燕科和雨燕科部分鳥類系統發育比較分析
2018-08-21 02:41:52劉方慶胡夢英文隴英
野生動物學報 2018年3期
關鍵詞:物種
劉方慶 胡夢英 文隴英
(西南山地瀕危鳥類保護四川省高等學校重點實驗室,樂山師范學院生命科學學院,樂山,614000)
燕科(Hirundinidae)鳥類隸屬于雀形目(Passeriformes),分布在中國的有4屬12種,分別為河燕屬Pseudochelidon(如白眼河燕P.sirintarae等),沙燕屬Riparia(如崖沙燕R.riparia等),燕屬Hirundo(如家燕Hirundorustica等)和毛腳燕屬Delichon(如煙腹毛腳燕D.dasypus等);雨燕科(Apodidae)鳥類隸屬于雨燕目(Apodiformes),分布在中國的有5屬9種,分別為金絲燕屬Aerodramus(如短嘴金絲燕Ae.brevirostris等),針尾燕屬Hirundapus(如白喉針尾雨燕Hirundapuscaudacutus等),棕雨燕屬Cypsiurus(如棕雨燕C.balasiensis等),高山雨燕屬Tachymarptis(如高山雨燕T.melba等)以及雨燕屬Apus(如白腰雨燕Ap.pacificus等)。
傳統的宏觀分類學認為燕科和雨燕科分別隸屬于雀形目和雨燕目,其分類依據是兩者之間的形態特征和羽色特點等;有關鳥類分類及進化的結果多是從形態結構生理特征等方面獲得的,由于學者們對許多動物表型特征觀點的不一致,許多問題并沒有形成統一的結果而未能得到解決[1-2],例如,淡腰金絲燕(Aerodramusgermani)屬雨燕目雨燕科金絲燕屬,又名戈氏金絲燕,是纖小(12 cm)的深色金絲燕,尾略呈叉形,上體黑褐,腰灰白、色淡,而尾部色深,下體灰褐,腹部具淺色橫斑;國內幾乎沒有淡腰金絲燕的影像資料,有些學者把此種歸入爪哇金絲燕下的亞種Aerodramusfuciphagagermani[3-4]。但是另外一些學者將金絲燕屬(如淡腰金絲燕(又名戈氏金絲燕))歸為Collocaliagermani[5]。
隨著分子生物學技術的迅速發展,分子進化已成為近年來進化生物學研究中的一個重要方面,而分子生物學手段的應用可以為解決這一問題提供很重要的佐證[6]。
線粒體DNA(Mitochondrial DNA,mtDNA)是動物細胞核外DNA,具有分子結構簡單,母系遺傳,進化速率快,缺少重組等特點,已經成為群體遺傳結構、地理變異、系統發育等研究領域的有效分子標記[7-9]。常用來作為系統發育和系統地理研究標記的基因有Cytb,COI(線粒體細胞色素C氧化酶亞單位Ⅰ),12S rRNA,16S rRNA以及ND2、 ND3等基因。其中細胞色素b(Cytochromeb,Cytb)基因的結構和功能在mtDNA 的13個蛋白質編碼基因中被了解的最為清楚,且進化速度適中,因此是鳥類分子系統發育中常用的遺傳標記之一[10]。已有的研究結果顯示,Cytb基因的種間分歧度為1%~10%,這一分歧度既能區別物種,又可忽略多重替換[11]。近年來利用線粒體基因對不同物種的系統發育研究已經越來越成熟和普遍。
目前國內對燕科鳥類多集中在繁殖生態和繁殖習性等方面[12-14]。對雨燕科鳥類尤其是金絲燕屬鳥類的研究多集中在燕窩的營養成分品質鑒定以及回聲定位機制等方面。如研究者利用Cytb、ND2、COI、12S rRNA等條形碼序列,采用實時熒光定量PCR技術對32種不同國家和不同品種的燕窩商品進行了品質鑒別,并探討不同種類燕窩的品質分類關聯,為燕窩的品質鑒定提供更為全面的分子生物學數據。簡葉葉[15]等對燕窩的來源、營養成分和主要活性物質、功能作用等問題進行了綜述,并總結了燕窩摻偽的鑒別方法:理化鑒定、光譜鑒定、生物鑒定及其他儀器鑒別法[15-16]。此外,一些學者對短嘴金絲燕回聲定位機制及其歸巢行為進行了研究,初步闡明了此物種回聲定位的叫聲類型和頻率波段范圍[17-18]。然而對于燕科和雨燕科系統發育及其親緣關系的研究在國內幾乎未見報道。
本文通過選擇物燕科和雨燕科共13個物種,其中雀形目燕科8種,雨燕目雨燕科5種,以線粒體基因Cytb作為分子標記,通過分子生物學手段對燕科和雨燕科部分鳥類進行重建系統發育樹,以驗證分子生物學分類結果是否與宏觀形態學分類結果相一致。
1 研究方法
從NCBI(National Center for Biotechnology Information)網站中下載燕科和雨燕科13個物種線粒體Cytb序列,所獲得的物種NCBI序列號見表1。數據處理:用Bioedit[19]軟件將所有序列進行多重比對后去掉兩端多余部分,裁剪整齊,得到序列長度為999 bp,然后保存為.pir文件,再將此文件導入軟件DanSP 5.0[20]中,通過此軟件生成.nex單倍型文件。采用貝葉斯推論法(Bayesian Inference)和最大似然法(ML,Maximum Likelihood)以雁形目(Anseriformes)鴨科(Anatidae)雁屬(Anser)灰雁(Anseranser)作為外群對燕科和雨燕科13個物種進行系統發育樹重建。通過MrModeltest 2.3[21]軟件檢驗得到貝葉斯和最大似然法的最適模型為GTR+I+G。在MrBayes[22]中對Cytb設置 1百萬代運行(至sd<0.01),分析樹時均舍棄前2500顆(burnin=2500)。在PAUP 4.10[23]軟件中運行ML樹,設置重復1000代。得到的樹文件(.tre)通過Figtree[24]軟件查看并進行系統發育樹節點支持率的設置,然后導出圖片格式保存。物種兩兩之間的遺傳距離通過MEGA 6[25]中的p遺傳距離方法計算。
表1 燕科和雨燕科部分鳥類NCBI序列號
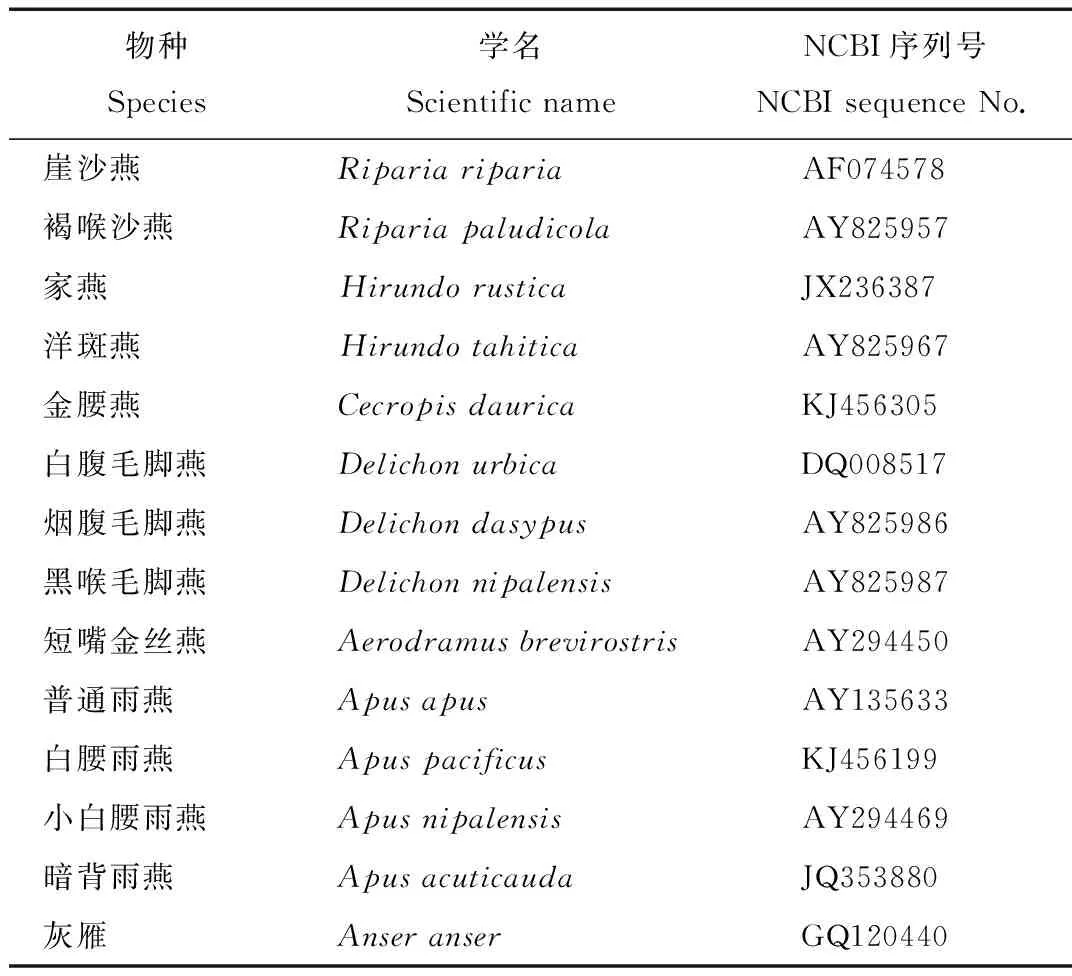
Tab.1 The NCBI sequence number of species for part ofHirundinidae and Apodidae
2 結果
2.1 堿基組成
燕科和雨燕科13種鳥類Cytb片段經過比對裁剪最后得到序列長度為999 bp(表2)。4種堿基含量平均分別為:A(27.707%)、T(24.544%)、C(34.306%)以及G(13.443%)。從堿基組成偏向性來看,A+T 含量(52.251%)略高于G+C 含量(47.749 %)(表2)。
表2 燕科、雨燕科13種鳥類Cyt b堿基含量
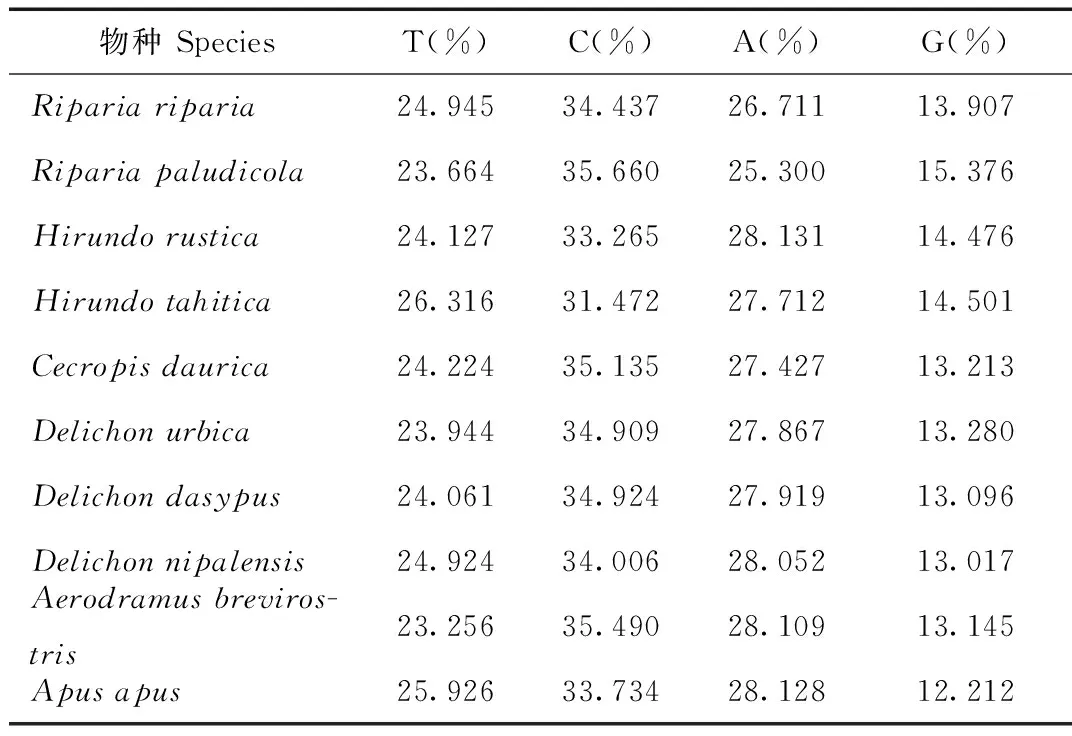
Tab.2 The contents of base pair for Hirundinidae and Apodidae
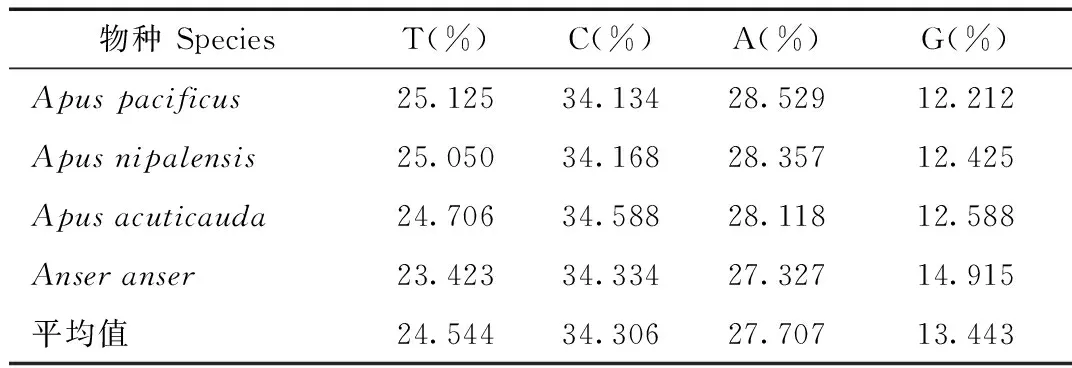
續表2
將比對并裁剪后的序列進行Modeltest檢驗后得到最大似然法(ML)和貝葉斯推論法(BI)的最適核苷酸替代模型為GTR+I+G。其中4種堿基頻率分別為A(0.3157)、T(0.1770)、C(0.4024)以及G(0.1049)。
表3 ML和 BI分析所用核苷酸替代模型及參數
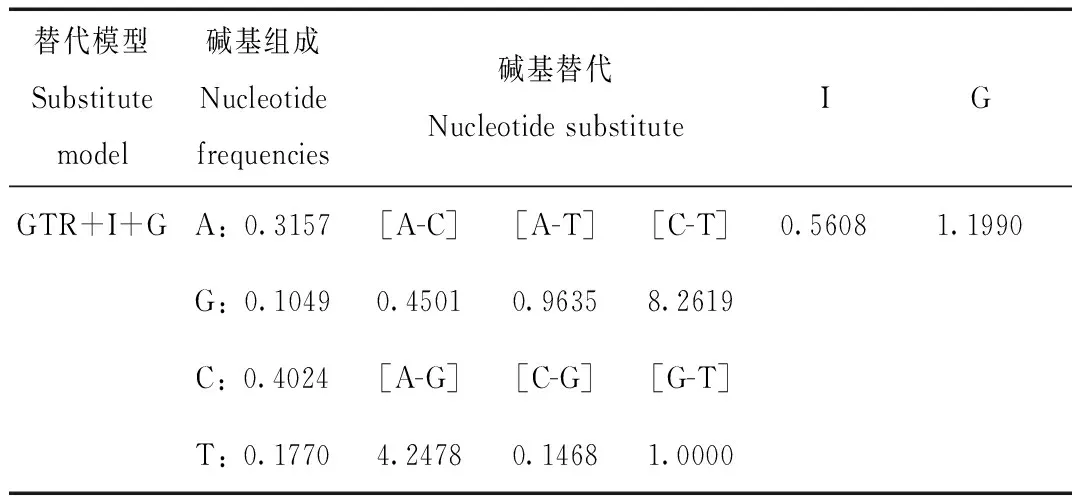
Tab.3 Models and parameters used in ML and BI analysis
注:利用hLRTs和AIC標準選擇的最佳模型都是GTR+I+G。其中G表示Gamma分布參數,I表示不變位點比例
Note:The best model of standard is GTR+I+G for hLRT and AIC.The G represents Gamma distribution shape parameter,I represents proportion of invariable sites
2.2 系統發育關系
采用貝葉斯推論法(BI)對燕科和雨燕科13種鳥類重建系統發育樹,結果表明,燕科8種鳥類和雨燕科5種鳥類分別形成兩大單獨的支系,且節點后驗概率達到了100%。其中,燕科鳥類分支中金腰燕(Cecropisdaurica)首先分化出來形成單獨的一個支系,隨后沙燕屬、燕屬和毛腳燕屬聚在一起形成一個大的分支。在雨燕科支系中,短嘴金絲燕最早分化出來形成單獨的分支,其余雨燕屬的5個種聚在一起形成一個大的支系(圖1)。
最大似然法(ML)構建的系統發育樹結果與貝葉斯推論法構建的系統發育樹具有相同的拓撲結構。燕科8種鳥類和雨燕科5種鳥類分別形成兩大單獨的支系,其中,燕科鳥類分支中金腰燕首先分化出來形成單獨的一個支系,隨后沙燕屬、燕屬和毛腳燕屬7種鳥類聚在一起形成一個大的分支。在雨燕科支系中,短嘴金絲燕最早分化出來形成單獨的分支,其余雨燕屬的5個種聚在一起形成一個大的支系(圖2)。
2.3 遺傳距離
燕科和雨燕科13個物種兩兩之間的遺傳距離結果表明,白腰雨燕和洋斑燕(Hirundotahitica)之間遺傳距離最大,達到0.197,暗背雨燕(Apusacuticauda)和白腰雨燕之間的遺傳距離最小,為0.035。燕科內部遺傳距離最大的為金腰燕和洋斑燕,兩者之間遺傳距離達到0.145,遺傳距離最小的是黑喉毛腳燕(Delichonnipalensis)和煙腹毛腳燕(D.dasypus),為0.042。雨燕科內部遺傳距離最大的為小白腰雨燕和短嘴金絲燕,其兩者之間的遺傳距離為0.096,遺傳距離最小的為白腰雨燕和暗背雨燕,兩者之間的遺傳距離為0.035。內群與外群之間遺傳距離均較大,其范圍在0.169~0.208之間。
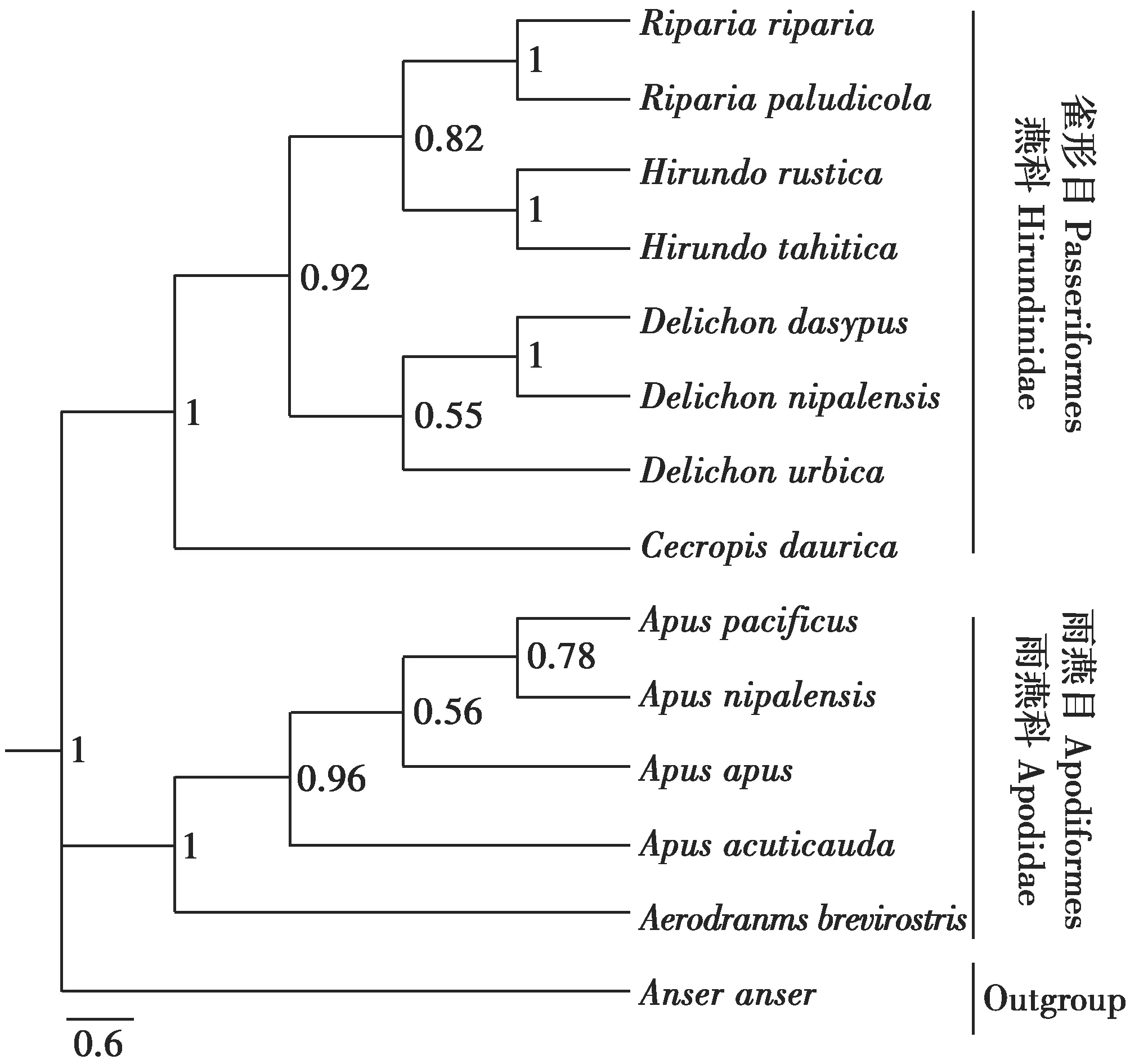
圖1 燕科和雨燕科部分鳥類系統發育樹(BI,Bayesian Inference)Fig.1 The phylogenetic tree of 13 species with Bayesianinference for Hirundinidae and Apodidae
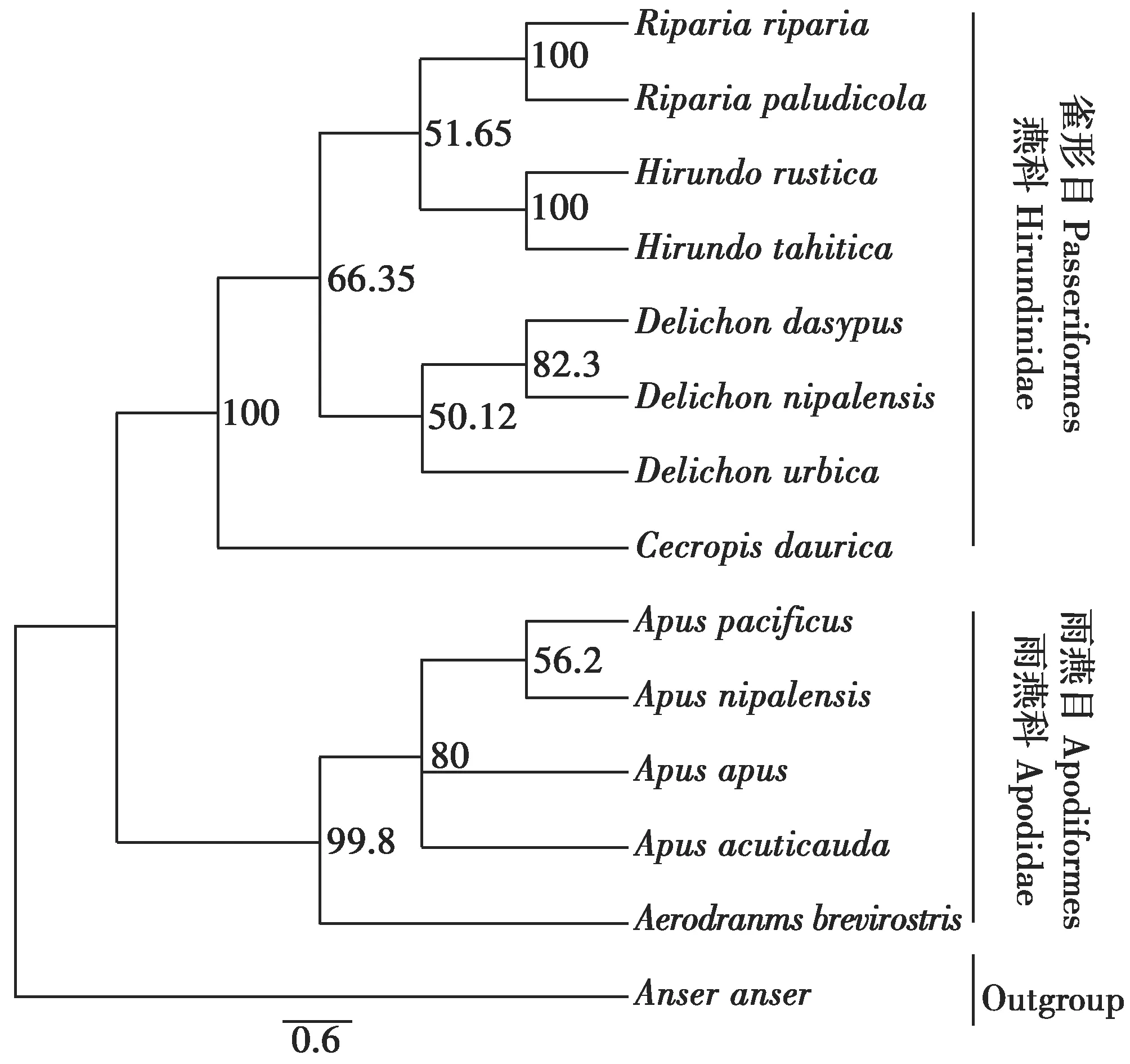
圖2 燕科和雨燕科部分鳥類系統發育樹(ML,Maximum Likelihood)Fig.2 The phylogenetic tree of 13 species with MaximumLikelihood for Hirundinidae and Apodidae
表4 燕科、雨燕科部分鳥類之間遺傳距離
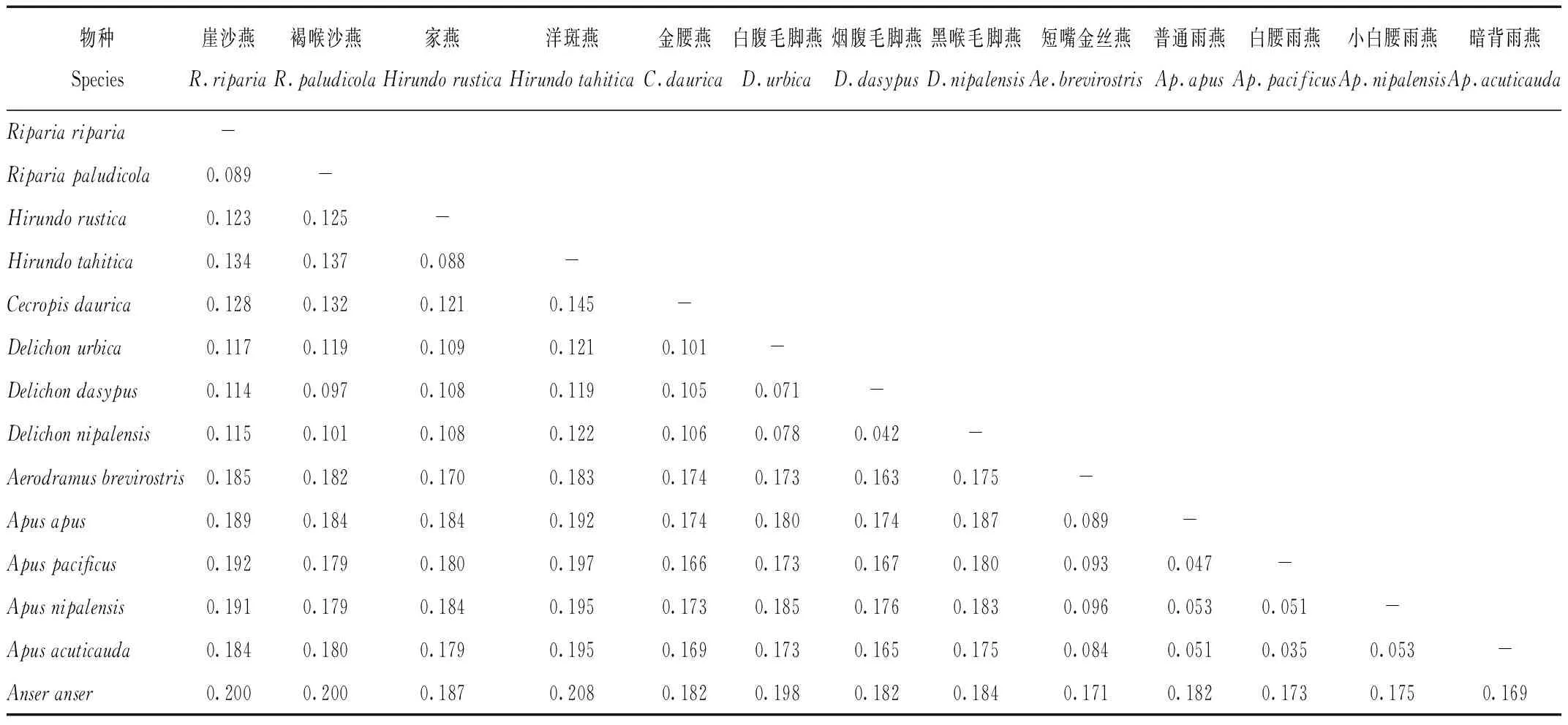
Tab.4 The genetic distance among species for Hirundinidae and Apodidae
3 討論
燕科和雨燕科13種鳥類Cytb基因分析結果顯示,4種堿基含量平均分別為:A(27.707%)、T(24.544%)、C(34.306%)以及G(13.443%)。堿基組成百分比顯示出堿基C的含量相對豐富,G的含量相對較低;所有序列嘧啶(C+T)的平均含量達58.850%,嘌呤(A+G)的平均含量為41.150%,嘧啶含量明顯高于嘌呤含量,表明堿基組成存在偏向性;且類似的堿基偏倚現象在雀形目鳥類中以及其他鳥類研究中也有發現[26-28]。本文通過對雀形目燕科和雨燕目雨燕科共13種鳥類基于線粒體基因Cytb進行系統發育樹的重建,結果表明燕科所有物種聚在一起形成單獨的一個分支,雨燕科所有物種聚在一起形成一個單獨的分支,其聚類結果與形態學分類相一致。
本文研究結果表明,燕科鳥類和雨燕科鳥類明顯分為兩個不同的支系,燕科8種鳥類單獨聚在一起,雨燕科5種鳥類聚在一起,且最大似然法和貝葉斯推理法所建系統發育樹具有相同的拓撲結構。對燕科和雨燕科鳥類進一步的遺傳距離分析表明,雨燕科和燕科之間遺傳距離較大,而各科內部種間遺傳距離較小。因此,燕科鳥類和雨燕科鳥類親緣關系較遠,分別隸屬于雀形目和雨燕目。
一直以來,金絲燕的分類系統及其物種的分類一直存在著很大的爭議。一些研究者根據金絲燕的形態特征和窩特征來分類[29-30],而另一些研究者是以金絲燕的回聲定位能力來作為分類的依據[31]。有研究者曾把所有的金絲燕歸依單一的屬名為Collocalia[32]。這個分類用了一百多年,一直到發現回聲定位能力的金絲燕。Brooke把金絲燕分為3個屬名為無回聲定位能力的Collocalia,無回聲定位能力的Hydrochous(包括巨瀑布雨燕Hydrochousgigas)和回聲定位能力的Aerodramus[33]。有關金絲燕屬內種的劃分依然存在不同的觀點,因此還需通過進一步的分子生物學手段,采用序列更長以及更多的基因作為標記來進行系統發育樹的重建,目前,雖然以線粒體DNA為標記進行系統發育和遺傳分析的研究已經越來越普遍和成熟,然而,以多個線粒體基因標記結合形成串聯序列以及線以粒基因和核基因標記相結合,特別是隨著測序技術的迅速發展,利用高通量測序技術(第二代測序技術)測定物種全基因組已越來越普遍,這些新技術的產生和發展對系統發育和譜系地理的研究起到了很大的推動作用,從而更加容易產生標記基因更豐富、序列更長以及更可靠的分析結果。因此,對燕科和雨燕科等鳥類的系統發育研究還應該以多個標記基因以及更長的序列進行分析,從而闡明金絲雀屬以及其他學界尚存在爭議的物種分類和歸屬問題。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
