高產舍雷肽酶的粘質沙雷氏菌誘變選育
2018-09-06 09:09:00梅建鳳蔡少芬李靚易喻應國清
食品與發酵工業 2018年8期
梅建鳳, 蔡少芬, 李靚, 易喻, 應國清
1(浙江工業大學 藥學院,浙江 杭州,310014) 2(浙江天科高新技術發展有限公司,浙江 杭州,310012)
舍雷肽酶(serrapeptase,serratiopeptidase或serralysin),又稱沙雷肽酶、沙雷蛋白酶等,是一種堿性蛋白水解酶。該酶最初是從腸桿菌粘質沙雷氏菌(Serratiamarcescens)E15中分離出的一種胞外酶[1-3],因其強大的抗炎活性,被人們稱之為“奇跡之酶”。舍雷肽酶具有良好的抗炎、抗腫脹、促進痰液、膿液溶解與排泄以及鎮痛作用,臨床用于手術后及外傷的消炎、副鼻竇炎、乳汁潴留性乳腺炎、膀胱炎、附睪炎、智齒周圍炎及牙槽膿腫時的消炎,以及治療支氣管炎、肺結核、支氣管哮喘時痰液不易咳出等[4-5]。早在19世紀70年代就在日本上市,隨后在歐洲、亞洲等多個國家上市[6],因出色的功效和無副作用,近年來,該酶作為輔助藥物和保健食品市場需求旺盛。
舍雷肽酶主要由粘質沙雷氏菌發酵生產,目前主要生產國家是日本和印度,國內雖有一些企業在銷售舍雷肽酶,但并非采用粘質沙雷氏菌發酵,而是采用枯草芽孢桿菌發酵,嚴格來說,此類產品不能稱為舍雷肽酶。李靚[7]已從家蠶腸道中分離獲得1株產舍雷肽酶的粘質沙雷氏菌,在優化的發酵條件下,酶活單位達到1 000 U/mL以上,但隨著菌種的傳代使用,產酶活力逐漸下降至500 U/mL左右。為了提高和穩定該菌株的產酶能力,實驗采用紫外線和微波對該菌株進行連續誘變,篩選高產且傳代穩定的舍雷肽酶突變菌株。
1 材料與方法
1.1 材料與儀器
1.1.1 菌種
粘質沙雷氏菌(Serratiamarcescens)LL-413菌株,從家蠶(Bombyxmori)腸道內分離,保藏于中國典型培養物保藏中心,保藏編號CCTCC M2015780。
1.1.2 試劑與儀器
酪蛋白:生物試劑,百奧萊生物試品有限公司;酪氨酸:生物試劑,生工生物工程(上海)股份有限公司;福林酚試劑:分析純,上海滬試化工有限公司;以及其他普通分析純試劑或生物試劑。
紫外-可見分光光度計(UV2600型),尤尼柯(上海)儀器有限公司;立式壓力蒸汽滅菌器(LS-B50L型),上海華線醫用核子儀器有限公司;生化培養箱(SPX-250B型),上海博迅實業有限公司;恒溫培養搖床(HWY-2112型),上海智城分析儀器制造有限公司;微波爐(G8023CTL-K3型),中山格蘭仕家用電器有限公司。
1.1.3 培養基
平板培養基(g/L):麥芽浸粉12.2,牛肉膏6.6,酵母浸出粉11.0,NaCl 5.0,MgSO4·7H2O 1.0,瓊脂20.0,pH 8.0,121 ℃高壓蒸汽滅菌20 min。種子和產酶培養基除不加瓊脂外,其他成分同平板培養基,250 mL三角瓶裝50 mL種子或產酶培養基,8層紗布扎口,121 ℃蒸汽滅菌20 min。
1.2 方法
1.2.1 酶活的測定
采用Folin-酚法測定舍雷肽酶的活力[8]。發酵液離心后,上清液經過適當倍數稀釋后待測,取1 mL酶液和1 mL酪蛋白溶液(20 g/L,pH 8.0的Tris-HCl緩沖液配制)于試管中,充分混合后于37 ℃水浴中保溫10 min,再加入2 mL的三氯乙酸(TCA,0.4 mol/L)終止反應,繼續水浴保溫20 min,經8 000×g離心5 min。取1 mL上清液,加入5 mL的Na2CO3(0.4 mol/L)和1 mL福林酚試劑,37 ℃水浴保溫顯色30 min,以TCA滅活的酶液相同處理為參比,測定A660,依據酪氨酸濃度-A660標準曲線計算出酶解液中酪氨酸濃度,再按舍雷肽酶的活力定義計算發酵液中舍雷肽酶的活力。舍雷肽酶活力的定義:在37 ℃反應條件下,1 min水解質量濃度為20 g/L的酪蛋白產生1 μg酪氨酸的所需的酶量(mL)為1個酶活力單位(U/mL)。
1.2.2 菌種誘變
菌懸液的制備:取培養至對數生長期(16 h)的培養液1 mL,離心收集菌體,加入等量無菌生理鹽水重懸菌體,再次離心收集菌體,重復2次。用100 mL無菌生理鹽水重懸菌體于三角瓶中,加入1根無菌回形針于磁力攪拌器上攪拌20 min后備用。
紫外線誘變[9]:直徑9 cm無菌培養皿7副,分別加入制備好的菌懸液2 mL。打開培養皿蓋,在距離15 W紫外燈30 cm處邊攪拌邊照射,分別照射0、40、60、80、100、120、140 s。完畢后將各菌液用生理鹽水稀釋至103~105倍,取0.1 mL稀釋菌液涂布平板,平板黑布包裹于30 ℃下培養24 h。
微波誘變[10]:微波爐高火預熱和殺菌消毒5 min,5 mL菌懸液于試管中,試管于冰水浴中,在微波爐800 W的功率下處理0、40、60、80、100、120、140 s。完畢后將各菌液用生理鹽水稀釋103~105倍,取0.1 mL稀釋菌液涂布平板,于30 ℃培養24 h。
1.2.3 菌株的初篩與復篩
初篩:選取致死率90%以上的平板,挑取菌落轉接平板培養24 h后,挑取菌體接種產酶培養基(不做重復樣),30 ℃、200 r/min培養24 h,發酵液4 ℃、8 000×g離心5 min,測定上清液舍雷肽酶活力。
復篩:接種初篩獲得的菌株菌體至種子培養基中,30℃、200 r/min培養24 h后,按5%的體積比接入產酶培養基,30 ℃、200 r/min培養24 h后測定舍雷肽酶的活力,每個菌株做3個重復。
1.2.4 傳代穩定性試驗
將誘變后的菌株以及原始菌株LL-413在平板培養基上傳代培養,每代菌體都經過種子擴培和產酶培養(3個重復),測定產酶活力,傳代4次。
2 結果與分析
2.1 紫外線誘變
2.1.1 紫外線誘變的致死率
測定粘質沙雷氏菌在30 ℃、180 r/min條件下的生長曲線,培養16 h進入對數生長的中期,制備的菌體經不同時長的紫外線照射,測得紫外線誘變的致死率見圖1。
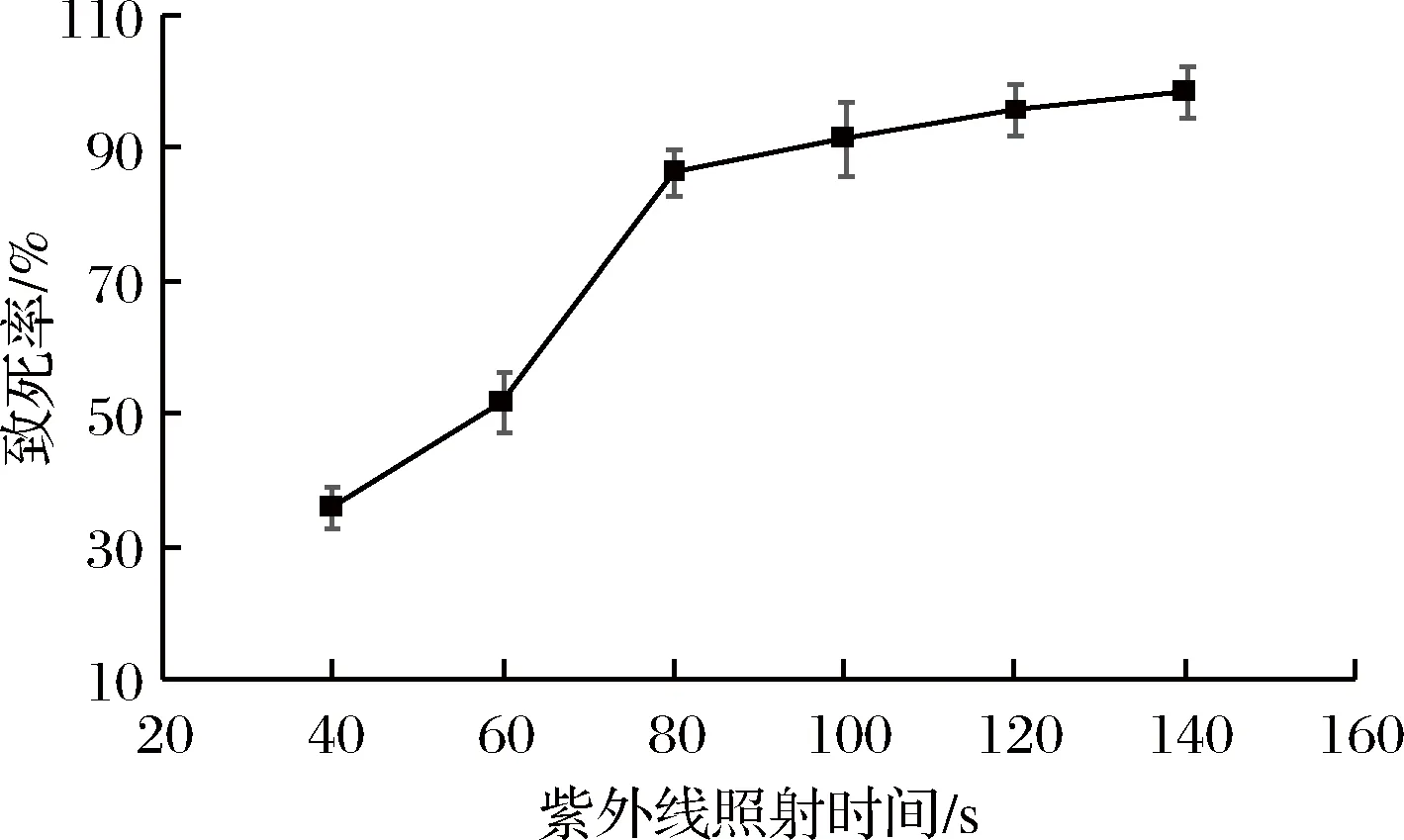
圖1 紫外線照射對粘質沙雷氏的致死率Fig.1 Fatality rate of Serratia marcescens irradiated by UV
由圖1可以看出,隨著紫外線照射時間的延長,致死率逐漸增加,當照射100 s時,致死率91.4%。一般認為致死率在90%~99.9%時,誘變效果較好[11],所以從紫外線照射時長100 s以上誘變后的平板中挑取突變株。
2.1.2 紫外線誘變菌種的篩選
從紫外線照射時長100 s以上誘變后的平板中挑取突變株50株,誘變菌株產酶培養24 h后測定舍雷肽酶活力,從中選取酶活提高30%的10個突變菌株進行復篩,復篩的10個菌株產舍雷肽酶的活性見表1。
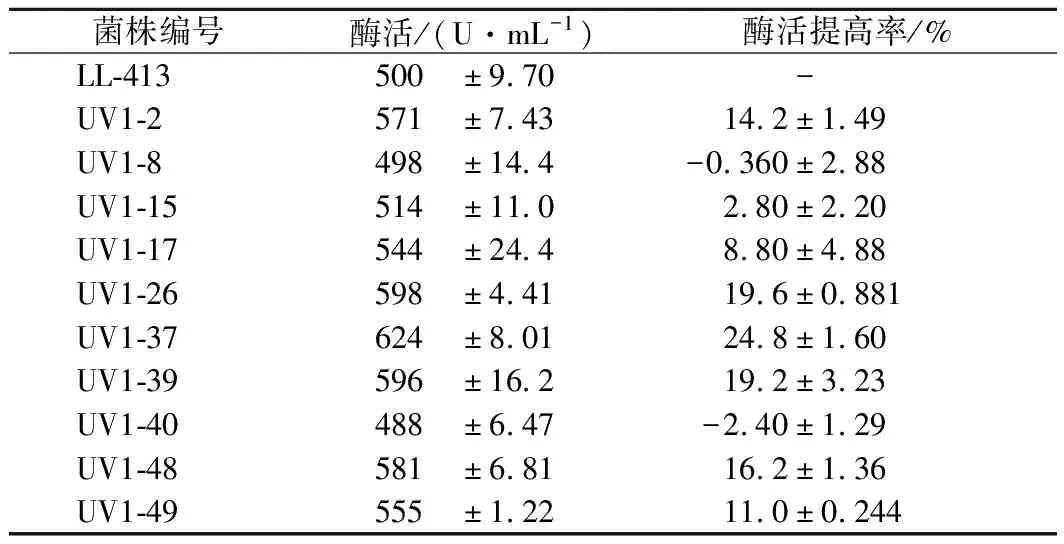
表1 紫外線誘變突變菌種的復篩產酶活力Table 1 Serrapeptase activity of the strains after the 1st UV mutation in re-screening
經過復篩后,UV1-26、UV1-37和UV1-39菌株的產酶活力提高幅度較大,且均達到了極顯著差異水平(p<0.01),其中UV1-39突變菌株不產靈菌紅素,菌落呈乳白色(圖2)。不產靈菌紅素菌株發酵生產舍雷肽酶,有利于提高酶產品的外觀品質,在篩選產酶活力提高的前提下,優先選擇此類突變株。
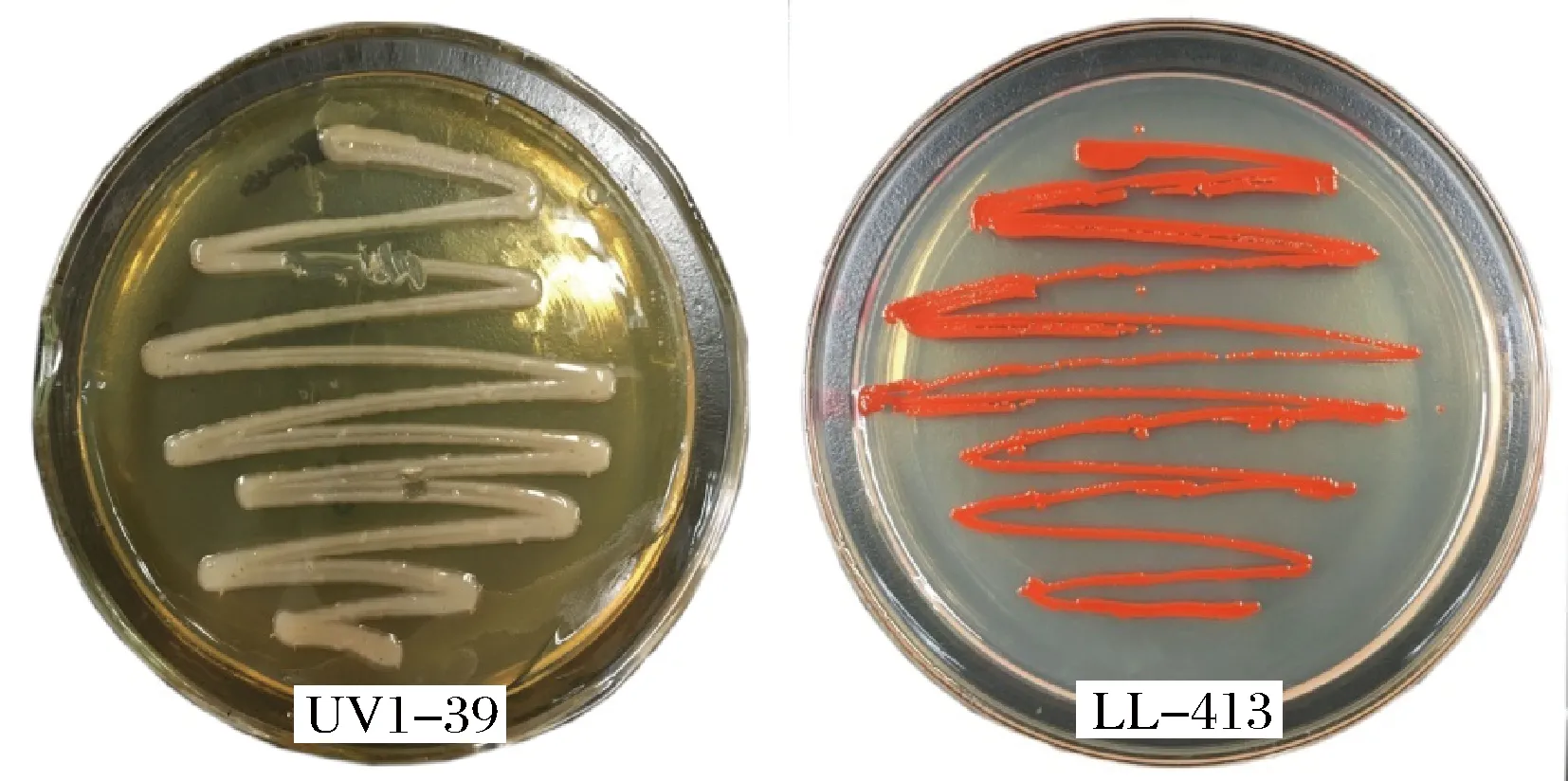
圖2 UV1-39株紫外誘變菌株和出發菌株LL-413菌落形態Fig.2 Colony morphology of strain UV1-39 and strain LL-413
2.1.3 二次紫外線誘變菌株的篩選
對復篩后的3株菌株再次用紫外線照射100 s,誘變后UV1-39為出發菌株長出的菌落均不產靈菌紅素,UV1-26和UV1-37為出發菌株長出的菌落也有少數不產靈菌紅素。從UV1-39平板中挑取了18株不產靈菌紅素的突變菌株,產酶培養24 h后檢測舍雷肽酶活力。從中選取產酶活力提高15%的6個突變菌株進行復篩,復篩的6個菌株產舍雷肽酶活力見表2。
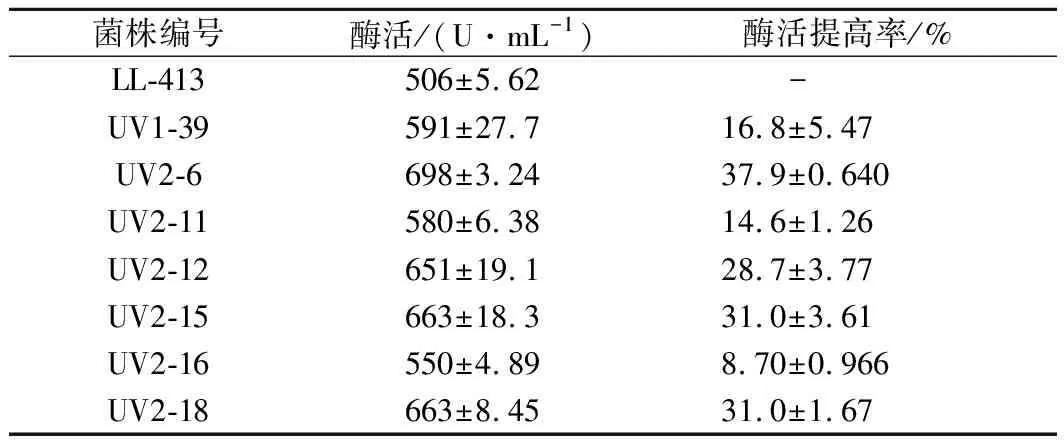
表2 二次紫外線誘變突變菌種的復篩產酶活力Table 2 Serrapeptase activity of the strains after the 2nd UV mutation in re-screening
從表2可知,UV2-6產酶活力提高幅度較大,且達到了極顯著差異水平(p<0.01),較其出發菌株UV1-39提高了18.1%,較原始出發菌株LL-413提高了37.9%,該菌株不產靈菌紅素,所以選其作為進一步微波誘變的出發菌株。
2.2 微波誘變
2.2.1 微波誘變的致死率
以UV2-6為出發菌株,在設定微波功率為800 W的條件下,采用不同時長的微波輻射,測定微波誘變對粘質沙雷氏菌的致死率,結果見圖3。
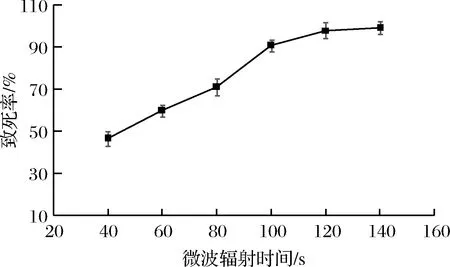
圖3 微波輻射對粘質沙雷氏的致死率Fig.3 Fatality rate of Serratia marcescens irradiated by microwave
隨著微波輻射時間的增加,菌體死亡率不斷的提高。輻射100 s時致死率為90.8%,所以從微波輻射時長100 s以上誘變后的平板中挑取突變株。
2.2.2 微波誘變突變菌種的篩選
從微波輻射時長100 s以上誘變后的平板中挑取突變株30株,搖瓶產酶培養24 h,測定各菌株產舍雷肽酶的活力,選取了產酶活力提高30%的10個菌株進行復篩,復篩的10個菌株產舍雷肽酶的活力見表3。可以看出,多數菌株產酶活力提高的幅度較大,且均達到了極顯著差異水平(p<0.01)。產酶活力提高30%的菌株有7株,說明微波對粘質沙雷氏菌誘變提高舍雷肽酶的效果非常顯著。
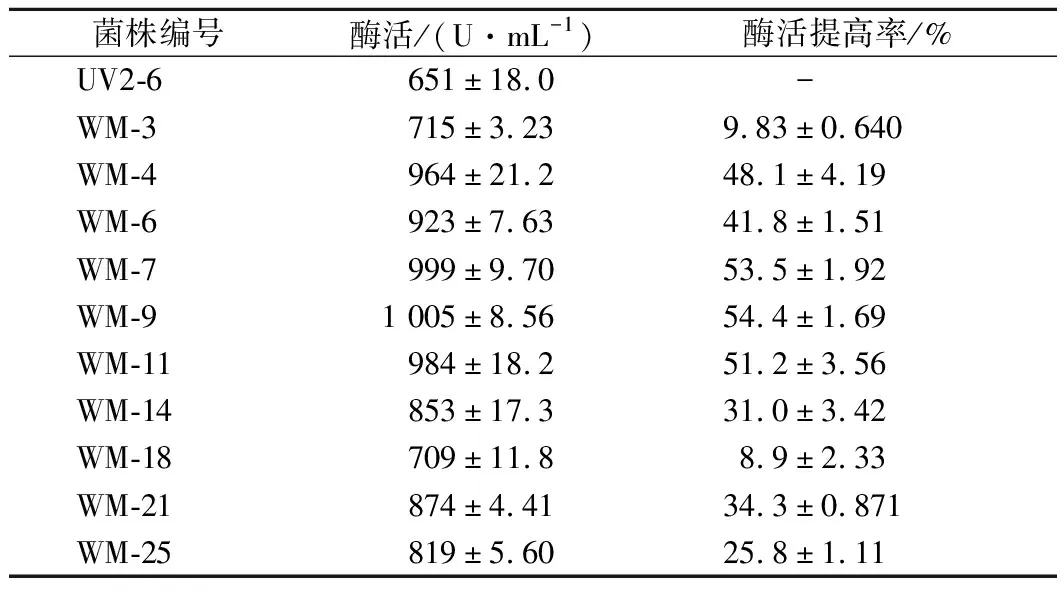
表3 微波誘變突變菌種的復篩產酶活力Table 3 Serrapeptase activity of the strains after micro-wave mutation in re-screening
2.3 突變菌株的傳代穩定性
將經微波誘變后的MW-4,MW-6,MW-7,MW-9,MW-11,MW-14,MW-21,與原始菌株LL-413一起在平板上劃線培養,傳代4次,且每代均做3個重復的搖瓶產酶培養,檢測舍雷肽酶活力,結果見表4。
從表4以看出,原始菌株LL-413穩定性較好,產舍雷肽酶活力穩定;MW-4、MW-6、MW-11和MW-21菌株所產舍雷肽酶活力逐代下降;MW-9和MW-14兩個突變菌株產酶活力穩定,分別為982 U/mL和956 U/mL,是原始菌株LL-413的1.98倍和1.92倍。
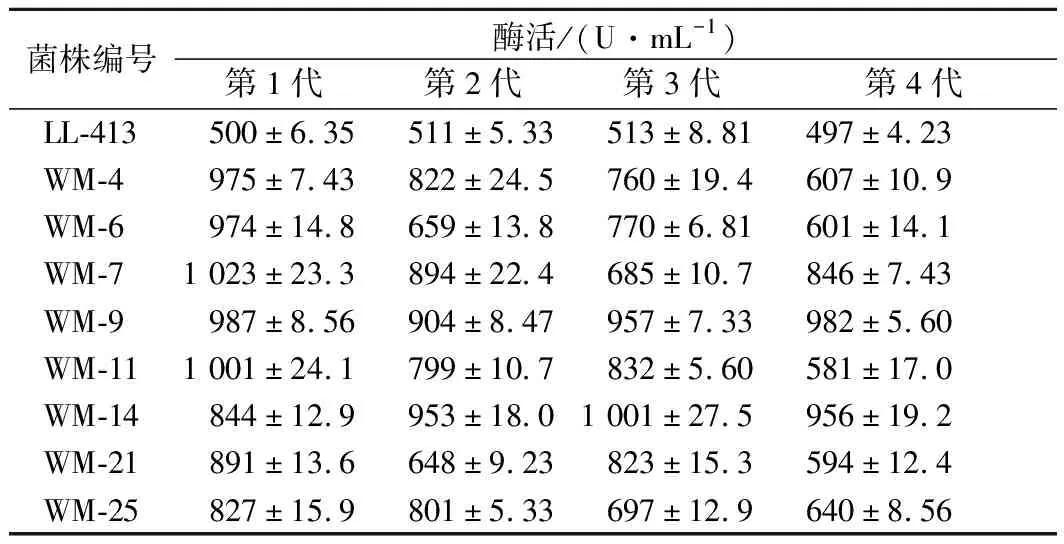
表4 突變菌株產舍雷肽酶活性的傳代穩定性Table 4 Hereditary stability of the mutant strains for producing serrapeptase
3 結論
從家蠶腸道中分離的原始菌株粘質沙雷氏菌LL-413,經紫外線和微波連續誘變后,篩選到2株產舍雷肽酶活力提高較大的突變株MW-9和MW-14,搖瓶發酵產酶活力分別為982 U/mL和956 U/mL,分別是原始菌株LL-413的1.98倍和1.92倍。目前,國內未見關于利用粘質沙雷氏菌產舍雷肽酶的研究報道;國外研究中,由粘質沙雷氏菌產舍雷肽酶活力最高報道是2 155 U/mL[12]。
此外,突變菌株MW-9和MW-14在30℃下培養均不產靈菌紅素,發酵液不再是暗紅色,這一特性有利于改善成品舍雷肽酶的外觀顏色。
