鹽脅迫應答基因GmWRKY6的克隆及轉基因百脈根的抗鹽分析
2018-08-18 03:56:00柯丹霞彭昆鵬夏遠君朱玉瑩張丹丹
草業學報 2018年8期
關鍵詞:大豆
柯丹霞,彭昆鵬,夏遠君,朱玉瑩,張丹丹
(信陽師范學院生命科學學院,大別山農業生物資源保護與利用研究院,河南 信陽 464000)
WRKY轉錄因子是植物特有的一類轉錄因子,在植物響應逆境脅迫信號途徑中起著重要作用[1]。目前,大量研究報道WRKY類轉錄因子在植物抵御鹽脅迫過程中發揮關鍵調控作用。玉米(Zeamays)ZmWRKY25、45、55、67基因在鹽堿脅迫下均呈上調表達,ZmWRKY62基因呈下調表達,它們可能以不同方式參與玉米對鹽脅迫的響應[2-5]。番茄(Solanumlycopersicum)SlWRKY39基因受鹽脅迫誘導表達,其過量表達增加了植株中滲透調節物質脯氨酸的含量,抑制了丙二醛的合成,誘導了脅迫相關基因SlRD22和SlDREB2A的表達,從而增強了番茄植株對鹽害的抗性[6]。棉花(Gossypiumhirsutum)GhWRKY41基因受鹽脅迫誘導上調表達,過表達該基因顯著提髙了轉基因棉花在高鹽條件下的發芽率,提高了轉基因棉花對鹽脅迫的耐受性[7]。過表達棉花GhWRKY39-1提高了轉基因煙草的耐鹽性[8]。棉花GhWRKY11[9]、巴西橡膠樹(Heveabrasiliensis)HbWRKY75[10]、巨桉(Eucalyptusgrandis)EgrWRKY70[11]、辣椒(Capsicumannuum)CaWRKY13[12]以及茶樹(Camelliasinensis)CsWRKY57[13]等WRKY類轉錄因子基因在高鹽脅迫下均呈現明顯的誘導表達,推測這些基因可能參與鹽脅迫的應答反應與調控。
關于WRKY類轉錄因子在牧草抗逆性改良方面的研究,近年來只在紫花苜蓿(Medicagosativa)中有少量報道。前人將野生大豆(Glycinesoja)GsWRKY20基因導入豆科牧草紫花苜蓿中,發現過表達該基因不僅可以提高轉基因苜蓿的耐鹽性,還能提高轉基因苜蓿的耐旱性[14]。最新的研究將野生大豆GsWRKY15基因轉入紫花苜蓿中,發現過表達GsWRKY15基因的紫花苜蓿在堿脅迫下生長狀態更好,相對質膜透性和丙二醛含量顯著降低,而葉綠素含量顯著升高,紫花苜蓿的耐堿能力顯著增強[15]。
百脈根(Lotusjaponicus)作為一種優良的豆科牧草,在世界范圍內廣泛種植。因其固氮效率高,細胞再生性能好,遺傳轉化效率高等優勢,已經成為研究共生固氮機理[16-21]、基因異位表達[22-23]等的理想材料。雖然百脈根具有抗旱和耐鹽堿的特性,但只適宜在輕度干旱和輕度鹽堿的土地上種植,在中、重度鹽漬化及干旱環境中種植,植株生長受到嚴重限制。Song等[24]率先在Williams 82大豆(Glycinemax)基因組中鑒定到176個WRKY類轉錄因子,RNA-Seq技術篩選出12個受鹽脅迫誘導上調表達的基因。Yu等[25]在大豆基因組中鑒定到188個WRKY類轉錄因子,應用RNA-Seq技術篩選出66個受鹽脅迫影響的基因,經qRT-PCR驗證,獲得35個受鹽脅迫誘導下調表達的基因和19個受鹽脅迫誘導上調表達的基因。將栽培大豆WRKY類鹽脅迫響應基因轉入百脈根,在獲得耐鹽轉基因百脈根新材料的同時,在百脈根中揭示該基因的抗鹽機理,此類研究目前未見報道。
本研究克隆得到1個受鹽脅迫誘導上調表達的大豆WRKY類轉錄因子基因GmWRKY6。構建由強組成型啟動子CaMV35S調控的植物過表達載體,利用根癌農桿菌介導的子葉節轉化法對百脈根進行遺傳轉化,經PCR、RT-PCR檢測獲得過表達GmWRKY6基因的百脈根陽性植株。對其中的3個轉基因株系進行抗鹽試驗,初步確定GmWRKY6基因在鹽脅迫應答中的功能,為進一步闡明GmWRKY6基因在鹽脅迫應答調控中的作用機制奠定了重要的理論基礎。本研究不僅為大豆耐鹽基因工程改良提供了新的候選基因,而且對于培育耐鹽百脈根新品種,增強其在鹽堿地區的生長能力,充分開發利用鹽堿化土地資源具有重要意義。
1 材料與方法
1.1 實驗材料及處理
大豆測序品種Williams 82種子由中國科學院東北地理生態研究所大豆研究室提供;百脈根MG-20種子、改造的植物表達載體p1302G(攜帶有GUS標簽)由華中農業大學農業微生物國家重點實驗室根瘤菌分子研究室提供。2016年7月,大豆種子經表面消毒后,置于潤濕濾紙上待萌發,溫度28 ℃,18 h光照,6 h黑暗。4~5 d后移入1/2 Hoagland培養液中,培養條件不變。待幼苗長出第1片復葉時開始鹽脅迫處理。將實驗組幼苗移入含200 mmol·L-1NaCl的1/2 Hoagland培養液中,分別取鹽處理后0、2、6和24 h大豆根尖組織,對照組幼苗不經鹽脅迫處理,與實驗組在相同時間取樣,樣品液氮速凍后置于-80 ℃冰箱儲存備用。處理組和對照組每組8棵苗,并設置3個重復。
1.2 GmWRKY6基因的克隆和植物表達載體的構建
抽提大豆根組織總RNA,按照反轉錄試劑盒(TIANGEN公司)說明獲得cDNA 第一鏈。根據https://www.soybase.org/search/網站公布的大豆Glyma.15g110300.1基因序列設計引物F-OX(5′-GGAATTCATGGACAAAGGATGG-3′)和R-OX(5′-TCCCCCGGGTCAGTTTC CTGAA AAGC-3′)。下劃線堿基分別為上游和下游酶切位點EcoRⅠ和SmaⅠ。PCR擴增目的片段,連接T載體測序驗證后插入改造過的植物過表達載體p1302G(Gus基因作為篩選標記基因)中,構建重組質粒p1302G-GmWRKY6。采用凍融法將構建成功的重組質粒轉化到根癌農桿菌EHA105中備用。
1.3 GmWRKY6基因的表達分析
從已公布的大豆轉錄組數據(https://www.soybase.org/search/)中獲取GmWRKY6基因的時空表達數據,包括不同組織或者同一組織在不同時期的表達情況,利用Excel表繪圖分析GmWRKY6基因的時空表達特征。此外,利用Primer 5.0軟件設計1對GmWRKY6特異性引物F-rt(5′-ATGGACAAAGGATGG-3′)和R-rt(5′-AACTTCACCGACAAC-3′),分別提取1.1中鹽脅迫處理和對照樣品總RNA,反轉錄獲得cDNA第一鏈后,利用熒光定量PCR檢測GmWRKY6基因表達量。以大豆肌動蛋白11(ACT11)為內參基因,引物為F-ACT11(5′-ATTTTGACTGAGCGTGGTTATTCC-3′)和R-ACT11(5′-GCTGGTCCTGGCTGTCTCC-3′)。根據相對定量法(2-ΔΔCT)公式計算結果。設置3次獨立生物學重復和3次技術重復,數據取3次重復的平均值。
1.4 轉GmWRKY6基因百脈根的獲得
參照《LotusjaponicusHandbook》[26]所述方法,用1.2中制備好的含有GmWRKY6基因的農桿菌菌懸液侵染百脈根子葉節,侵染后的外植體經共培養-再生-篩選-芽誘導-芽生長-芽伸長-根誘導-根伸長等步驟后,獲得15株抗性苗,將其移栽到盆缽中保鮮膜覆蓋煉苗4~5 d,18 h光照,6 h黑暗,22 ℃培養,每天澆灌1/8 Hoagland 營養液1次。
提取15株抗性苗葉片總DNA,PCR擴增Gus標簽基因,引物為F-Gus(5′-GTCGCGCAAGACTGTAACCA-3′)和R-Gus(5′-CGGCGAAATTCCATACCTG-3′)。經PCR檢測的陽性苗,進一步進行RT-PCR檢測。提取葉片總RNA,反轉錄后用1.2中引物擴增GmWRKY6基因。以百脈根GPDH為內參基因,引物為F-GPDH(5′-GTGGTGCCAAGAAGGTTGTTAT-3′)和R-GPDH(5′-CTGGGAATGATGTTGAAGGAAG-3′)。RT-PCR檢測為陽性的7株百脈根植株,在光照培養箱中以相同條件繼續培養,待開花結莢后,收集T0代種子并作好記錄。
1.5 轉基因百脈根的鹽脅迫處理
將1.4中收集的T0代轉基因百脈根種子與野生型種子表面消毒,移入潤濕濾紙上待萌發。1周后將幼苗種入含基質盆缽中,每盆5顆苗,18 h光照,6 h黑暗,22 ℃培養,每天澆灌1/8 Hoagland 營養液1次。3周后選取生長狀態良好,長勢相似的植株,平均分成3組,每組6盆共30株苗進行鹽脅迫處理,分別用含有0、100和200 mmol·L-1NaCl的1/8 Hoagland營養液每天澆灌1次,2周后分析轉基因和野生型百脈根的抗鹽表型,測量株高,根長并進行相關生理指標測定。
1.6 抗鹽相關生理指標的檢測
采用水合茚三酮法測定脯氨酸含量[14];采用硫代巴比妥酸(TBA)比色法測定丙二醛(MDA)含量[22];采用80%丙酮抽提比色法測定葉片中葉綠素含量[22];利用電導率儀檢測葉片的相對電導率,測定相對質膜透性[22]。利用火焰分光光度計測定葉片中Na+和K+含量[14]。所有指標測定均設置3次重復,取平均值。
1.7 統計分析
用SPSS 13.0軟件檢驗差異顯著性。
2 結果與分析

圖1 GmWRKY6基因的PCR擴增(A)和重組質粒p1302G-GmWRKY6的酶切鑒定(B) Fig.1 PCR amplification of GmWRKY6 gene (A) and enzyme digestion of the recombinant plasmid p1302G-GmWRKY6 (B) M: DL2000 DNA marker; 1: GmWRKY6目的片段 Target fragment of GmWRKY6 gene; 2: 重組質粒p1302G-GmWRKY6的EcoRⅠ和SmaⅠ雙酶切 Recombinant plasmid p1302G-GmWRKY6 digested with EcoRⅠ and SmaⅠ.
2.1 GmWRKY6基因的克隆及重組質粒的構建
根據GmWRKY6基因的已知序列設計引物,以大豆根組織cDNA為模板,通過PCR 擴增得到1條長度為1674 bp 的條帶,與預期大小一致(圖1A)。將目的片段插入改造過的植物表達載體p1302G中,重組質粒經EcoRⅠ和SmaⅠ雙酶切后獲得大小正確的載體片段和目的片段(圖1B),將質粒送交公司測序。利用凍融法將測序結果正確的質粒轉入根癌農桿菌EHA105中保種備用。
2.2 GmWRKY6蛋白的同源比對及進化樹分析
利用NCBI中的Blastp程序以及DNAMAN軟件,對大豆GmWRKY6氨基酸序列與其他植物中同源蛋白進行多序列比對發現(圖2),GmWRKY6基因編碼的氨基酸序列與野生大豆GsWRKY6同源性最高,將該基因命名為GmWRKY6。對GmWRKY6蛋白保守結構域進行分析可知,該蛋白包含1個WRKY結構域,包括N-端高度保守的WRKYGQK七肽及C-端的C2H2鋅指結構(Cx5Cx23HxH)。根據WRKY結構域的個數、鋅指結構的類型及WRKY結構域中某些氨基酸差異,將GmWRKY6蛋白歸為第Ⅱ大類b小類WRKY轉錄因子。上述10種WRKY類同源蛋白的進化樹分析結果表明(圖3),大豆GmWRKY6與野生大豆GsWRKY6處在同一進化分支上,在進化過程中二者親緣關系較近。
2.3 GmWRKY6基因的組織表達分析
從已公布的大豆轉錄組數據中獲取GmWRKY6基因在不同組織及不同時期的表達情況,如圖4所示,GmWRKY6基因在根中表達量最高,其次是花、開花后14和10 d莢殼、根瘤、幼葉以及1 cm豆莢,在開花后10、14、21、25、28、35、42 d種子中表達量很低甚至檢測不到GmWRKY6基因的表達。說明GmWRKY6基因的表達在大豆中具有組織特異性,其行使功能的主要部位可能在根中。

圖2 GmWRKY6與其他植物WRKY蛋白的多序列比對Fig.2 Multiple sequence alignment of GmWRKY6 and other WRKY proteins

續圖2 GmWRKY6與其他植物WRKY蛋白的多序列比對Continued Fig.2 Multiple sequence alignment of GmWRKY6 and other WRKY proteins 下劃線部分表示WRKY結構域 The underlined region indicates WRKY domain;* 部分表示WRKYGQK氨基酸序列 * represents WRKYGQK amino acid sequence;↓ 表示C2H2鋅指結構的C 和H 殘基 ↓ means C and H residues in C2H2 zinc-finger motif.
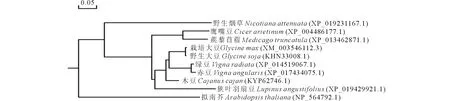
圖3 大豆GmWRKY6與其他植物WRKY轉錄因子進化樹分析Fig.3 Phylogenetic tree of GmWRKY6 with other plants’ WRKY transcription factors
2.4 GmWRKY6基因的鹽脅迫表達分析

圖4 大豆GmWRKY6基因在不同組織中的表達分析Fig.4 Relative expression levels of GmWRKY6 in different tissues 1: 幼葉 Young leaf;2: 花 Flower; 3:豆莢 Pod (1 cm); 4: 莢殼 Pod shell(開花后10 d 10 days after flower); 5: 莢殼 Pod shell(開花后14 d 14 days after flower); 6: 種子 Seed(開花后10 d 10 days after flower); 7: 種子 Seed(開花后14 d 14 days after flower); 8: 種子 Seed(開花后21 d 21 days after flower); 9: 種子 Seed(開花后25 d 25 days after flower); 10: 種子 Seed(開花后28 d 28 days after flower); 11: 種子 Seed(開花后35 d 35 days after flower); 12: 種子 Seed(開花后42 d 42 days after flower); 13: 根 Root; 14: 根瘤 Nodule.

圖5 大豆GmWRKY6基因在鹽脅迫下的表達分析Fig.5 Expression analysis of GmWRKY6 treated with 200 mmol·L-1 NaCl *:差異顯著(P<0.05) Significant difference (P<0.05); **:差異極顯著(P<0.01) Extremely significant difference (P<0.01); 下同 The same below.
通過Real-time PCR檢測GmWRKY6基因在無鹽脅迫下以及鹽處理后0、2、6和24 h的表達情況,如圖5所示,200 mmol·L-1NaCl處理2 h時,GmWRKY6基因的表達量迅速上升,隨處理時間的延長表達量持續升高,至6 h 時達到最大值,為脅迫前的22倍,24 h時略有下降,為脅迫前的17倍,而對照組中GmWRKY6基因在4個不同時間的表達量沒有顯著性差異。說明GmWRKY6基因確實是一個受鹽誘導上調表達的WRKY類轉錄因子。
2.5 轉GmWRKY6基因百脈根的分子生物學檢測
應用根癌農桿菌介導的子葉節轉化法將植物表達載體p1302G-GmWRKY6導入百脈根,經Kana篩選得到15株抗性植株,應用Gus標簽基因的特異引物對抗性植株進行PCR 鑒定,在#3~#10這8個轉基因植株中檢測到較明顯的目的條帶,#11、#13和#16植株中出現較弱的條帶,只有#12、#14、#15和#17這4個植株中未檢測到目的條帶,說明抗性植株的PCR 陽性率較高,達到了73%,如圖6A所示,共獲得PCR 陽性植株11 株。利用半定量RT-PCR方法檢測部分PCR陽性植株中GmWRKY6基因的表達,在#3~#9這7個轉基因植株中均能檢測到GmWRKY6基因的轉錄表達,而在野生型對照中未檢測到該基因的轉錄信號(圖6B)。
2.6 轉GmWRKY6基因百脈根的耐鹽表型分析
選取RT-PCR陽性植株#4、#6和#7這3個轉基因株系進行鹽脅迫處理。將轉基因和野生型株系分別用0、100 和200 mmol·L-1NaCl處理2周,結果發現在正常條件下,轉基因與野生型株系的生長狀態并無明顯差異,而在100 mmol·L-1NaCl脅迫下,3個轉基因株系的生長狀態明顯優于對照,當鹽濃度增加到200 mmol·L-1時,轉基因株系仍能正常生長,而野生型株系出現明顯的干枯萎蔫甚至死亡現象(圖7),說明轉基因株系耐鹽性明顯好于野生型對照。隨后對所有株系的株高與根長進行了測量,隨著鹽濃度的增大,轉基因與對照植株的株高與根長都呈下降趨勢,但3個轉基因株系的下降幅度均明顯小于對照(圖8)。如在200 mmol·L-1NaCl處理條件下,轉基因株系#6的株高是對照株系的1.63倍,轉基因株系#4的根長是對照株系的1.54倍。以上證據表明轉GmWRKY6基因百脈根耐鹽能力較強。

圖6 轉基因百脈根的分子生物學檢測Fig.6 Molecular detection of transgenic L. japonicus A: PCR檢測轉基因百脈根中GUS基因的表達 Detection of GUS gene expression in transgenic L. japonicus by PCR; B: RT-PCR檢測轉基因百脈根中GmWRKY6基因的表達 Detection of GmWRKY6 gene expression in transgenic L. japonicus by RT-PCR; M: DL2000 DNA marker; 1: 條帶大小為600 bp的質粒陽性對照 Plasmid positive control with 600 bp size; 2: 野生型對照 Wild type control; #3~#17: 轉基因株系 Transgenic plants.

圖7 轉基因百脈根抗鹽表型分析Fig.7 Salt stress phenotype analysis of transgenic L. japonicus Wt:野生型株系 Wild type plant. 下同 The same below.
2.7 鹽脅迫相關生理指標的檢測
脯氨酸作為一種滲透性保護物質,參與調節細胞質內滲透勢。從圖9A可知,在無鹽脅迫時,脯氨酸在轉基因和野生型株系中的含量基本保持一致。經鹽處理后二者的脯氨酸含量均呈上升趨勢,但轉基因株系的上升幅度明顯高于對照,在200 mmol·L-1NaCl處理條件下差異極顯著(P<0.01)。說明鹽脅迫下,轉基因株系中的脯氨酸大量積累,保護細胞避免脫水,從而降低膜損傷。
丙二醛是膜脂過氧化的主要產物之一,它的含量反映了鹽脅迫對膜氧化損傷程度的大小。圖9B 顯示,隨著鹽濃度的升高,轉基因株系和對照的丙二醛含量均呈上升趨勢,但3個轉基因株系的增加幅度顯著低于對照(P<0.01),說明轉基因百脈根降低了高鹽脅迫對膜氧化的損傷程度,耐鹽能力得到了顯著提高。

圖8 鹽脅迫對轉基因百脈根株高(A)和根長(B)的影響Fig.8 Changes of shoot height (A) and root length (B) of transgenic L. japonicus under salt stress
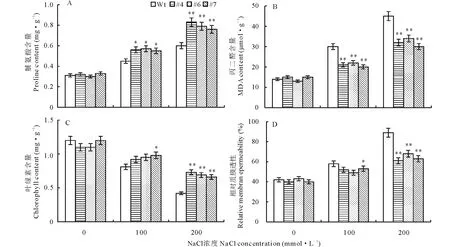
圖9 鹽脅迫下轉基因百脈根的生理指標測定Fig.9 Analysis on physiological characteristics of transgenic L. japonicus under salt stress
從圖9C可知,在無鹽脅迫時,葉綠素在轉基因和野生型株系中的含量未見明顯差異。經鹽處理后,3個轉基因株系和野生型對照的葉綠素含量均迅速下降,但轉基因株系的下降幅度低于野生型對照,在200 mmol·L-1NaCl處理條件下,野生型對照的葉綠素含量顯著低于轉基因株系(P<0.01)。顯然在鹽脅迫條件下轉GmWRKY6基因百脈根株系的葉綠體受傷害程度較小,植株具有更強的光合能力,從而能夠保持更好的生長狀態。
圖9D表明,隨著鹽濃度的增加,轉基因和野生型株系的相對質膜透性也隨之增加,但轉基因植株的增加幅度低于野生型對照,在200 mmol·L-1NaCl處理條件下,野生型對照的相對質膜透性顯著高于轉基因株系(P<0.01)。說明在鹽脅迫條件下,轉GmWRKY6基因株系細胞膜所受的損害較輕,質膜完整性較好,因此細胞膜具有較強的生理活性,植株表現出較高的耐鹽性。
2.8 鹽脅迫下轉基因百脈根Na+和K+含量測定
通過測定轉基因和對照植株葉片中Na+和K+含量發現(圖10),非脅迫條件下,轉基因百脈根葉片內Na+和K+含量與野生型之間沒有明顯差異。隨著鹽濃度的增加,3個轉基因株系和野生型對照葉片中Na+含量均隨之升高,與之相反的是,K+含量均隨之降低。在同一鹽濃度處理下,轉基因株系葉片中Na+含量顯著低于對照株系,K+含量顯著高于對照株系。在100 mmol·L-1NaCl處理條件下二者差異顯著(P<0.05),在200 mmol·L-1NaCl處理條件下二者差異極顯著(P<0.01)。說明轉基因株系在高鹽脅迫下通過維持較低的Na+水平和較高的K+水平,從而調節細胞內的離子穩態平衡,增強植株的耐鹽性。

圖10 鹽脅迫下轉基因百脈根葉片中Na+(A)和K+(B)含量測定Fig.10 Analysis on Na+ (A) and K+ (B) content in transgenic L. japonicus leaves under salt stress
3 討論
WRKY轉錄因子最早由Ishiguro等[27]從甘薯中分離得到,隨后不同植物中的WRKY轉錄因子相繼被鑒定出來[28-30]。WRKY蛋白都包含1個或者兩個WRKY結構域,N-端含有1個保守的七肽WRKYGQK,C-端含有1個鋅指結構(Cx4-7Cx22-23HxH/C)。WRKY轉錄因子按照WRKY結構域個數和鋅指結構類型的不同,可分為3類:第Ⅰ類成員WRKY結構域個數為2個,鋅指結構類型為C2H2,第Ⅱ、Ⅲ類成員WRKY結構域個數為1個,第Ⅱ類成員鋅指結構類型為C2H2,第Ⅲ類成員鋅指結構類型為C2HC。根據親緣關系及WRKY結構域中個別氨基酸的差異,可將第Ⅱ類WRKY蛋白細分為a、b、c、d、e 5個小類[31]。本研究克隆得到1個鹽誘導WRKY類轉錄因子基因GmWRKY6(圖1和圖5),同源蛋白比對和進化樹分析發現(圖2和圖3),該基因與野生大豆GsWRKY6相似性最高,親緣關系最近,因此將其命名為GmWRKY6。GmWRKY6基因位于大豆第15號染色體上,含有1個WRKY結構域和C2H2(Cx5Cx23HxH)型鋅指結構,屬于第Ⅱ類b小類WRKY轉錄因子。
大量證據表明,大豆WRKY類轉錄因子在植物抗逆反應中發揮重要作用。如GmWRKY21是一個冷脅迫響應基因。GmWRKY54可能通過調控DREB2A和STZ/Zat10抵抗鹽和干旱脅迫。過表達GmWRKY13轉基因擬南芥表現出對鹽和甘露醇脅迫的敏感性增加,對脫落酸的敏感性下降。過表達GmWRKY28提高了擬南芥的耐鹽性,轉基因擬南芥萌發率和根長明顯增加[32]。過表達GmWRKY28-like顯著增強了擬南芥的耐鹽性[33]。GmWRKY35基因受干旱脅迫誘導表達,在干旱脅迫下,與對照相比,轉基因煙草長勢較好,POD和SOD活性較高,MDA含量和電解質滲漏率較低,說明GmWRKY35基因的過表達增強了煙草的抗旱能力[34]。目前,利用轉基因技術向苜蓿中導入具有優良性狀的目的基因來提升轉基因苜蓿的耐鹽堿性狀,已成為改良農業和畜牧業生產的重要途徑。大豆和百脈根的親緣關系更近,遺傳物質更容易交流。因此,采用生物技術手段,將栽培大豆WRKY類鹽脅迫響應基因轉入百脈根,利用大豆的優良抗逆性狀培育優質高抗百脈根新品種,增強其在鹽堿地區的生長能力,對提高百脈根牧草的產量,充分開發利用鹽堿化土地資源具有重要意義。
本研究利用農桿菌介導的子葉節轉化法獲得了過表達GmWRKY6基因的轉基因百脈根(圖6)。在鹽脅迫下,與野生型對照相比,轉基因百脈根長勢較好(圖7),株高較高,根長較長(圖8)。對相關生理指標的測定發現,轉基因株系的脯氨酸和葉綠素含量顯著高于對照植株,而丙二醛含量和相對質膜透性顯著低于對照植株(圖9)。脯氨酸含量的大量增加,提高了細胞的滲透調節能力,轉基因植株能更好地抵御鹽脅迫造成的損傷。葉綠素的增加有利于維持植株正常的光合作用,使轉基因植株保持良好的生長狀態。丙二醛含量和相對質膜透性的降低減輕了轉基因植株質膜的受損程度,使質膜維持正常的生理功能。此外,進一步對轉基因和對照植株葉片中Na+和K+含量檢測發現,轉基因植株在高鹽處理下葉片中Na+含量顯著低于對照植株,但K+含量顯著高于對照植株(圖10)。說明轉基因植株在高鹽脅迫下通過調節細胞內的離子穩態平衡,從而增強百脈根的耐鹽能力。以上結果說明GmWRKY6的超表達能夠提高百脈根的耐鹽能力。本研究為深入研究GmWRKY6基因的功能和培養百脈根耐鹽新品種奠定了基礎。
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
