2種溶解態銅對凡納濱對蝦Cu2+-ATPase及Ca2+-Mg2+-ATPase和 Cu-Zn-SOD的影響
2018-08-07 06:30:50彭士明史永富席寅峰施兆鴻
海洋漁業 2018年4期
關鍵詞:實驗
宋 泰,彭士明,史永富,席寅峰,黃 艇,施兆鴻
(1.中國水產科學研究院東海水產研究所,農業部東海與遠洋漁業資源開發利用重點實驗室,上海 200090;2.上海海洋大學水產與生命學院,上海 201306;3.國際銅業協會,上海 200000)
銅(Cu)是甲殼類賴以生存的金屬元素之一。甲殼類體內用于攜氧的主要蛋白是以銅為中心原子的血藍蛋白(CP)[1-2],可見銅元素直接關系到甲殼類的代謝功能。銅屬于重金屬元素,單質銅與Cu2+存在200~800 mV的氧化還原電位差,因此其代謝相關活性物質具有特異性,銅三磷酸腺苷酶(Cu2+-ATPase)作為 Cu2+的通道,其活力大小很大程度上影響著銅的代謝水平。Ca2+、Mg2+和Cu2+有著比較接近的電位差,當銅代謝過盛時,Cu2+能夠通過某種機制抑制鰓中的蛋白酶活力,如鑲嵌轉運亞基的基團或競爭性抑制離子通道,從而導致體內離子組分的失衡[3]。銅鋅超氧化物歧化酶(Cu-Zn-SOD)活力水平也受水體中銅水平的影響,Cu-Zn-SOD作為甲殼類清除自由基的主要酶蛋白,其活力的強弱關系到甲殼類健康。凡納濱對蝦(Litopenaeus vannamei)的Cu-Zn-SOD在對抗溶藻弧菌和白斑綜合癥病毒脅迫中起著重要的作用[4]。因此研究水體中銅對甲殼類體內 Cu2+-ATPase、Ca2+-Mg2+-ATPase和 Cu-Zn-SOD的活性大小的影響,對了解銅代謝的水平有著理論意義。
甲殼類對重金屬的吸收主要有兩種途徑:一是通過攝食把水體中或餌料中的重金屬經消化道進入體內,最終在肝胰臟中富集,另一種途徑是經過鰓不斷吸收溶解在水中的重金屬離子,經體液循環最終積累在細胞中[5]。因此,研究不同溶解態的銅對甲殼類的作用有著積極的生物學意義。銅合金在海水中可以通過電離、氧化還原反應來釋放微量銅離子(包含Cu+和Cu2+),而海水會通過絡合反應使水體離子銅絡合,總銅保持在一個相對穩定的范圍內[6]。銅合金板和Cu2+均能起到使細菌失活的作用,有報道認為,過量的Cu2+可破壞細菌內蛋白[7],銅合金板本身存在的電位差,使其接觸的細菌失活[8],可以在不溶于水體的狀態下降低養殖水體有害細菌含量。本研究以凡納濱對蝦為實驗對象,采用目前在養殖過程中作為抑制原生動物或殺菌的LC6911型銅板為釋放絡合銅源的材料,并以五水硫酸銅(CuSO4·5H2O)作為Cu2+的原料進行對比,研究不同時長、不同溶解態銅以及不同添加量銅對蝦體內不同組織中 Cu2+-ATPase、Ca2+-Mg2+-ATPase和Cu-Zn-SOD的影響。旨在探討在水體中使用銅合金(LC6911型銅板)作為絡合銅源提升凡納濱對蝦代謝功能的可行性,為今后的生產和研究提供參考數據。
1 材料與方法
1.1 實驗材料
實驗在中國水產科學研究院東海水產研究所福鼎研究中心內進行。凡納濱對蝦購自福建當地養殖場,體長(9.0±0.9)cm,體質量(11.5±2.3)g。實驗海水經自然沉淀、砂濾、蛋白質分離器處理后使用,處理后的海水經檢測離子銅含量低于1×10-6mg·L-1,總銅含量(3.1×10-5±1.21×10-6)mg·L-1,鹽度26±0.8。
實驗用飼料為正大牌對蝦飼料,為避免飼料中銅干擾實驗,參照董曉慧等[9]的方法用EDTA浸泡處理后烘干儲存備用。實驗用LC6911型銅板由國際銅業協會提供,將厚度0.1 cm的銅板裁成50 cm×60 cm的小塊儲存備用。硫酸銅為CuSO4·5H2O,分析純(上海國藥集團化學試劑有限公司)。
1.2 實驗設計
1.2.1 LC6911銅合金在海水中銅釋放量
用直徑2 m、深1 m的玻璃鋼圓桶9個,每桶放置2 m3沙濾海水,實驗分設0.3 m2銅板和0.6 m2銅板平鋪桶底,即按水體計算分別為0.15 m2銅板·m-3和0.3 m2銅板·m-3,另設空白對照組。每組實驗設3平行,水溫28℃,鹽度26±0.8,不間斷充氣,不換水。分別在 0 h、1 h、2 h、4 h、8 h、12 h、24 h、36 h、48 h、60 h、72 h、96 h取各平行水樣,用離子計(雷磁 PXSJ—216F型離子計)測定水體Cu2+含量以及水體總銅含量。水體總銅含量測定方法參照李登新[10]:將水樣降溫至25℃,用鹽酸滴定至水體pH 5.2后再用離子計測 Cu2+濃度(注:25℃Ksp[Cu(OH)2]=2.2×10-20)。
1.2.2 養殖實驗設計
根據1.2.1的結果,空白對照組測定的Cu2+量值穩定在(3.18×10-5±6.88×10-7)mg·L-1,在2 m3的海水中每0.3 m2銅板48 h內的釋放總銅量為(0.245±3.70)×10-4mg·L-1。另參照中華人民共和國農業部頒發的海水水質標準[10]和劉存岐等[11]對中國明對蝦 (Penaeus chinensis)養殖實驗中海水添加銅濃度梯度,本實驗設1個對照組和4個試驗組即2個用銅板作為絡合銅試驗組和2個添加CuSO4·5H2O的Cu2+試驗組,分別為:A組(空白對照),B組(0.15 m2銅板·m-3水體),C組(0.3 m2銅板·m-3水體),D組(0.25 mg Cu2+·L-1水體),E組(0.50 mg Cu2+·L-1水體),每組均設3個平行。實驗用容器與1.2.1所述相同,每個重復分別放置40尾蝦。實驗開始前暫養1周。暫養和實驗期間環境條件均為水溫(28±1.1)℃、鹽度26±0.8、不間斷充氣、每隔48h換水10%,清除殘餌糞便,換水前檢測各平行水體內總銅含量與離子Cu2+含量。由于Cu2+會緩慢絡合,每次換水時向D、E組中補充因換水流失濃度的CuSO4·5H2O,補充流失的Cu2+。投餌時均使用蝦料筐,避免飼料接觸容器底部的銅板。投料量約為蝦體質量的6%。
1.3 采樣方法
飼養期間,在 0、2、4、8、16、32、56 d取樣,每重復實驗取3尾,用1 mL的注射針自對蝦胸下動脈抽取血淋巴,再解剖分取鰓、肝胰臟后半部分和肌肉,取得樣品置離心管內,-80℃冷凍保存待測。
1.4 蝦體銅含量以及酶活力的測定
檢測不同組織中的鈣鎂三磷酸腺苷酶(Ca2+- Mg2+-ATPase)、銅 三 磷 酸 腺 苷 酶 (Cu2+-ATPase)和銅鋅超氧化物歧化酶(Cu-Zn-SOD)。通過單位時間內ATP酶分解的無機磷的量來測定ATP酶活力;1 mg組織蛋白在1 mL反應液中SOD抑制率達50%時所對應的SOD量為一個活力單位(U)。實驗所涉及酶及蛋白類指標使用南京建成生物科技有限公司的試劑盒進行測定,具體方法參見說明書。實驗所測蝦體內銅含量方法為原子火焰吸收法,樣品前處理采用硝酸-高氯酸濕式消解。
1.5 數據處理
除水體銅含量實驗的數據外,其它實驗數據處理均進行統計學分析。分析使用Spss Statistics 19軟件,使用單因素ANOVA對組間進行差異顯著性分析;Ducan法進行方差齊次性檢驗。P<0.05表示差異具有顯著性。
2 結果與分析
2.1 LC6911銅合金在海水中的銅釋放量和各組的銅含量
在無養殖生物條件下,24 h內銅板單位水體的總銅釋放量和時間呈正比(表1):24~96 h,0.15 m2銅板·m-3和 0.3 m2銅板·m-3釋放Cu2+量分別穩定在2.18×10-1~1.75×10-1mg·L-1和3.27×10-1~6.47×10-1mg·L-1。實驗期內未放置銅板的空白對照組的Cu2+量值穩定在(3.18×10-5±6.88×10-7)mg·L-1。
養殖實驗期間,空白對照A組總銅含量低于1.08×10-6mg·L-1,B、C組 Cu2+含量低于1.42×10-5mg·L-1,B組總銅含量(2.48×10-1±3.34×10-2)mg·L-1,C組總銅含量(4.86×10-1±5.76×10-2)mg·L-1。D組 Cu2+含量區間調控在2.1×10-1~2.5×10-1mg·L-1,E組Cu2+含量區間調控在4.2×10-1~5.0×10-1mg·L-1。實驗期間除采樣外,各試驗組、對照組凡納濱對蝦存活率均為100%,無死亡現象。
2.2 2種溶解態銅對凡納濱對蝦肌肉銅含量影響
凡納濱對蝦肌肉中銅含量在實驗周期內變化規律見圖1。對照組中肌肉銅含量在實驗過程各時間點間變化不顯著(P>0.05);2組絡合態銅的試驗組在2 d起至實驗結束始終顯著高于對照組(P<0.05);2組Cu2+試驗組在實驗初期也同樣在2 d時就與對照組呈顯著性差異(P<0.05),隨實驗時間的延長,與對照組之間的差異逐漸減小,高Cu2+試驗組在32 d后已低于對照組,且在實驗結束時(56 d)低于對照組。低Cu2+試驗組同樣也隨實驗時間延長而含量下降。
2.3 2種溶解態銅對凡納濱對蝦不同組織中Cu2+-ATPase活力的影響
實驗期內,各試驗組凡納濱對蝦鰓組織中的Cu2+-ATPase活力均顯著高于對照組(P<0.05)。且呈不斷上升的趨勢,實驗結束時0.3 m2銅板·m-3試驗組已經高出對照組的8倍,0.15 m2銅板·m-3試驗組也高于對照組4.2倍;Cu2+試驗組呈現與絡合銅試驗組相同的變化趨勢,高Cu2+試驗組和低Cu2+試驗組分別高于對照組6.4倍和4.9倍。2種溶解態銅不同濃度之間也呈顯著性差異(圖2)。結果表明,水體中不論何種溶解態的銅對鰓組織中Cu2+-ATPase活力均具有隨時間延長而酶活力增大的趨勢。

表1 0.15 m2·M-3和0.30 m2·m-3銅板試驗組隨時間釋放的總銅含量Tab.1 Total Cu release of 0.15 m2·m-3 and 0.30 m2·m-3 copper plate
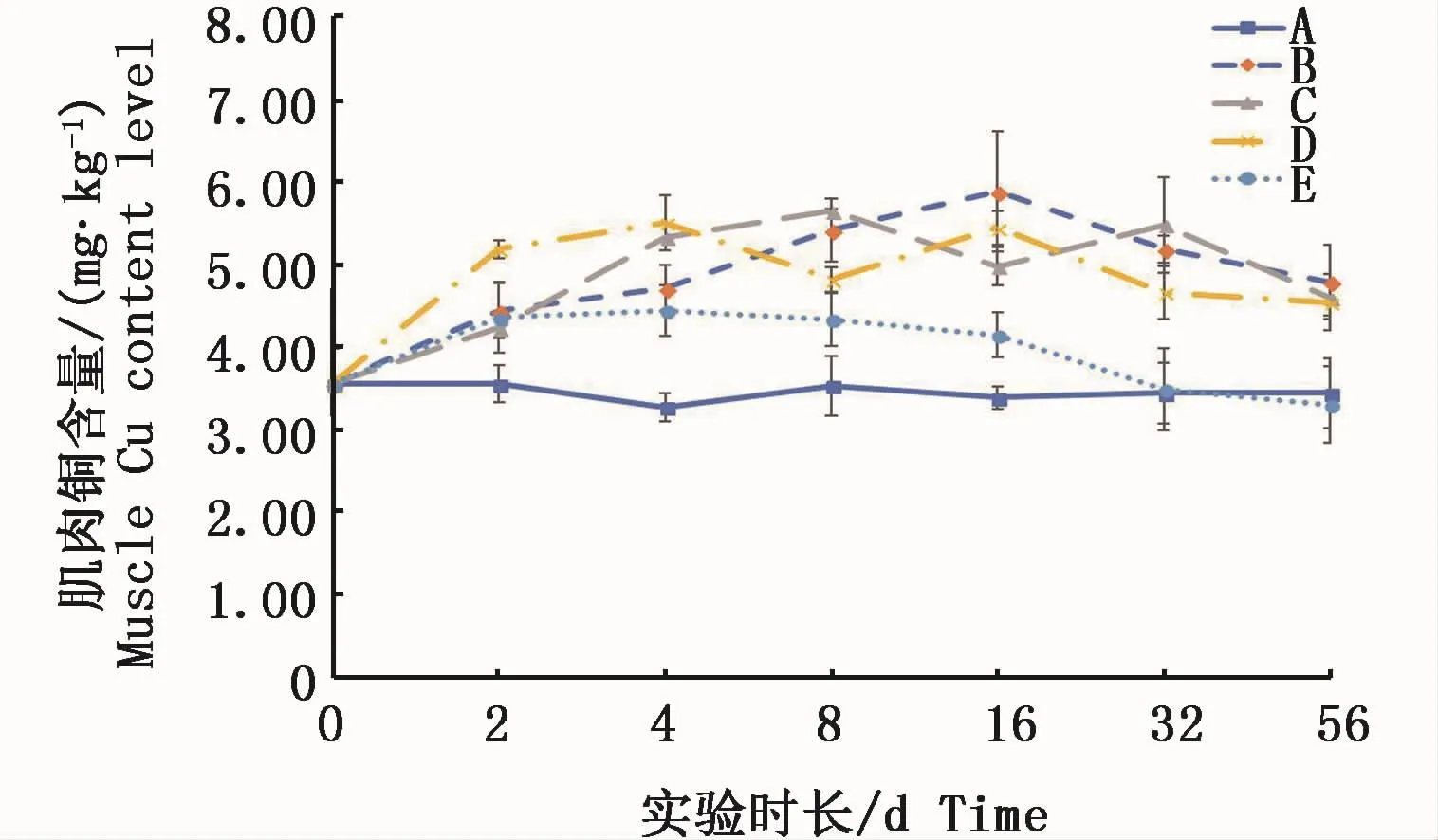
圖1 不同處理條件下凡納濱對蝦肌肉銅含量變化Fig.1 Changes of Cu content level in L.vannamei muscle tissue under different treatments注:A:空白對照組;B:0.15 m2銅板·m-3水體;C:0.3 m2銅板·m-3水體;D:0.25 mg Cu2+·L-1水體;E:0.5 mg Cu2+·L-1水體Note:A:Blank group;B:0.15 m2 copper plate·m-3;C:0.3 m2 copper plate·m-3;D:0.25 mg Cu2+·L-1;E:0.5 mg Cu2+·L-1
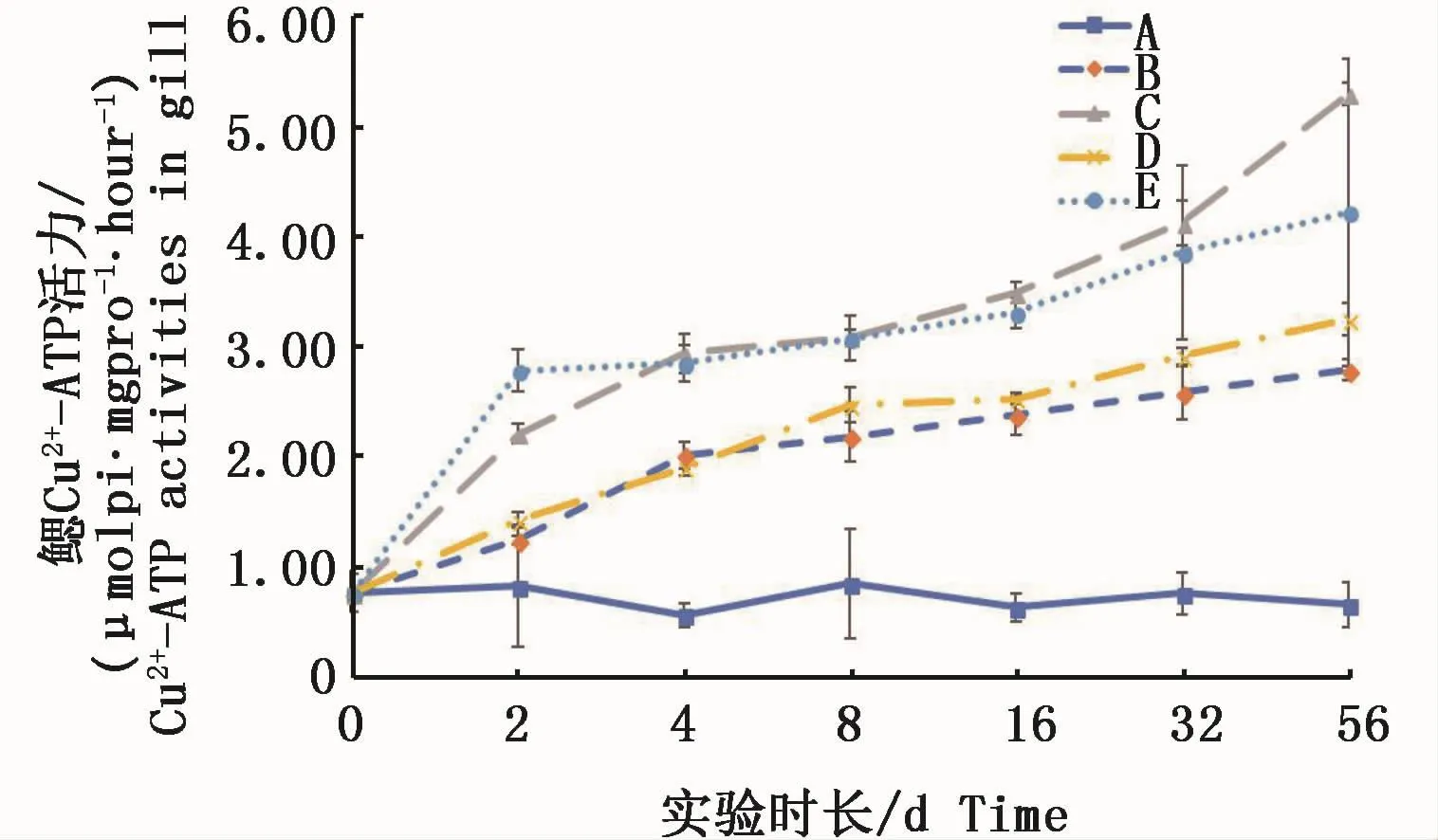
圖2 不同處理條件下凡納濱對蝦鰓Cu2+-ATPase活力變化Fig.2 Changes of Cu2+-ATPase activities in L.vannamei gills under different treatments注:A:空白對照組;B:0.15 m2銅板·m-3水體;C:0.3 m2銅板·m-3水體;D:0.25 mg Cu2+·L-1水體;E:0.5 mg Cu2+·L-1水體Note:A:Blank group;B:0.15 m2 copper plate·m-3;C:0.3 m2 copper plate·m-3;D:0.25 mg Cu2+·L-1;E:0.5 mg Cu2+·L-1
各試驗組肝胰腺中Cu2+-ATPase活力隨實驗時間延長而不斷增高,且與對照組呈顯著性差異(P<0.05)。不同溶解態的銅均是高濃度試驗組中的酶活力大于低濃度試驗組,Cu2+試驗組中低濃度組在4 d起與高濃度試驗組呈顯著性差異(P<0.05)。結果表明,凡納濱對蝦的肝胰腺中Cu2+-ATPase活力不僅與水體中銅的溶解態相關,還與濃度呈正相關(圖3)。
2.4 2種溶解態銅對凡納濱對蝦不同組織中Ca2+-Mg2+-ATPase活力的影響
鰓組織中的 Ca2+-Mg2+-ATPase活力在實驗前期(2 d)都呈上升趨勢,其中絡合銅試驗組和低Cu2+試驗組均與對照組呈顯著性差異(P<0.05)。隨后,0.15 m2銅板·m-3試驗組和低Cu2+試驗組先繼續上升后下降,而高濃度Cu2+試驗組和0.3 m2銅板·m-3試驗組則從2 d起不斷下降,各試驗組至實驗結束時均顯著低于對照組(P<0.05)(圖4)。結果表明,鰓組織中的 Ca2+-Mg2+-ATPase活力受2種溶解態銅濃度的影響均先升后降,8 d時除高濃度Cu2+試驗組顯著低于對照組外(P<0.05),其它各組均與對照組無顯著性差異(P>0.05),至實驗結束時各試驗組均顯著低于對照組(P<0.05);并且,鰓組織中Ca2+-Mg2+-ATPase活力與 Cu2+-ATPase活力一樣,在本實驗條件下均表現出銅水平對酶活力的影響大于銅的溶解態對酶活力的影響。
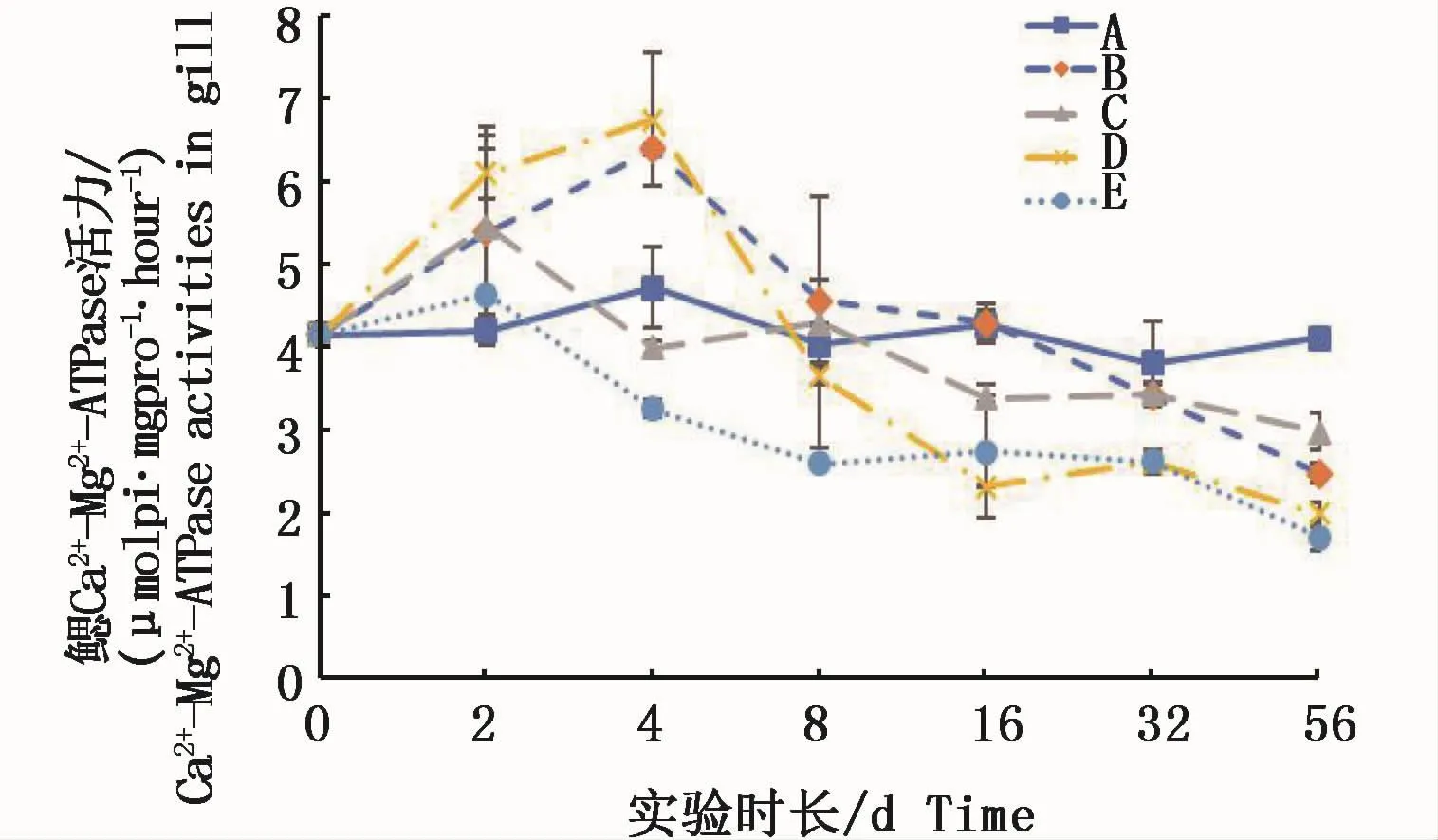
圖4 不同處理條件下凡納濱對蝦鰓Ca2+-Mg2+-ATPase活力變化Fig.4 Changes of Ca2+-Mg2+-ATPase activities in L.vannamei gills under different treatments注:A:空白對照組;B:0.15 m2銅板·m-3水體;C:0.3 m2銅板·m-3水體;D:0.25 mg Cu2+·L-1水體;E:0.5 mg Cu2+·L-1水體Note:A:Blank group;B:0.15 m2 copper plate·m-3;C:0.3 m2 copper plate·m-3;D:0.25 mg Cu2+·L-1;E:0.5 mg Cu2+·L-1
肝胰腺中 Ca2+-Mg2+-ATPase活力,不論何種溶解態均呈現出實驗前期上升、后期下降的態勢。絡合銅形態的試驗組在實驗4 d均顯著高于對照組(P<0.05),離子態的試驗組在4 d已經降至低于對照組,且高濃度離子態的試驗組與對照組呈顯著性差異(P<0.05)。實驗結束時56 d各試驗組均顯著低于對照組(P<0.05)。結果表明,水體中不論何種溶解態的銅,短時間會增加肝胰腺中Ca2+-Mg2+-ATPase的活力,而長時間則均對肝胰腺中Ca2+-Mg2+-ATPase活力產生抑制(圖5)。
2.5 2種溶解態銅對凡納濱對蝦不同組織中Cu-Zn-SOD活力的影響
凡納濱對蝦鰓組織的中Cu-Zn-SOD活力在2種溶解態銅的水體中實驗前期隨時間延長而升高,8 d均達到峰值,與對照組有顯著性差異(P<0.05)。16 d各試驗組均呈現回落態勢,但仍與對照組有顯著性差異(P<0.05);32 d各試驗組又呈現出上升并出現第二次峰值;實驗結束時(56 d)高絡合態和高離子態試驗組均驟降且顯著低于對照組(P<0.05),0.15 m2銅板·m-3和低Cu2+試驗組也分別下降至接近對照組,但與對照組無顯著性差異(P>0.05)(圖6)。
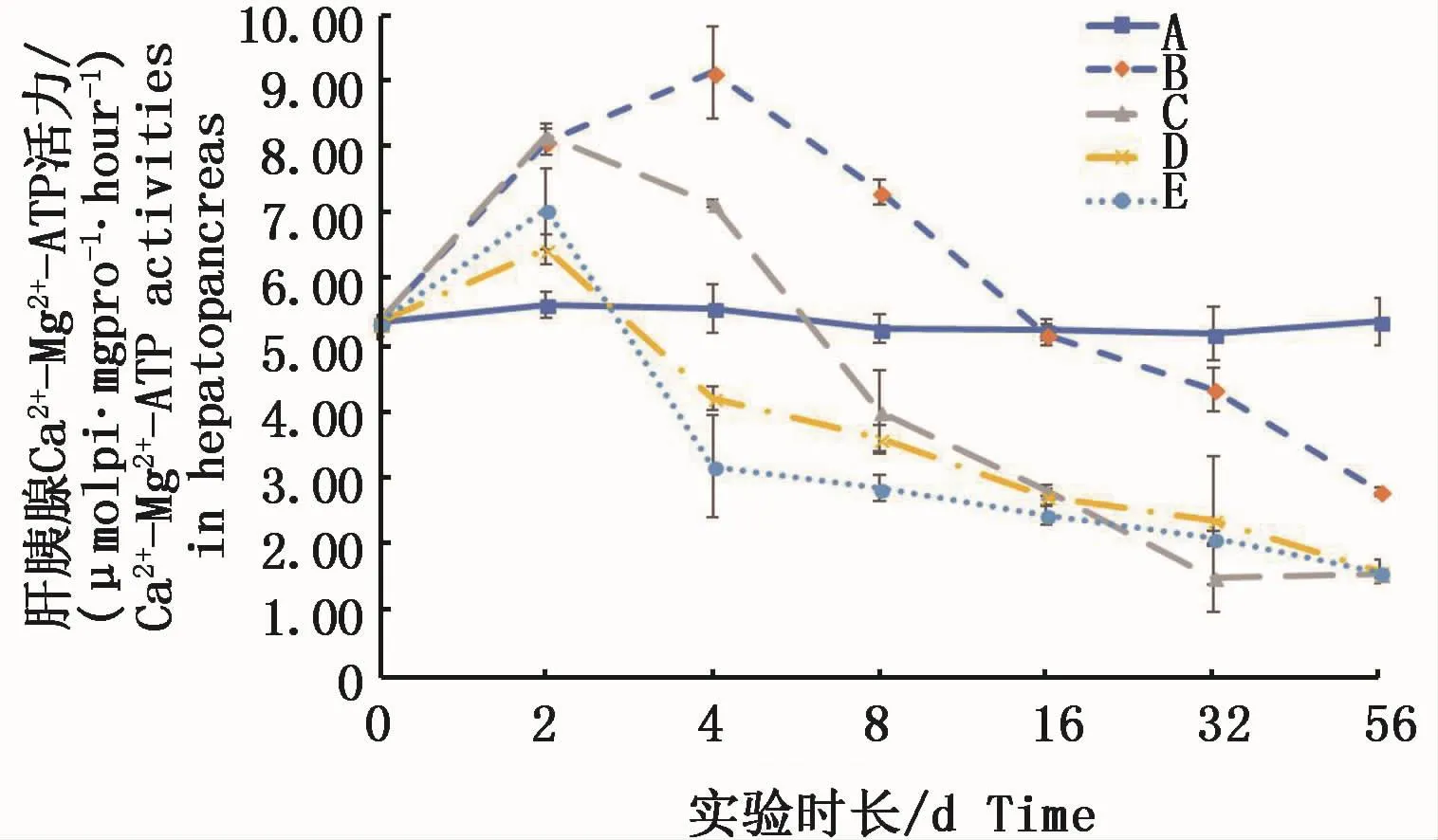
圖5 不同處理條件下凡納濱對蝦肝胰腺Ca2+-Mg2+-ATPase活力變化Fig.5 Changes of Ca2+-Mg2+-ATPase activities in L.vannamei hepatopancreas under different treatments注:A:空白對照組;B:0.15 m2銅板·m-3水體;C:0.3 m2銅板·m-3水體;D:0.25 mg Cu2+·L-1水體;E:0.5 mg Cu2+·L-1水體Note:A:Blank group;B:0.15 m2 copper plate·m-3;C:0.3 m2 copper plate·m-3;D:0.25 mg Cu2+·L-1;E:0.5 mg Cu2+·L-1
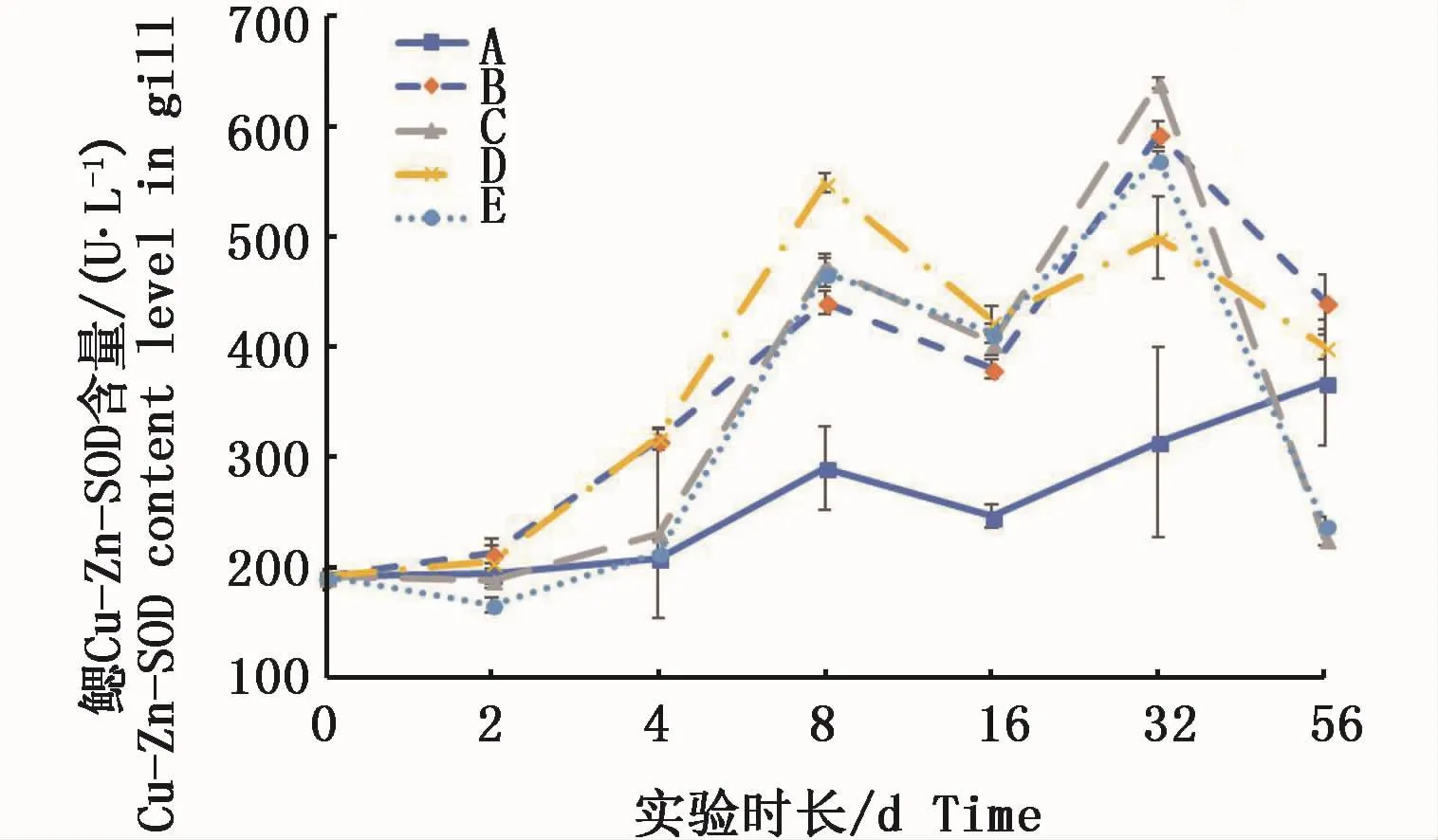
圖6 不同處理條件下凡納濱對蝦鰓Cu-Zn-SOD活力變化Fig.6 Changes of Cu-Zn-SOD activities in L.vannamei gills under different treatments注:A:空白對照組;B:0.15 m2銅板·m-3水體;C:0.3 m2銅板·m-3水體;D:0.25 mg Cu2+·L-1水體;E:0.5 mg Cu2+·L-1水體Note:A:Blank group;B:0.15 m2 copper plate·m-3;C:0.3 m2 copper plate·m-3;D:0.25 mg Cu2+·L-1;E:0.5 mg Cu2+·L-1
肝胰腺組織中的Cu-Zn-SOD活力在16 d前變化基本平穩,除2 d時絡合態試驗組均呈現出與對照組有差異外,其它各取樣點各試驗組均與對照組差異不顯著;32 d時各試驗組的Cu-Zn-SOD活力都驟升,達到對照組的2倍以上,56 d又同時回落,但仍與對照組之間有顯著性差異(P<0.05)(圖7)。
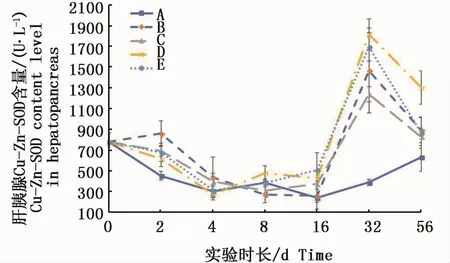
圖7 不同處理條件下凡納濱對蝦肝胰腺Cu-Zn-SOD活力變化Fig.7 Changes of Cu-Zn-SOD activities in L.vannamei hepatopancreas under different treatments注:A:空白對照組;B:0.15 m2銅板·m-3水體;C:0.3 m2銅板·m-3水體;D:0.25 mg Cu2+·L-1水體;E:0.5 mg Cu2+·L-1水體Note:A:Blank group;B:0.15 m2 copper plate·m-3;C:0.3 m2 copper plate·m-3;D:0.25 mg Cu2+·L-1;E:0.5 mg Cu2+·L-1
血淋巴中的Cu-Zn-SOD活力基本呈現出低濃度試驗組高于對照組、高濃度試驗組低于或接近對照組。各試驗組與對照組相似,均呈波浪起伏狀變化。
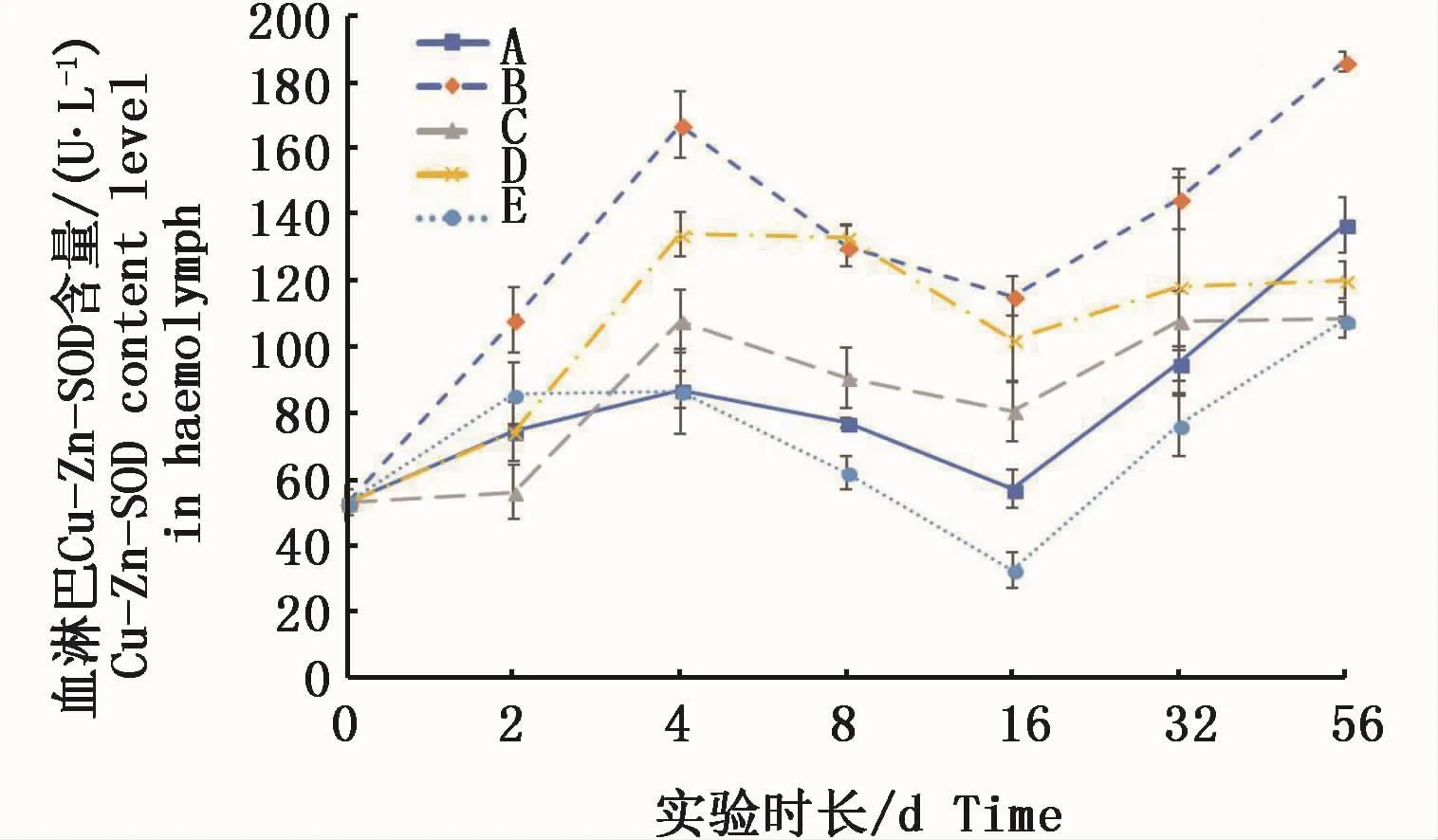
圖8 不同處理條件下凡納濱對蝦血淋巴Cu-Zn-SOD活力變化Fig.8 Changes of Cu-Zn-SOD activities in L.vannamei haemolymph under different treatments注:A:空白對照組;B:0.15 m2銅板·m-3水體;C:0.3 m2銅板·m-3水體;D:0.25 mg Cu2+·L-1水體;E:0.5 mg Cu2+·L-1水體Note:A:Blank group;B:0.15 m2 copper plate·m-3;C:0.3 m2 copper plate·m-3;D:0.25 mg Cu2+·L-1;E:0.5 mg Cu2+·L-1
3 討論
3.1 海水養殖環境銅源控制方法分析
物理腐蝕原理中認為,金屬銅表面與海水的摩擦系數為0.1左右,相較于其它材料,此摩擦系數能夠很大程度上降低物理磨損和物理腐蝕[12]。此外,電化學腐蝕原理認為,金屬銅在海水中可以形成一層Cu-Al-Mg共析氧化物的致密、均勻、與基體結合良好的氧化膜,該氧化膜能在銅的陽極以及陰極電解中起到重要保護作用[13]。本實驗數據表明,長期養殖環境中不間斷充氣的條件下,金屬銅釋放效率在48 h內與面積呈正比,通過適量的換水可保證水體總銅含量安全可控,可保證水體內銅含量水平準確。另外,銅合金具有較好耐腐蝕性能[14],本研究中56 d實驗后取出的銅板并未發現明顯的腐蝕痕跡。一定程度上說明LC6911銅合金不會因在海水內存在時間過長或養殖主體存在發生嚴重化學腐蝕現象。
3.2 水環境中銅水平對凡納濱對蝦肌肉中銅含量的影響
甲殼類在含銅的水體中,鰓是最先富集的組織之一。在自然狀況下,鰓中銅含量最高,其次是肝胰腺,而肌肉中的銅含量最低[15]。銅無論是通過攝入的餌料帶入、還是通過鰓進入生物體,在代謝過程中,當銅供大于求時就會暫存于肝胰腺中。蓄積于肝胰腺等組織中的銅以Cu2+形式通過血液循環到達肌肉以供代謝需要[16]。從本研究中可以看出(圖1),水體中的銅不論是何種溶解形態均會導致凡納濱對蝦肌肉中的銅含量升高,但隨實驗時間的增加,銅含量則逐漸減少。根據已有資料報道,細胞膜上有離子通道,是否存在除離子通道外的其它形態的通道,則需進一步研究。一般認為銅較難在水產動物體內富集。本實驗也證明了對蝦肌肉組織中不會隨時間或水體中的銅水平升高而大量富集,反而隨時間的延長銅含量會減少。本實驗條件下,是否鰓組織中銅含量最高以及肝胰腺中的銅含量是否富集,有待進一步的實驗證實。
3.3 2種溶解態銅對凡納濱對蝦銅代謝的影響
近幾年銅對水產動物的毒性、毒理、生長、代謝及細胞生理、生化作用的研究已有不少報道[17-20]。研究認為,以銅為中心原子的血藍蛋白對水產經濟動物的代謝和免疫至關重要。事實上,有證據表明一價銅在海水中大量是以絡合態的形式存在的[6],而一價銅和二價銅之間的氧化還原電位差可能對細胞膜上蛋白酶類有不同的作用。關于銅的離子態、化合態難溶物及絡合態的不同作用機理還值得深入研究。
有報道認為,銅缺乏可使機體內一些重要的抗氧化酶如Cu-Zn-SOD、血漿銅藍蛋白(CP)、谷胱甘肽過氧化物酶(GSH-Px)等的活性下降,導致自由基增加,而自由基也會攻擊細胞膜上的蛋白酶類,如ATP酶[22],ATP酶活力減弱使得細胞內Mg2+、K+離子濃度降低,會使細胞內DNA合成受阻,從而影響組織細胞的物質交換和呼吸代謝功能[23]。而銅含量對抗氧化酶和代謝酶都有著直接的影響。
3.4 銅對 Cu2+-ATPase、Ca2+-Mg2+-ATPase和Cu-Zn-SOD的影響
Cu2+-ATPase在負責維持生物細胞和組織銅含量穩定、調節銅在吸收平衡上具有重要意義[24]。生物在 Cu2+脅迫條件下短期內會刺激Cu2+-ATPase活性的增強,并且認為當細胞對銅的需求達到飽和后,銅不會被源源不斷地“轉入”細胞中,并且 Cu2+-ATPase活力不會立即降低[25]。因此可以認為水體銅的存在對 Cu2+-ATPase會起到激活作用。從本實驗的結果看,隨銅含量的增加,各組織中的Cu2+-ATPase呈不斷上升的態勢,符合上述的觀點。水體中的Cu2+能夠通過某種機制抑制代謝酶活力,如鑲嵌轉運亞基的基團或競爭性抑制離子通道,從而導致體內離子組分的失衡[26]。當 Ca2+在細胞內聚積,可與各種酶蛋白結合,使其ATP生成量驟降,影響整個細胞的能量代謝[6]。本實驗中 Ca2+-Mg2+-ATPase呈先升后降變化可能與水體中的銅含量積聚的濃度有關。
Cu-Zn-SOD在不同組織器官中活力有顯著差異,一般認為SOD活力從高到低是遵循肝胰腺、鰓、肌肉這樣的規律[24]。本實驗中也得出同樣的結果,在實驗的任何階段都是肝胰腺中Cu-Zn-SOD活力顯著高于血淋巴和鰓部組織。隨著實驗組銅含量增加,各組織中的Cu-Zn-SOD活力有所提高,但并不呈正比。本實驗中2種ATP酶的活力均不與Cu-Zn-SOD活力變化相對應。這是否說明了水體中銅含量的增加促進了 Cu2+-ATPase酶的活力,使得銅進入機體內,當機體內銅達到一定量時改變了代謝能源的水平而影響到抗氧化酶的活力,反映在Cu-Zn-SOD活力呈波浪起伏變化?需要進一步研究證實。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
