大理州溫泉捕食線蟲真菌資源調查研究*
2018-07-26 09:51:08范喜杰楊曉燕
楚雄師范學院學報 2018年3期
關鍵詞:資源
范喜杰,張 欣,張 發,楊曉燕*
(1.大理大學公共衛生學院,云南 大理 671000;2.大理大學農學與生物科學學院,云南 大理 671003;3.大理大學東喜瑪拉雅研究院,云南 大理 671003)
捕食線蟲真菌(Nematode-trapping fungi)是一類以營養菌絲特化形成黏性捕食器官(黏性菌網、非收縮環、黏球和黏性分枝)及機械捕食器官(收縮環)捕食線蟲的真菌[1―2]。由于該類真菌在形態和生態功能上的特殊性及其在線蟲生物防治中的潛在價值,逐漸成為微生物學研究的熱點之一[3―5]。捕食線蟲真菌廣泛分布于世界各地,甚至在南極[6]、熱泉[7]和重金屬污染區[8]等極端環境中都有分布。到目前為止,絕大部分研究都集中在陸生環境中的捕食線蟲真菌[9―11],對于水環境中捕食線蟲真菌的研究相對較少[12―14]。目前全世界已知的2萬多種線蟲,有許多生活在水環境中[15],水體中豐富的線蟲種類必定伴隨著豐富的水生捕食線蟲真菌的存在。從已有文獻報道可知水體中水生真菌資源豐富,生物多樣性極高[16,17]。
眾所周知,溫泉是一種特殊的水生環境,廣泛分布于全球各地。由于溫泉與地球早期環境比較接近,溫泉中的微生物生態系統相對簡單、穩定和封閉。同時,在數千萬年的進化歷程中,溫泉微生物已經具備獨特的生存適應能力,這些微生物所具有的特殊生理機制和蘊涵的獨特基因資源,產生的酶或其他代謝產物及其在生命起源和生物進化過程中所處的特殊地位,已經受到國內外學者的廣泛關注[18―20]。如:有研究者從美國黃石公園溫泉中分離得到嗜熱細菌Thermusaquaticus[21],從云南騰沖溫泉中分離得到硫化葉菌Sulfolobustengchongensis[22],從冰島溫泉中分離得到氫氧化細菌Hydrogenophilusislandicus[23],這些皆表明溫泉中蘊藏著豐富的微生物資源。而目前對溫泉中微生物資源的研究主要針對放線菌、嗜熱真菌、細菌和古生菌等,國內外僅Hu等[24]報道過一株溫泉食線蟲真菌(Arthrobotrysmultisecundaria),該菌分離自云南騰沖溫泉,表明溫泉捕食線蟲真菌資源還未引起國內外學者的關注。云南省溫泉資源非常豐富,是中國地熱資源最豐富的地區之一,所以本研究對大理州人為干擾較少的溫泉進行捕食線蟲真菌資源調查,了解溫泉中捕食線蟲真菌資源的分布狀況,分離、鑒定和保存溫泉捕食線蟲真菌資源,為捕食線蟲真菌的起源和進化研究提供菌種資源。
1.材料和方法
1.1 樣品采集
2012年3―5月,采集大理市、祥云縣、彌渡縣、劍川縣、漾濞縣的溫泉底泥共105份,樣品于4℃保存,7d內處理完所有采回的泥土樣品。樣品采集的同時測定溫泉pH值和溫度。
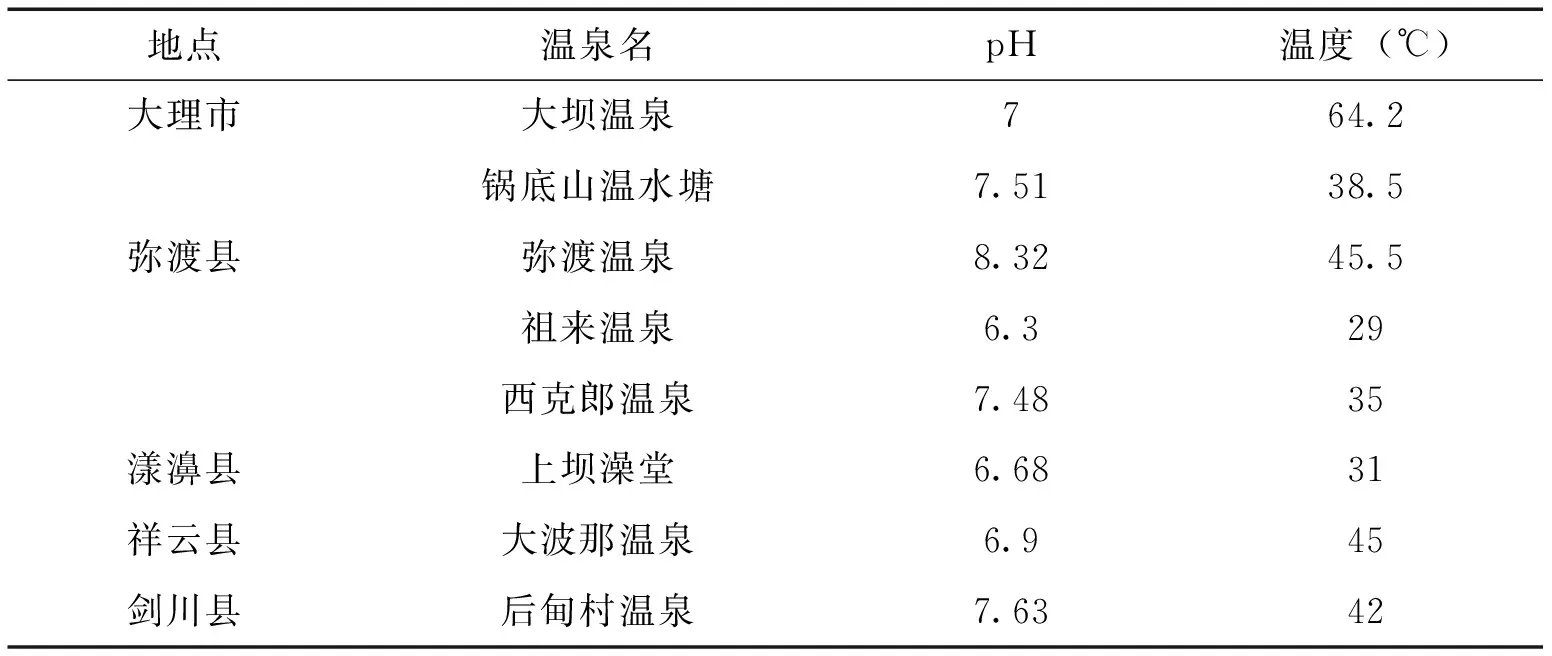
表1 各溫泉基本信息
1.2 CMA培養基的制備
玉米粉20g,瓊脂18g,水1000mL,自然pH。玉米粉加水適量煮沸30min,四層紗布過濾,濾液加水至1000mL,混勻,分裝,121℃高溫滅菌30min[25]。
1.3 誘餌線蟲的制備
采用貝爾曼漏斗法[26]制備:即取少許全齒復活線蟲Panagrellusredivivus放于四層擦鏡紙上置于漏斗(漏斗通過軟管與收集管相接),加適量無菌水,擠壓軟管排出氣泡,靜置一定時間,直至濾出物達到需要量,加無菌水清洗線蟲,使線蟲濃度約為5×103條/mL。
1.4 捕食線蟲真菌的培養、分離與鑒定
采用常規撒土法培養[27],每個樣品設置3個重復,撒樣后在平板中加入上述線蟲懸液1ml(約5000條),室溫下誘導培養4周后在體視鏡下進行鏡檢,將鏡檢到的菌株轉接到CMA平板上,并在培養基中間挖一個2×2 cm2的觀察室,在30℃的恒溫箱內培養5―7d,待觀察室長滿菌絲后加入誘餌線蟲,培養12―24h后進行鏡檢,觀察捕器的產生情況[28]。根據孢子著生方式、孢子梗形態、孢子形態及捕器類型等特征采用Li等[29―30]分類系統對捕食線蟲真菌進行形態學鑒定。
1.5 數據處理
檢出頻率(OF):OF = 某個種出現的土樣數/總的土樣數×l00% 。
Ni為第i種的數量;N為所有種的數量;Pi為第i個種所占百分數;S為采樣點的物種。
2.結果
從大理州溫泉中共分離到2屬14種捕食線蟲真菌,其中Arthobotrys12個種,Dactylellina2個種;在祥云縣的大波那溫泉中分離出一個新種(A.xiangyunensis,sp.nov)[32];A.musiformis為溫泉捕食線蟲真菌的優勢種(檢出率為26.92%)。彌渡縣的彌渡溫泉中捕食線蟲真菌多樣性較高,為0.99,其次為祥云縣的大波那溫泉(0.69);產黏性菌網的捕食線蟲真菌占總檢出的86.54%(表2)。

表2 溫泉底泥中捕食線蟲真菌資源調查結果
3.討論
本研究從大理溫泉中共分離、鑒定出2屬14種捕食線蟲真菌,發現一個新種(A.xiangyunensissp.nov)[32],表明大理溫泉中存在較為豐富的捕食線蟲真菌資源。在調查的溫泉中,各溫泉(包括縣內的溫泉之間)檢出種皆不同,其主要原因可能是:溫泉作為與地球早期生境最為接近的一種特殊生境,其中保留了最為原始的菌種資源[21―23,33,34],在地球生物演化的過程中受到的人為干擾較少,使得空間距離上較為接近的溫泉之間保留了各自的特有物種。同時,各溫泉間的生境差異,如pH和溫度的差異,也可能會導致生活在其中的菌種的不同。捕食線蟲真菌的最適生長溫度為25―28℃,而本次調查的溫泉溫度范圍為29―64.2℃, 表明捕食線蟲真菌的溫度適應能力也存在菌株差異,這種差異由遺傳物質導致。因此,對溫泉捕食線蟲真菌菌種資源的發掘可獲得常規生境中捕食線蟲真菌沒有的基因資源。隨著人類對溫泉的開發加劇[35―37],分布其中的獨特的菌種資源也必將受到影響,因此盡快發掘和保存溫泉菌種資源是一項很迫切的研究工作,同時,應當從政府層面加強對溫泉資源的保護。A.musiformis在大理溫泉中的檢出率最高,是大理溫泉中捕食線蟲真菌的優勢種群;A.oligospora在其中檢出率較低,且僅在大理市的大壩溫泉中有檢出,但已有文獻報道A.oligospora是不同生境土壤中的優勢種群[6, 38,39],表明不同生境中捕食線蟲真菌的分布存在一定差異,產相同捕食器官的捕食線蟲真菌也存在環境適應能力的差異。
據化石記錄顯示,食肉類真菌大約起源于1億年前的中生代,在當時,這個古老的類群主要營腐生生活,但在漫長的進化歷史中,其菌絲體為適應生存環境(如氮源缺乏),可變態為各種形態獨特、結構精巧的捕食器官捕食線蟲[40,41]。因此,捕食器官的進化也就成為該類真菌進化的主要特征之一,不同基于捕食器官形態、捕食率和分子系統發育學特性的進化假說也相繼被提出[25,30,42―45]。但在這些假說中,各捕器類型的進化地位也各不相同,甚至完全相反,如:李天飛等[25]和Yangetal.,[43]提出黏性菌網是一種比較古老、進化地位低且捕食率也較低的捕器類型,但Lietal[30]、Rubner[44]和Enceetal.[45]則認為黏性菌網是一種進化地位比較高且捕食率也較高的捕器類型。林慧彪等[46]對產不同捕器類型的捕食線蟲真菌進行捕食率的定量測定后發現,產黏性菌網的捕食線蟲真菌的捕食率相較產其他捕器類型的捕食線蟲真菌低,而本研究結果也顯示產黏性菌網的捕食線蟲真菌為大理溫泉中的優勢種群,考慮到溫泉生境的特殊性,產黏性菌網的捕食線蟲真菌可能更原始。
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
資源節約與環保(2022年8期)2022-09-20 02:25:22
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
藝術品鑒(2020年7期)2020-09-11 08:04:44
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
當代貴州(2018年28期)2018-09-19 06:39:04
資源再生(2017年3期)2017-06-01 12:20:59
決策(2015年9期)2015-09-10 07:22:44
