朱頂紅染色體核型分析及倍性鑒定
2018-09-09 05:26:14張永春姜紅紅李青竹楊柳燕
上海農業學報 2018年4期
李 心,張永春,姜紅紅,孫 翊,李青竹,楊柳燕
(上海市農業科學院林木果樹研究所,上海201403)
朱頂紅為石蒜科朱頂紅屬(HippeastrumHerb.)多年生球根花卉,是本屬原生種和現代改良園藝雜交種的總稱[1-2]。朱頂紅原產于南美洲,17世紀傳入歐洲和北美,20世紀初引入中國,目前全球約有75個原生種和超過600個園藝品種[3-4]。朱頂紅花大色艷,形態優美,觀賞價值極高,不僅可以作為重要的盆栽和切花材料,還可以作為露地景觀栽植,具有極其廣泛的應用價值[5]。
植物細胞染色體的觀察與分析技術是細胞遺傳學一項基本技術,多被應用于染色體核型分析[6-7]、倍性鑒定[8-9]等生物學領域。針對染色體形態進行的核型分析有助于了解生物的遺傳物質組成、變異規律、物種起源、進化關系等,在細胞分類學、染色體工程、品種間親緣關系鑒定中具有重要地位[10]。依據染色體計數進行的倍性鑒定方法是目前最直接、最可靠、應用最廣泛的鑒定方法,在輔助植物雜交育種研究和植物遺傳學研究中具有重要的應用價值[11]。
迄今為止,國內外鮮見關于朱頂紅染色體制片技術及核型分析的報道。1994年,鄒琦麗等[12]通過對朱頂紅原生種‘線縞華胄’(H.rutilum)的染色體核型分析,發現其染色體數目為33條,核型公式為K(2n)=3x=33=12 m+9 sm+12 st,被鑒定為三倍體朱頂紅。2013年,陳瑞嬌[13]通過0.1%秋水仙素溶液預處理朱頂紅根尖進行常規壓片,對白肋朱頂紅(H.reticulatumvar.striatifolium)進行染色體制片,核型分析結果表明,白肋朱頂紅的染色體數目為22條,核型公式為K(2n)=2x=8 m+6 sm+8 st,核型不對稱系數(As.K)為69.96%,核型類型為3B型,被鑒定為二倍體朱頂紅。
本試驗以朱頂紅新生根尖為材料,通過低溫與8-羥基喹啉水溶液相結合的預處理方法和HCl解離對朱頂紅的染色體制片技術進行進一步的研究,并首次對朱頂紅園藝品種‘檸檬冰糕’(‘Lemon Sorbet’)進行了染色體核型分析。同時,依據染色體計數結果對6個朱頂紅商業品種‘鳳蝶’(H.papilio)、‘特倫蒂諾’(‘Trentino’)、‘粉色驚奇’(‘Pink Surprise’)、‘快車’(‘Rapido’)、‘蘋果花’(‘Apple Blossom’)、‘櫻桃妮芙’(‘Cherry Nymph’)細胞水平進行倍性鑒定,以期為朱頂紅屬資源多樣性研究、新品種選育、親本選配等奠定細胞遺傳學基礎。
1 材料與方法
1.1 材料
取9℃冷藏處理后的朱頂紅開花球,于2015年12月種植于上海市農業科學院花卉基地塑料大棚中,待長出新根后,取新根用于染色體制片。
1.2 方法
切取朱頂紅品種‘檸檬冰糕’新生根尖1 cm左右為試驗材料,取樣時間為上午9:00—10:00;4℃條件下,用0.002 mol∕L 的8-羥基喹啉水溶液對根尖材料分別進行不同時間(1 h、3 h、6 h、9 h、12 h、24 h)的預處理;預處理后的根尖用蒸餾水清洗后置于卡諾氏固定液中,4℃固定12—24 h;蒸餾水清洗后可暫存于70%酒精中;之后取出根尖用蒸餾水清洗,在60℃條件下用1 mol∕L HCl水溶液進行解離處理,時間分別為6 min、9 min、12 min;清洗后在蒸餾水中低滲30 min;用刀片切除根冠后,再切取1 mm根尖于載玻片上,搗碎后滴加卡寶品紅染色液染色10—20 min;常規壓片后鏡檢并于100倍油鏡下拍照。探索到合適的條件后,以同樣的方法對‘鳳蝶’‘特倫蒂諾’‘粉色驚奇’‘快車’‘蘋果花’‘櫻桃妮芙’進行染色體制片。
根據染色體制片結果,對朱頂紅品種‘檸檬冰糕’進行核型分析,并通過染色體計數對‘鳳蝶’‘特倫蒂諾’‘粉色驚奇’‘快車’‘蘋果花’‘櫻桃妮芙’進行倍性鑒定。
核型分析根據李懋學等[14]制定的標準進行,使用ImageJ圖像處理軟件進行染色體配對、排列、測量等。染色體類型分類根據Levan等[15]的方法進行,核型不對稱系數采用Arano[16]的方法計算。核型類型根據Stebbins[17]的方法劃分。相關公式為:
臂比(L∕S)=長臂(L)∕短臂(S);
染色體相對長度=染色體長度∕染色體組總長度×100%;
核型不對稱系數(As.K)=長臂總長∕染色體組總長度×100%。
2 結果與分析
2.1 染色體制片
2.1.1 預處理時間
隨著0.002 mol∕L 8-羥基喹啉水溶液預處理時間的增加,染色體逐漸縮短,細胞質濃度逐漸降低,染色體分散程度越來越好(圖1)。其中1 h、3 h、6 h預處理后,染色體雖漸漸縮短,但仍然未濃縮到一定程度,相互纏繞現象嚴重;而9 h、12 h、24 h預處理后,染色體縮短程度基本相差不明顯,且分散程度良好,結構清晰,因此在朱頂紅染色體制片中,選用9—24 h預處理時間較為合適。
2.1.2 解離時間
使用1 mol∕L的HCl溶液解離6 min后,細胞分散效果良好,染色清晰,但染色體酸解程度不夠,相互纏繞現象嚴重,無法進行染色體相關分析;解離12 min后,細胞分散較好,但染色體過度酸解,結構已受到破壞,著色較淡,同樣無法進行分析;而9 min的解離時間相對適宜,細胞分散良好,染色體酸解適當,著色良好,結構清晰(圖2)。
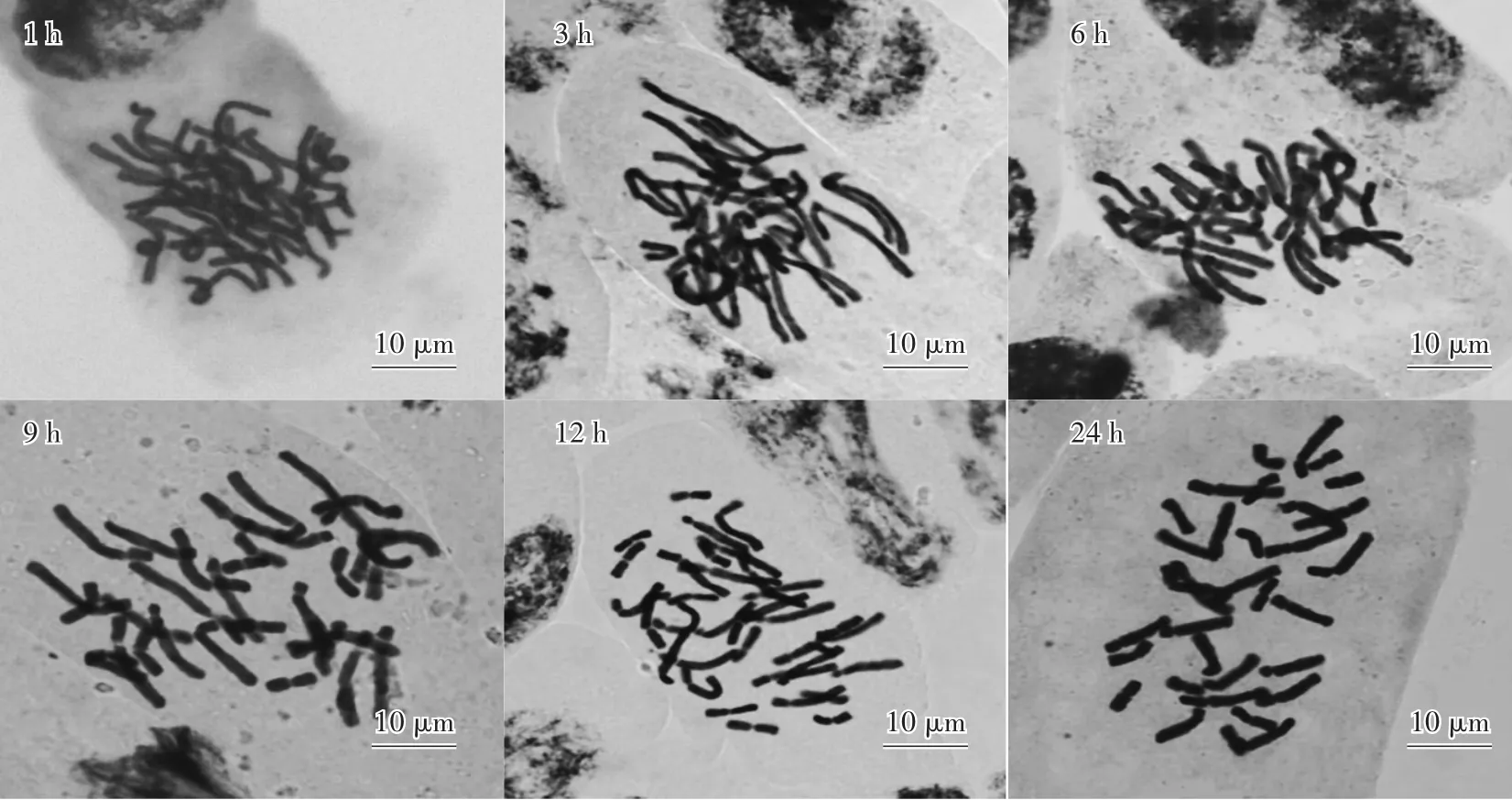
圖1 不同預處理時間下的染色體形態Fig.1 Chromosome morphology under different pretreatment time
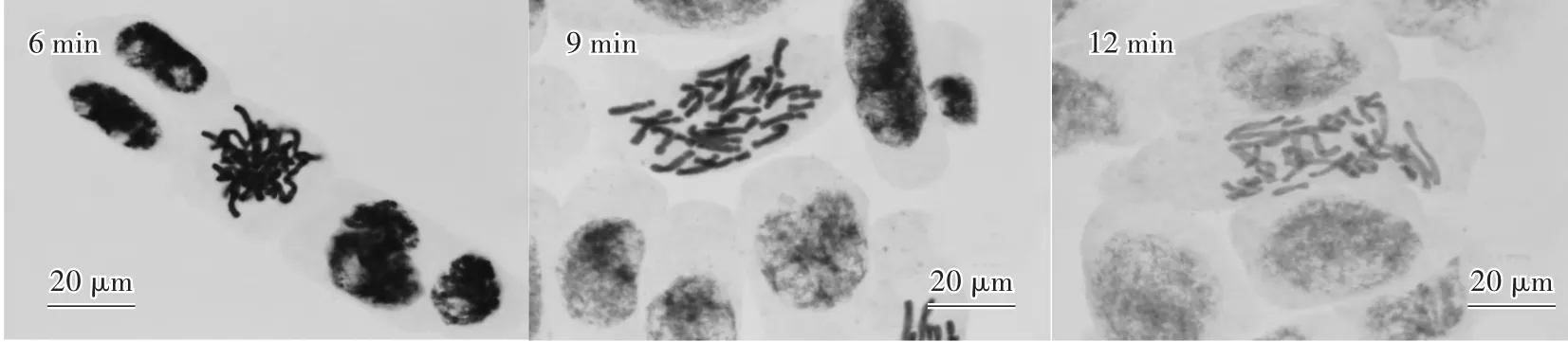
圖2 不同解離時間下的染色體形態Fig.2 Chromosome morphology under different dissociation time
2.2 核型分析
在‘檸檬冰糕’染色體制片圖中,選擇30個染色體分散良好的細胞進行計數,所有染色體數目均為33條,占計數細胞的比例為100%,因而確定‘檸檬冰糕’的染色體數目為2n=33。選擇染色體形態清晰、分散較好的分裂相進行染色體長短臂測量,根據染色體長度由長至短進行配對、排列和編號(表1、圖3—5)。

表1 ‘檸檬冰糕’染色體參數Table 1 Chromosome parameters of‘Lemon Sorbet’
根據配對結果確定‘檸檬冰糕’為三倍體朱頂紅品種,其染色體數目為2n=3x=33,具有3個染色體組,染色體基數為11。
核型分析結果表明,‘檸檬冰糕’核型公式為K(2n)=3x=33=12m+12sm+9st。其中1、2、3、4號為近中部著絲粒染色體(sm);5、6、7號為近端部著絲粒染色體(st);8、9、10、11號為中部著絲粒染色體(m)。染色體的相對長度變化范圍為5.50%—12.60%,臂比的變化范圍為1.13—3.85,最長染色體與最短染色體長度比為2.29。核型不對稱系數(As.K)為70.19%。臂比大于2∶1的染色體所占比例為63.64%,根據Stebbins的核型分類標準,‘檸檬冰糕’核型類型為3B型。
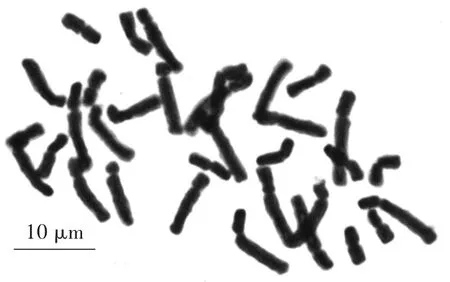
圖3 ‘檸檬冰糕’染色體形態Fig.3 Chromosome morphology of‘Lemon Sorbet’
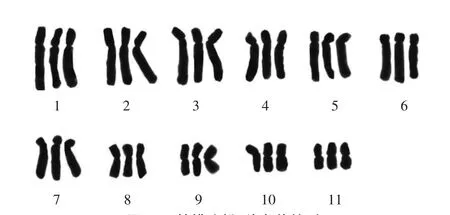
圖4 ‘檸檬冰糕’染色體核型Fig.4 Chromosome karyotype of‘Lemon Sorbet’
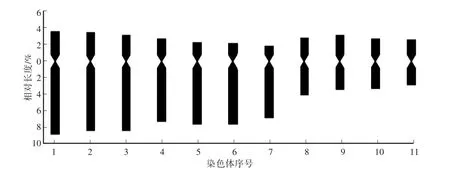
圖5 ‘檸檬冰糕’染色體核型模式圖Fig.5 Chromosome karyotype pattern of‘Lemon Sorbet’
2.3 倍性鑒定
6個朱頂紅園藝品種的染色體制片結果表明:‘鳳蝶’染色體計數為22條;‘特倫蒂諾’染色體計數為33條;‘粉色驚奇’‘快車’‘蘋果花’‘櫻桃妮芙’染色體計數均為44條(圖6)。因朱頂紅品種染色體基數為11,故‘鳳蝶’染色體數目為2n=2x=22,具有兩個染色體組,為二倍體朱頂紅;‘特倫蒂諾’染色體數目為2n=3x=33,具有三個染色體組,為三倍體朱頂紅;‘粉色驚奇’‘快車’‘蘋果花’‘櫻桃妮芙’染色體數目為2n=4x=44,具有四個染色體組,均為四倍體朱頂紅。
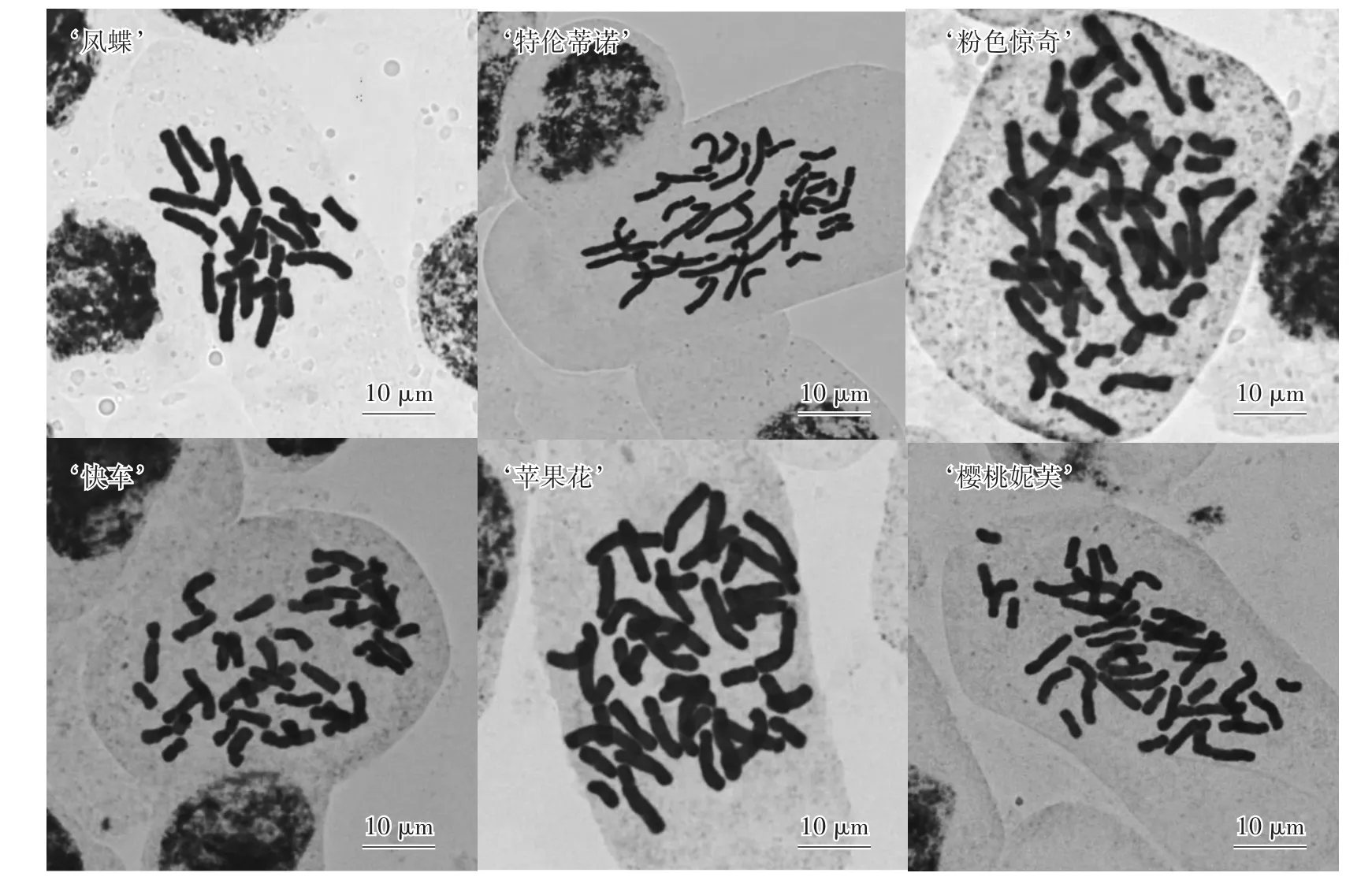
圖6 不同朱頂紅品種的染色體形態Fig.6 The chromosome morphology of different Hippeastrum cultivars
3 討論
染色體制片技術環節主要包括:取材時間與部位、預處理方法、固定時間、解離試劑與時間、低滲時間等[18]。其中預處理的主要作用是阻斷紡錘體形成、提高中期分裂相頻率、促進染色體縮短、改變胞質粘度等。目前常用的化學處理試劑為8-羥基喹啉、秋水仙素、飽和對二氯苯和α-溴萘,藜蘆堿、風油精、放線菌酮等也有少量研究者使用[19]。物理處理方法主要是0—8℃低溫處理[20]。朱頂紅的核型分析中,鄒琦麗等[12]并未提及研究中所用的預處理方法,陳瑞嬌[13]采用的是秋水仙素預處理朱頂紅根尖的方法。本研究首次使用了8-羥基喹啉與低溫處理相結合的方法,其試劑毒性相對秋水仙素較小,獲得的染色體形態分散良好、縮短長度適宜、縊痕明顯,可以在朱頂紅染色體制片研究中推廣應用。解離的作用是促進細胞分散和染色體分離。酶解主要使用的試劑是纖維素酶與果膠酶,費用相對昂貴,溫度在25—37℃,時間一般在1—4 h[6]。酸解一般使用的試劑為鹽酸,費用低廉,溫度為60℃,時間一般為5—20 min[7,20]。由此可知,酸解是一種比較高效低廉的解離手段,本研究和陳瑞嬌[13]均采用了酸解的方法,效果非常理想,因而在朱頂紅制片中應優先采用。
本研究認為朱頂紅品種‘檸檬冰糕’的核型公式為K(2n)=3x=33=12 m+12 sm+9 st,其中1、2、3、4號染色體類型為sm,5、6、7號染色體類型為st,8、9、10、11號染色體類型為m。鄒琦麗等[12]和陳瑞嬌[13]分別對朱頂紅原生種‘線縞華胄’和白肋朱頂紅進行了核型分析,其核型公式分別為K(2n)=3x=33=12 m+9 sm+12 st和 K(2n)=2x=8 m+6 sm+8 st,其中 1、2、3 號染色體類型為 sm,4、5、6、7 號染色體類型為st,8、9、10、11號染色體類型為m。本研究和前人研究結果基本一致,唯一的分歧在于4號染色體類型。前人的研究中4號染色體臂比值為3.06—3.07,而本研究中4號染色體臂比值為2.85,比值相差并不大,因而這種差異很有可能是染色體測量誤差導致。綜合這3次核型分析結果來看,朱頂紅核型不對稱系數(As.K)分別為72.59%、69.96%、70.19%,核型類型均為3B型。在生物進化過程中,染色體核型不對稱程度越高,其進化程度也越高[17],因此朱頂紅應該是植物演化史中進化程度較高的物種。另外,這3個朱頂紅品種染色體基數均為11,核型也基本一致,說明朱頂紅屬植物在親緣關系上較為接近,這與朱頂紅大部分原生種可以輕易雜交這一現象也比較一致。
本研究還對6個朱頂紅品種‘鳳蝶’‘特倫蒂諾’‘粉色驚奇’‘快車’‘蘋果花’‘櫻桃妮芙’進行了倍性鑒定。‘鳳蝶’原產巴西,是1970發現的原生種;‘特倫蒂諾’為南非園藝品種,于2007年育成;‘粉色驚奇’‘快車’‘蘋果花’‘櫻桃妮芙’是荷蘭育種家分別于2005年、2001年、1954年和2004年培育成的園藝品種。研究結果表明:‘鳳蝶’為二倍體,‘特倫蒂諾’為三倍體,‘粉色驚奇’‘快車’‘蘋果花’和‘櫻桃妮芙’均為四倍體。花瓣形態數據表明,‘粉色驚奇’‘快車’‘蘋果花’和‘櫻桃妮芙’相比‘鳳蝶’‘特倫蒂諾’花徑更大(未發表),觀賞性狀更為優良。說明朱頂紅多倍體育種在觀賞性方面改進較大,具有較為重要的意義。另外,雜交育種在朱頂紅育種工作中占有重要地位,在近年來國內外育種家的共同努力下,朱頂紅園藝品種越來越多,而對這些來源不同的品種進行的系統性歸納和研究并不多。本研究對其中1個原生種和5個園藝品種在染色體水平上進行了倍性鑒定,為雜交育種和倍性育種工作的開展提供了細胞遺傳學理論基礎。
