑料桶中進行適應(yīng)性培養(yǎng)。每桶1株,培養(yǎng)周期為15 d,期間實時補充營養(yǎng)液至初始體積。
1.2 鎘處理
適應(yīng)性培養(yǎng)結(jié)束后,對供試材料進行鎘(Cd)脅迫處理。鎘溶液由CdCl2·2.5H2O和1/2 Hoagland營養(yǎng)液配制而成,共設(shè)置4個濃度梯度,分別為0、5、25和75 mg·L-1(以Cd2+濃度計算)(分別記為CK、A1、A2和A3)。各處理重復(fù)3次,每個重復(fù)8株。分別于第0,7,17,25天時測定葉片保護酶活性、葉綠素含量、光合氣體交換參數(shù)及葉綠素?zé)晒鈪?shù)等生理生化指標。其中,光合氣體交換參數(shù)和葉綠素?zé)晒鈪?shù)選擇同一葉片的中部(避開主脈)進行測定。每次取樣和相關(guān)指標測定結(jié)束后,及時更換營養(yǎng)液并添加對應(yīng)濃度的鎘溶液。
1.3 測定指標與方法
葉綠素含量測定采用丙酮乙醇混合液法[17];相對電導(dǎo)率測定采用電導(dǎo)法[18];SOD、POD及CAT活性測定分別采用氮藍四唑(NBT)光還原法[19]、愈創(chuàng)木酚法和紫外吸收法[18]。
光合氣體交換參數(shù)測定:于晴朗無風(fēng)的天氣,使用Ciras-2型便攜式光合測定系統(tǒng)于上午9:00-11:30測定各處理下植株上部功能葉光合氣體交換參數(shù)。測定過程中,使用開放氣路,葉室溫度設(shè)置為25 ℃,光強設(shè)置為1000 μmol·m-2·s-1),CO2濃度設(shè)置為390 μmol·mol-1,相對濕度為60%。測定指標包括凈光合速率(Pn)、蒸騰速率(E)、氣孔導(dǎo)度(Gs)以及胞間CO2濃度(Ci)等。
葉綠素?zé)晒鈪?shù)測定:于上午9:30-11:30用Handy PEA植物效率儀測定系統(tǒng)對各處理下梭魚草葉片的葉綠素?zé)晒鈪?shù)進行測定。用暗適應(yīng)夾夾住葉片中部,使葉片充分暗適應(yīng)30 min。暗適應(yīng)后將分析探頭置于葉夾上,打開葉夾遮光片,使葉片暴露在飽和脈沖光(3000 mmol·m-2·s-1)下1 s。初始熒光(Fo)、最大熒光(Fm)、相對可變熒光(Vj、Vi)及PSⅡ最大光化學(xué)效率(Fv/Fm)等葉綠素?zé)晒鈪?shù)從Handy PEA軟件測定結(jié)果中直接導(dǎo)出。
1.4 數(shù)據(jù)分析
采用SPSS 19.0進行數(shù)據(jù)處理和分析,對不同處理之間進行單因素方差分析和Duncan多重比較(P<0.05),用WPS表格作圖。
2 結(jié)果與分析
2.1 鎘脅迫對梭魚草葉片細胞膜透性的影響

圖1 鎘脅迫對梭魚草葉片相對電導(dǎo)率的影響Fig.1 Effects of Cd on the relative electric conductivities in P. cordata’s leaves 數(shù)據(jù)為平均值±平均誤差(n=3),同一時期不同處理之間小寫字母表示0.05水平顯著差異性(P<0.05)。下同。Data are mean±SD (n=3). Different small letters among treatments in the same period indicate significant differences at the P<0.05 level. The same below.
植物葉片細胞膜透性的大小可以用相對電導(dǎo)率來衡量。與對照組相比,不同濃度鎘脅迫下葉片相對電導(dǎo)率均呈上升趨勢,且隨著鎘濃度的增加,葉片相對電導(dǎo)率的上升幅度增大(圖1)。5 mg·L-1鎘脅迫下葉片相對電導(dǎo)率在第25天時顯著上升,比對照增加了29.92%;25 mg·L-1鎘脅迫下葉片相對電導(dǎo)率在第17天時比對照顯著增加了27.27%;75 mg·L-1鎘脅迫下葉片相對電導(dǎo)率在第7天比同期對照顯著增加了39.69%,脅迫結(jié)束時植株已全部死亡,表明低濃度鎘在短時間內(nèi)沒有對植株造成傷害,但隨鎘濃度的增大和處理時間的延遲,植株受害程度加劇,膜脂過氧化作用增強,細胞膜透性逐漸增大,胞內(nèi)電解質(zhì)大量外滲,導(dǎo)致葉片枯黃、脫落甚至植株死亡。
2.2 鎘脅迫對梭魚草葉片保護酶活性的影響
2.2.1SOD活性 不同濃度鎘處理下梭魚草葉片SOD活性的變化如圖2A所示。與對照組相比,當(dāng)鎘濃度為5 mg·L-1時,梭魚草葉片SOD酶活性無顯著變化。當(dāng)鎘濃度為25 mg·L-1時,葉片SOD活性先下降,在第7天時降至最低(157.90 U·g-1),這可能是梭魚草對鎘脅迫的應(yīng)激性反應(yīng);隨著脅迫時間的延長,SOD活性逐漸上升至對照水平(200.81 U·g-1),這是梭魚草對鎘脅迫的適應(yīng)性反應(yīng),即梭魚草葉片SOD清除O2-·的能力增強,有利于保護梭魚草葉片膜結(jié)構(gòu)的完整性。當(dāng)鎘濃度為75 mg·L-1時,SOD活性隨脅迫時間的推移呈下降趨勢,25 d時葉緣或葉尖變枯黃甚至葉片凋落,表明高濃度鎘降低了SOD清除能力,使細胞內(nèi)O2-·大量積累,對細胞產(chǎn)生毒害作用,從而加劇植株受害程度。
2.2.2POD活性 與對照組相比,不同濃度鎘處理下梭魚草葉片POD活性呈先上升后下降趨勢(圖2B)。當(dāng)鎘濃度為5、25 mg·L-1時,葉片POD活性在第17天時分別上升至峰值(135.28,149.42 U·g-1),比對照組增加了127.73%、151.71%,其中5 mg·L-1鎘處理下的葉片POD活性在25 d時維持在相對穩(wěn)定水平,有利于清除過氧化氫(H2O2),而25 mg·L-1鎘處理下的葉片POD活性在25 d時急劇下降至對照水平,表明梭魚草能夠通過提高葉片POD活性來減緩鎘對其造成的傷害,但這種調(diào)節(jié)作用具有一定的局限性。當(dāng)鎘濃度為75 mg·L-1時,葉片POD活性在第17天時上升至118.91 U·g-1,比對照組增加了98.46%,但此時仍然出現(xiàn)了葉緣或葉尖枯黃、葉片脫落甚至植株死亡的現(xiàn)象,這可能是高濃度鎘減緩了葉片POD活性的增加,導(dǎo)致胞內(nèi)H2O2的積累大于其清除而引起的。
2.2.3CAT活性 不同濃度鎘處理對梭魚草葉片CAT活性的影響如圖2C所示。5、25 mg·L-1鎘脅迫下葉片CAT活性基本上處于相對穩(wěn)定狀態(tài),表明胞內(nèi)H2O2

圖2 鎘脅迫對梭魚草葉片保護酶活性的影響Fig.2 Effects of Cd on antioxidant enzyme activities in P. cordata’s leaves
等活性氧自由基的產(chǎn)生與清除處于動態(tài)平衡狀態(tài),其中25 mg·L-1鎘脅迫下葉片CAT活性僅在第25天時比對照組顯著下降了56.27%,表明梭魚草對鎘脅迫具有一定的適應(yīng)性。75 mg·L-1鎘脅迫下葉片CAT活性在第17天上升至24.74 U·g-1,但與對照組差異不顯著;第25天葉片已枯黃甚至凋落,表明隨鎘濃度的增加和脅迫時間的延長,CAT清除能力下降,細胞內(nèi)H2O2等活性氧積累過多,從而加速了葉片衰老。
2.3 鎘脅迫對梭魚草葉片光合色素含量的影響
不同濃度鎘脅迫下梭魚草葉片Chla、Chlb、ChlT含量及Chla/b比值的變化如表1所示。處理7 d時,各處理下葉片Chlb、ChlT含量及Chla/b比值與對照組均無顯著差異,而Chla含量則顯著下降了7.93%、6.33%和16.63%,表明Chla對重金屬鎘脅迫比較敏感。處理17 d時,葉片Chla含量顯著低于對照組;Chlb含量隨鎘濃度的升高呈下降趨勢,其中75 mg·L-1脅迫下Chlb含量比對照顯著下降了38.36%;Chla/b比值呈下降趨勢,但與對照組未達到顯著水平;ChlT含量隨鎘濃度的升高呈下降趨勢,其中25、75 mg·L-1鎘脅迫下ChlT含量比對照組顯著下降了17.67%、45.17%。處理25 d時,各處理下葉片Chla、Chlb、ChlT含量及Chla/b比值均低于對照組,且差異達到顯著水平。總體上來看,梭魚草葉片各色素含量隨鎘濃度的升高呈下降趨勢,這可能是因為鎘在梭魚草葉綠體中積累,破壞了葉綠體結(jié)構(gòu),使葉綠素酶活性失調(diào),導(dǎo)致葉綠素的合成速率下降或分解速率加快。
2.4 鎘脅迫對梭魚草葉片光合氣體交換參數(shù)的影響
2.4.1凈光合速率(Pn) 試驗期間,鎘處理均在不同程度上抑制了梭魚草葉片Pn(圖3A)。與對照組相比,鎘脅迫下梭魚草葉片Pn隨鎘濃度的增加呈下降趨勢,且下降幅度隨脅迫時間的延長而增加。5、25 mg·L-1鎘脅迫下葉片Pn在第25天時比同期對照組分別顯著下降了26.20%、54.80%。75 mg·L-1鎘脅迫下葉片Pn在第7,17天比同期對照組分別下降了 55.42%、67.70%。
2.4.2蒸騰速率(E) 鎘脅迫下梭魚草葉片E隨鎘濃度的增加呈下降趨勢(圖3B)。其中,5 mg·L-1鎘脅迫下葉片E變化不明顯,此時植株的水分代謝活動沒有受到影響。25、75 mg·L-1鎘脅迫下葉片E均顯著低于同期對照組,且下降幅度隨脅迫時間的延長而增大,其中25 mg·L-1鎘脅迫結(jié)束時葉片E比對照組降低了55.10%,而75 mg·L-1鎘脅迫至17 d時葉片E即比對照組顯著降低了60.12%。
2.4.3氣孔導(dǎo)度(Gs) 氣孔是植物葉片與大氣進行氣體交換的重要通道,其閉合程度是決定葉片蒸騰速率的重要因素之一。鎘脅迫下梭魚草葉片Gs與E的變化趨勢基本一致(圖3C)。脅迫結(jié)束時,5、25 mg·L-1鎘脅迫下葉片Gs比對照組分別顯著下降了39.38%、51.40%。當(dāng)鎘濃度繼續(xù)增大時,葉片Gs下降幅度進一步增加,葉片E的變化也證明了這一結(jié)果。
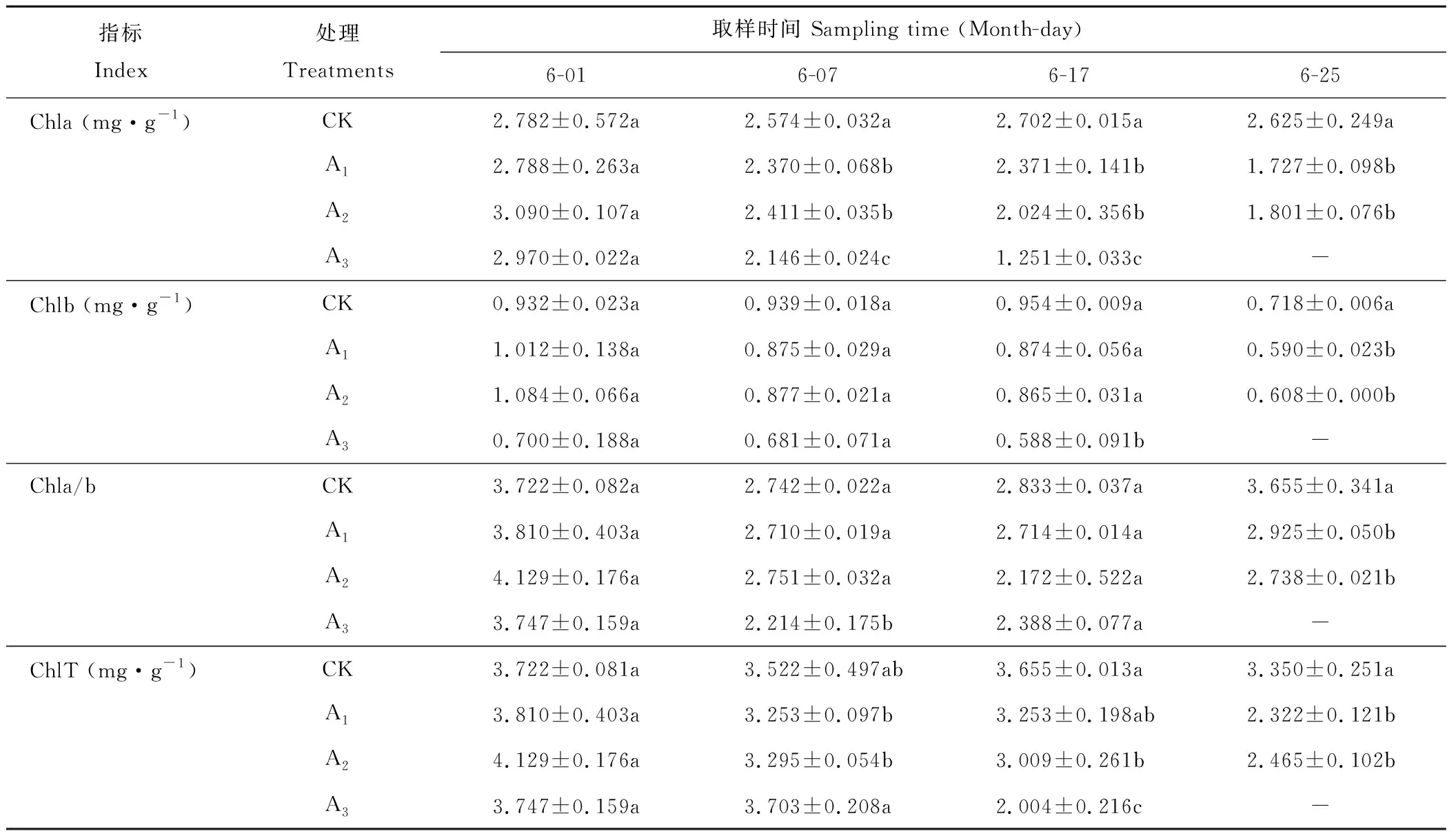
表1 不同濃度鎘脅迫下梭魚草葉片Chla、Chlb、ChlT含量及Chla/b的變化Table 1 Effects of Cd on Chla, Chlb, ChlT contents and Chla/b in P. cordata’s leaves
注:數(shù)據(jù)為平均值±平均誤差(n=3),同一列內(nèi)不同小寫字母表示0.05水平顯著差異性(P<0.05)。下同。
Note: Data are mean±SD (n=3). Different small letters in the same row indicate significant differences at theP<0.05 level. The same below.
2.4.4胞間二氧化碳濃度(Ci) 與對照相比,各處理下梭魚草葉片Ci與Pn變化趨勢較為一致(圖3D)。第7天時鎘脅迫下葉片Ci沒有明顯變化;第17和25天時鎘脅迫下葉片Ci均顯著低于對照組,其中5、25 mg·L-1鎘脅迫結(jié)束時葉片Ci比對照組分別降低了21.90%、25.25%,這可能是由葉片Gs的下降而導(dǎo)致的。
2.4.5飽和蒸氣壓虧缺 (VPD)VPD是影響植物葉片蒸騰作用的主要因素之一。隨著鎘濃度的增加和脅迫時間的延長,梭魚草葉片VPD呈上升趨勢(圖3E)。其中,5、25 mg·L-1鎘脅迫結(jié)束時葉片VPD比對照分別升高了19.28%、41.77%,75 mg·L-1鎘脅迫下葉片VPD第17天時比對照顯著升高了22.50%,這可能會使其通過Gs而間接抑制光合作用。
2.5 鎘脅迫對梭魚草葉片葉綠素?zé)晒馓匦缘挠绊?/h3>
2.5.1鎘脅迫下梭魚草葉片F(xiàn)o的變化Fo為葉片暗適應(yīng)后的最小熒光強度,其值的大小與Chla的含量有關(guān),可反映環(huán)境脅迫對植物造成的傷害程度。如表2所示,與對照組相比,隨著鎘脅迫程度的增加和時間的延長,各處理下梭魚草葉片F(xiàn)o變化趨勢有所不同。當(dāng)鎘濃度為5 mg·L-1時,葉片F(xiàn)o與同期對照組差異不顯著;當(dāng)鎘濃度為25、75 mg·L-1時,葉片F(xiàn)o呈上升趨勢,表明鎘降低了梭魚草葉片葉綠素原初光能轉(zhuǎn)化效率下降,誘發(fā)光抑制,導(dǎo)致光系統(tǒng)Ⅱ(PSⅡ)反應(yīng)中心失活。
2.5.2鎘脅迫下梭魚草葉片F(xiàn)m的變化Fm為葉片暗適應(yīng)后的最大熒光強度,可以反映環(huán)境脅迫下通過葉片PSⅡ反應(yīng)中心電子傳遞的最大潛力。與對照組相比,當(dāng)鎘濃度為5、25 mg·L-1時,葉片F(xiàn)m變化不明顯;當(dāng)鎘濃度為75 mg·L-1時,葉片F(xiàn)m呈上升趨勢,第17天時比對照組增加了6.71%(表2),表明鎘脅迫下梭魚草葉片PQ庫增加,有活力反應(yīng)中心的數(shù)量顯著增加。
2.5.3鎘脅迫下梭魚草葉片可變熒光強度的變化Vj、Vi分別為植物葉片在J、I相時的相對可變熒光強度,反映了電子經(jīng)過質(zhì)體醌A(QA)、質(zhì)體醌B(QB)時的能量耗散比率。隨著鎘濃度的增加和脅迫時間的延長,梭魚草葉片Vj、Vi均呈上升趨勢(表2)。鎘脅迫至17 d時,25、75 mg·L-1下葉片Vj比對照組分別增加了26.19%、19.05%;Vi比對照組分別增加了14.47%、10.53%,表明鎘脅迫下梭魚草葉片PSⅡ反應(yīng)中心受體側(cè)光合電子傳遞活性被抑制,這與質(zhì)體醌QA和光系統(tǒng)I(PSI)的氧化還原反應(yīng)受阻有關(guān)。
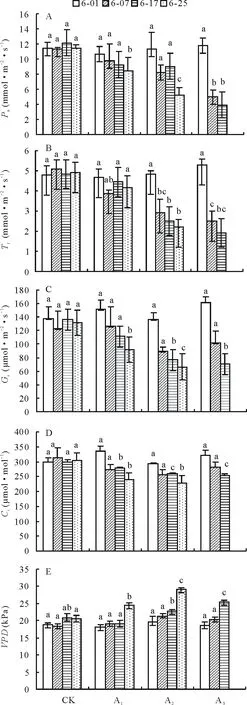
圖3 鎘脅迫對梭魚草葉片光合氣體交換參數(shù)的影響Fig.3 Effects of Cd on photosynthetic gas exchange parameters in P. cordata’s leaves
2.5.4鎘脅迫下梭魚草葉片F(xiàn)v/Fm的變化Fv/Fm為植物葉片暗適應(yīng)后PSⅡ的最大光化學(xué)效率,可以反映環(huán)境脅迫下葉片光抑制的程度。與對照組相比,不同濃度鎘處理下梭魚草葉片F(xiàn)v/Fm的變化趨勢存在一定的差異(表2)。當(dāng)鎘濃度為5、75 mg·L-1時,葉片F(xiàn)v/Fm的變化不明顯;當(dāng)鎘濃度為25 mg·L-1時,葉片F(xiàn)v/Fm呈下降趨勢,第17天時比對照組降低了5.95%,表明鎘脅迫降低了梭魚草葉片PSⅡ光化學(xué)活性及能量轉(zhuǎn)化效率,這可能是因為鎘抑制了PSⅡ反應(yīng)中心QA到QB的電子傳遞,使非還原性QB積累較多,從而降低PSⅡ反應(yīng)中心放氧活性。
2.5.5鎘脅迫下梭魚草葉片F(xiàn)m/Fo和Fv/Fo的變化Fm/Fo為植物葉片PSⅡ反應(yīng)中心電子傳遞效率,其可以反映植物對光能的利用效率。由表2可知,當(dāng)鎘濃度為5、75 mg·L-1時,梭魚草葉片F(xiàn)m/Fo沒有明顯的變化。第17天時,25 mg·L-1鎘脅迫下葉片F(xiàn)m/Fo比對照顯著下降了24.22%,這是由Fo的上升幅度大于Fm而引起的,表明鎘脅迫導(dǎo)致梭魚草葉片光能利用效率降低。同時,鎘脅迫下梭魚草葉片F(xiàn)m/Fo與Fv/Fo(PSⅡ反應(yīng)中心的潛在活性)的變化規(guī)律一致,即隨著葉片F(xiàn)m/Fo的降低,PSⅡ反應(yīng)中心的潛在活性呈下降趨勢,從而導(dǎo)致光能轉(zhuǎn)化為活躍化學(xué)能的效率降低。
2.5.6鎘脅迫下梭魚草葉片光能的吸收、傳遞與轉(zhuǎn)化 不同濃度鎘對梭魚草葉片單位反應(yīng)中心各種量子效率的影響如表2所示。第17天時25 mg·L-1鎘脅迫下梭魚草葉片單位反應(yīng)中心吸收的能量(ABS/RC)、反應(yīng)中心捕獲的能量(TRo/RC)以及用于電子傳遞的能量(ETo/RC)比對照組增加了32.97%、42.48%和28.57%,且差異均達到顯著水平,但同時單位反應(yīng)中心熱耗散的能量(DIo/RC)也比對照組顯著增加了36.36%,與75 mg·L-1鎘脅迫下葉片單位反應(yīng)中心量子效率的變化趨勢一致,表明鎘脅迫下梭魚草葉片捕光天線面積、反應(yīng)中心捕獲電子能力及進入電子傳遞鏈的能量增加,同時也有較多能量以熱耗散的形式被釋放,這可能是鎘脅迫下梭魚草進行自我保護的一種機制。
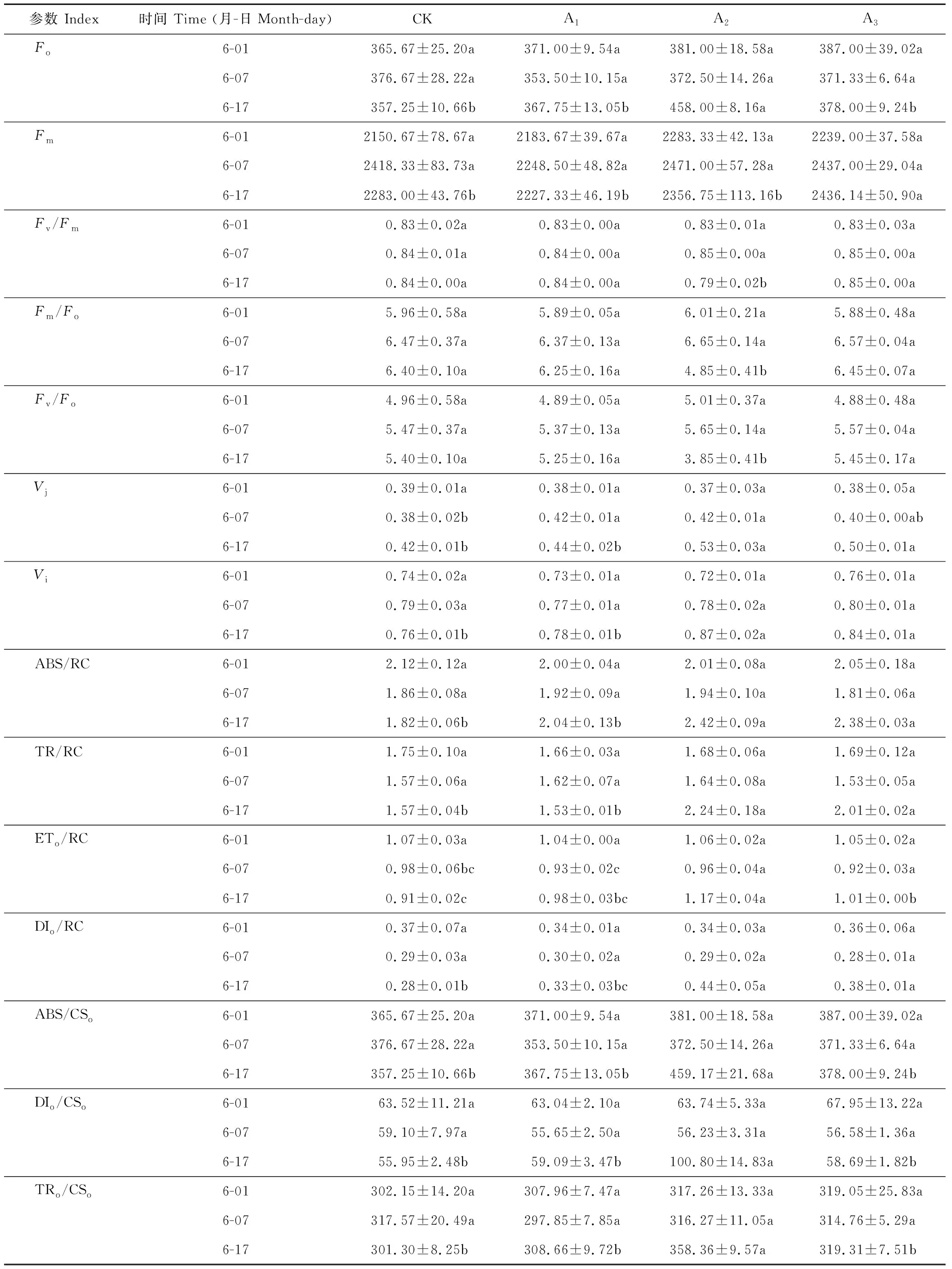
表2 鎘脅迫下梭魚草葉片葉綠素?zé)晒鈪?shù)的變化Table 2 Variations of chlorophyll fluorescence parameters in P. cordata’s leaves under Cd stress

續(xù)表2 Continued Table 2
不同濃度鎘對梭魚草葉片單位面積各種量子效率的影響存在一定的差異(表2)。第17天時25 mg·L-1鎘脅迫下梭魚草葉片單位面積吸收的能量(ABS/CSo)、單位面積捕獲的能量(TRo/CSo)比對照組顯著增加了28.53%、18.94%,單位面積熱耗散的能量(DIo/CSo)比對照組增加了80.16%,而單位面積用于電子傳遞的能量(ETo/CSo)、單位面積傳遞到電子鏈末端的能量(RE/CSo)則表現(xiàn)出相反的變化趨勢,比對照組分別顯著下降13.50%、31.50%;75 mg·L-1鎘脅迫下葉片單位面積量子效率也表現(xiàn)出相似的變化趨勢,表明鎘脅迫下葉片單位面積有活性反應(yīng)中心的數(shù)量增加,但仍會導(dǎo)致大部分的能量被熱淬滅,從而抑制單位面積光能利用率、電子傳遞和PSI氧化還原過程。
2.5.7鎘脅迫下梭魚草葉片性能指數(shù)(PIabs)的變化 PIabs反映了植物葉片PSⅡ的總體性能,可以反映逆境脅迫下葉片PSⅡ功能與結(jié)構(gòu)的變化。從表2可以看出,與對照組相比,隨著鎘濃度的增加和脅迫時間的延長,PIabs基本上呈下降趨勢,鎘脅迫下葉片PIabs在第17天比對照組分別下降了14.11%、57.82%和42.57%,表明鎘脅迫導(dǎo)致梭魚草葉片PSⅡ功能與結(jié)構(gòu)受到了不同程度的損害,不利于光能向活躍化學(xué)能進行轉(zhuǎn)變。
3 討論
植物細胞膜為細胞生命活動提供相對穩(wěn)定的內(nèi)環(huán)境,是細胞內(nèi)外物質(zhì)運輸與交換的重要屏障。當(dāng)植物遭受逆境脅迫時,體內(nèi)活性氧自由基積累,膜質(zhì)過氧化作用加強,導(dǎo)致細胞膜透性增大,胞內(nèi)電解質(zhì)大量外滲,電導(dǎo)率增加。本研究中,高濃度(75 mg·L-1)鎘處理下梭魚草葉片相對電導(dǎo)率顯著增加,這與曲丹陽等[20]的研究結(jié)果相一致,表明鎘脅迫下葉片細胞膜結(jié)構(gòu)損傷,膜系統(tǒng)的完整性被破壞;低濃度(5 mg·L-1)鎘脅迫下葉片相對電導(dǎo)率能夠保持較長時間的相對穩(wěn)定性,這可能與鎘誘導(dǎo)下梭魚草體內(nèi)抗氧化酶活性處于相對穩(wěn)定的狀態(tài)(圖2)及根系的早期調(diào)控作用有關(guān)[21]。
植物體內(nèi)存在著多種保護酶可以緩解或降低活性氧自由基對其造成的傷害[22-23],其中SOD是植物體內(nèi)清除自由基的首要物質(zhì),能夠?qū)2-·轉(zhuǎn)化為H2O2和O2,而POD和CAT則可以將H2O2分解為H2O和O2,從而降低活性氧的含量[24-25]。當(dāng)鎘濃度超過一定范圍之后,植物自身的調(diào)節(jié)作用將被破壞,導(dǎo)致細胞H2O2積累過多,從而加劇細胞膜受損程度,如蠶豆(Viciafaba)在50 μmol·L-1Cd2+處理9 d時,葉片SOD活性顯著高于其他處理,同時葉片POD活性上升,但CAT活性被抑制,導(dǎo)致細胞內(nèi)H2O2大量積累,從而對細胞產(chǎn)生毒害作用[26]。本研究中,鎘脅迫下梭魚草葉片SOD、POD及CAT活性均表現(xiàn)出不同的變化趨勢,其中CAT活性變化基本不明顯,而SOD、POD對鎘脅迫的響應(yīng)較為強烈,協(xié)調(diào)參與O2-·、H2O2的清除,以降低鎘對梭魚草膜系統(tǒng)的傷害程度,故低濃度(5 mg·L-1)鎘對梭魚草細胞的毒害作用較小,與其他學(xué)者在萬壽菊(Tageteserecta)[27]、水稻(Oryzasativa)[28]等植物上的研究結(jié)果較為類似。但是,隨著鎘濃度的增加和脅迫時間的延長,梭魚草細胞受害程度加劇,表明此時活性氧自由基的產(chǎn)生大于保護酶系統(tǒng)的清除能力[29]。
葉綠素是植物進行光合作用的重要物質(zhì)基礎(chǔ),其含量的多少反映了植物光合效率的高低。研究表明,鎘脅迫會使植物葉片葉綠素含量降低[30-31],這可能是因為鎘脅迫阻礙了葉綠素分子結(jié)合色素-蛋白復(fù)合物的過程[32],也可能是鎘脅迫通過抑制Fe2+、Mn2+及Mg2+等離子吸收和5-氨基酮戊酸脫水酶(ALAD)活性或提高葉綠素分解酶活性等途徑干擾葉綠素的生物合成[33-34]。Ebrahim[35]指出,葉片衰老過程中Chla比Chlb含量下降速率更快。本研究發(fā)現(xiàn),隨著脅迫程度的增大和時間的延長,梭魚草葉片ChlT含量呈下降趨勢,葉片Chla含量顯著下降的時間早于Chlb,同時Chla/b的變化反映出Chla含量的下降大于Chlb含量,與Xue等[36]對鎘脅迫下大豆(Glycinemax)幼苗葉片Chla、Chlb含量及Chla/b比值變化的研究結(jié)果相一致。葉綠素a是光能的“捕捉器”和“轉(zhuǎn)換器”,其含量降低會抑制梭魚草葉片光合作用,使植株出現(xiàn)葉片發(fā)黃、凋落甚至死亡現(xiàn)象。
鎘積累會導(dǎo)致葉片PSⅡ反應(yīng)中心受損,如D1蛋白質(zhì)降解[37]或水裂解蛋白酶活性下降[38],從而抑制植物光合作用[39-40]。本試驗中,隨著鎘濃度的增加和脅迫時間的延長,梭魚草葉片Pn表現(xiàn)為下降趨勢,表明鎘脅迫導(dǎo)致梭魚草葉片光合碳同化的能力下降,同時葉片Pn的下降也伴隨著E、Gs及Ci的下降,這與其他學(xué)者對鎘脅迫下玉米(Zeamays)[41]、水龍(Jussiaearapens)[42]等植物葉片光合氣體交換參數(shù)變化的研究結(jié)果相一致。氣孔是植物與大氣進行氣體交換的主要通道,為植物葉片的光合作用提供CO2,因此鎘脅迫下梭魚草葉片Pn的降低與Gs的下降有著直接的關(guān)系[44],同時也可能與葉綠體ATP合酶和硝酸還原酶活性的降低有關(guān)[44-45]。
葉綠素?zé)晒馓匦允侨~片PSⅡ光化學(xué)過程的指示器,可以反映逆境對PSⅡ活性、電子傳遞及能量分配的影響。研究表明,鎘離子(Cd2+)能夠替代葉綠素分子中心位置的鎂離子(Mg2+),從而改變?nèi)~綠素?zé)晒鈪?shù)[46]。本試驗中,在前期(第1,7天)各處理對梭魚草葉片葉綠素?zé)晒馓匦曰旧蠜]有影響,表明梭魚草對鎘具有一定的耐受性;但隨著脅迫時間的推移,25、75 mg·L-1鎘脅迫下梭魚草葉片F(xiàn)o、Vj及Vi明顯上升,F(xiàn)v/Fm、Fm/Fo、Fv/Fo和PIabs則呈下降趨勢,其中PIabs比Fv/Fm更能準確反映PSⅡ活性的變化,與Von Heerden等[47]的研究結(jié)果一致,這意味著梭魚草葉片光合作用發(fā)生了光抑制,光合電子傳遞系統(tǒng)的活性、光能利用及轉(zhuǎn)化效率降低,但其可以通過激活未活化的反應(yīng)中心來適應(yīng)鎘脅迫,具體表現(xiàn)為葉片F(xiàn)m的上升。
合理的能量分配機制是植物適應(yīng)逆境脅迫的重要策略,同時也是光保護的重要機制之一。本試驗中,后期(第17天時)25 mg·L-1鎘脅迫顯著影響了梭魚草葉片PSⅡ反應(yīng)中心光能的分配參數(shù)。首先,由于光抑制導(dǎo)致梭魚草葉片ABS/RC、TRo/RC及ETo/RC顯著增加,過剩激發(fā)能通過非光化學(xué)途徑進行耗散以減少鎘脅迫下活性氧自由基的產(chǎn)生及緩解其對光合機構(gòu)的破壞,從而促使單位反應(yīng)中心能量耗散(DIo/RC)的上升,與Ali等[48]的研究結(jié)果相一致,這對于維持梭魚草葉片光能吸收和利用之間的平衡及光合碳同化能力具有重要意義。其次,與對照組相比,梭魚草葉片ABS/CSo顯著增加,從而導(dǎo)致單位面積上用于還原QA的激發(fā)能(TRo/CSo)增加。鎘脅迫下葉片PSⅡ反應(yīng)中心過剩的激發(fā)能以電子的形式存在于電子傳遞鏈中,本研究中梭魚草葉片ETo/CSo和RE/CSo顯著下降,導(dǎo)致葉片DIo/CSo顯著增加,表明鎘脅迫下梭魚草葉片光能利用效率降低,但其可通過增加熱耗散來降低PSⅡ反應(yīng)中心過剩的激發(fā)能,從而緩解PSⅡ反應(yīng)中心光抑制[49]。
4 結(jié)論
本研究表明,梭魚草可以通過調(diào)節(jié)保護酶系統(tǒng)以適應(yīng)低濃度(5 mg·L-1)鎘脅迫。由于保護酶系統(tǒng)的調(diào)節(jié)作用有限,隨鎘濃度的增大和處理時間的延長,梭魚草葉片細胞膜結(jié)構(gòu)完整性被破壞;同時葉片光合作用發(fā)生光抑制,這可能與葉綠素含量、氣孔導(dǎo)度下降以及PSⅡ活性受損有關(guān),但其能夠激活未活化的反應(yīng)中心以適應(yīng)鎘脅迫,并通過非光化學(xué)淬滅能量以保護光合機構(gòu)免受破壞。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
