新疆博樂和伊犁河谷敏麻蜥的種下分類地位探討
2018-07-25 03:16:24龔雄劉金龍周天和宋琪郭憲光
四川動物 2018年4期
龔雄, 劉金龍, 周天和, 宋琪, 郭憲光
(1.中國科學院成都生物研究所,成都610041; 2. 中國科學院大學,北京100049)
2017年6月,在新疆野外考察中于博樂市連霍高速過四臺路段的戈壁生境(81.692 29°E,44.615 63°N,海拔1 324 m)采集到30余號麻蜥屬Eremias標本,經鑒定均為敏麻蜥Eremiasarguta。然而,其外部形態特征與準噶爾盆地記錄分布的敏麻蜥東方亞種E.a.potanini差異較大,而與伊犁河谷敏麻蜥種群的形態特征相似。經查閱相關文獻(趙爾宓等,1999;戴鑫等,2006),博樂雖有敏麻蜥的記載,但一直缺乏對其種下分類地位的探討。長期以來,多數國內學者認為準噶爾盆地的敏麻蜥為東方亞種,伊犁河谷的為指名亞種E.a.arguta(趙爾宓等,1999)。近年來,Poyarkov等(2014)基于線粒體細胞色素b(Cytb)基因的研究發現,哈薩克斯坦伊犁河谷的敏麻蜥為一獨立進化的支系,是一個未描述的新亞種Eremiasargutassp.。然而,他們的研究并未涵蓋中國的標本,中國伊犁河谷敏麻蜥的種下分類地位仍不清楚。因此,本研究擬聯合形態學和分子系統學的方法對新疆博樂和伊犁河谷的敏麻蜥種群進行種下分類識別和比較研究,旨在探討其分類地位,為深入研究亞洲內陸干旱區敏麻蜥的譜系地理結構和演化歷史積累有價值的資料。
1 材料和方法
1.1 樣品來源
用于分子系統發育分析的敏麻蜥樣品共28號,包括從新疆博樂隨機選取的10號樣品,以及用于比較的采自新疆托里縣、伊寧縣和鞏留縣的各6號樣品。標本經75%乙醇固定后,保存于中國科學院成都生物研究所兩棲爬行動物標本館。所有肝臟組織樣品均固定于95%乙醇,于-20 ℃保存。從GenBank下載Pavlicev和Mayer(2009)以及Poyarkov等(2014)發表的敏麻蜥Cytb基因序列共61條,用于分子系統發育分析。樣品采樣點的分布見圖1,所有樣品和序列的信息見表1。
1.2 方法
1.2.1形態觀測鑒于本研究側重于探討敏麻蜥亞種的分類地位,而不是種群內形態變異,故從博樂采集的標本中僅選取了雌、雄成體各2號作為代表用于形態測量。按照性狀選取原則,并參考戴鑫等(2006)對麻蜥屬的形態學研究和劉金龍等(2014)對蟲紋麻蜥Eremiasvermiculata的形態描述,選取了13個常規性狀進行測量和計數:頭體長(snout-vent length,SVL)、尾長(tail length,TL)、頭長(head length,HL)、頭寬(head width,HW)、頭高(head depth,HD)、前肢長(foreleg length,FLL)、后肢長(hindleg length,HLL)、腹鱗行數(number of ventral scales in a longitudinal row across belly,VS)、最長橫向腹鱗數(number of ventral scales in the longest line,NVLL)、股孔數(number of femoral pores,FPRS)、股孔間鱗數(number of scales between the femoral pores,VBFP)、喉鱗數(gulars,G)和尾部鱗數(number of caudal scales,CDA)。數顯游標卡尺(精確到0.1 mm)測量標本的可量性狀。
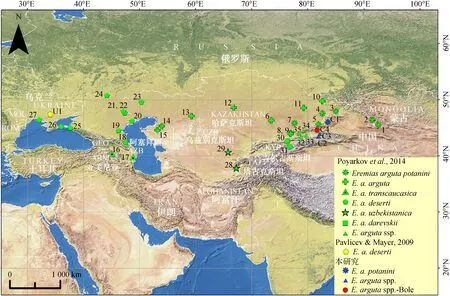
圖1 敏麻蜥采樣點分布
Fig. 1 Sampling localities ofEremiasarguta
阿拉伯數字和U1、C1~C4代表種群編號, 對應表1中的樣點號; 不同圖標代表敏麻蜥的各主要支系, 同圖4和圖5
Populations ofE.argutaused in this study are shown by the numbers corresponding to those in table 1; icons represent the main lineages ofE.argutain fig. 4 and fig. 5
1.2.2基因組DNA提取、PCR擴增及測序取約20 mg肝臟組織,采用核酸純化試劑盒(EasyPure Genomic DNA kit EE101-01, 北京全式金生物技術有限公司),按照說明書操作步驟提取基因組DNA。用超微量紫外檢測儀(NanoDrop 2000C,Thermo Scientific)和瓊脂糖凝膠電泳檢測DNA濃度和純度。最后,將提取的總DNA置于-20 ℃保存備用。
PCR擴增參考Burbrink等(2000)和Zhao等(2011)使用的Cytb引物,結合實際擴增效果對引物的個別堿基進行了優化和調整,最終使用改良后的引物:CytbF:5’-AACCGCCGTTGTTATTCAAC-3’和CytbR-A:5’-TGGTTTACAAAACCAGGGCTTTAGGTT-3’。所有引物均由成都擎科梓熙生物技術有限公司合成。PCR反應Mix采用成都擎科梓熙生物技術有限公司TSINGKE Master Mix (blue)。PCR反應體系為20 μL:2×TSINGKE Master Mix (blue) 10 μL,ddH2O 8 μL,引物各0.5 μL,DNA模板1 μL。PCR反應條件:94 ℃預變性5 min;94 ℃變性30 s,42 ℃退火30 s,72 ℃延伸80 s,35個循環;72 ℃終末延伸10 min。每次PCR擴增均設置陰性對照,以確保實驗過程不受外源DNA的影響。取PCR產物4 μL于1%瓊脂糖凝膠電泳檢測,陽性PCR產物送成都擎科梓熙生物技術有限公司進行雙向測序。測序引物為:CytbF1-A:5’-GGCTCAAACAACCCAACAGGAC-3’和CytbR1-A:5’-TGCGGGGGTAAAA-TTTTC-3’。
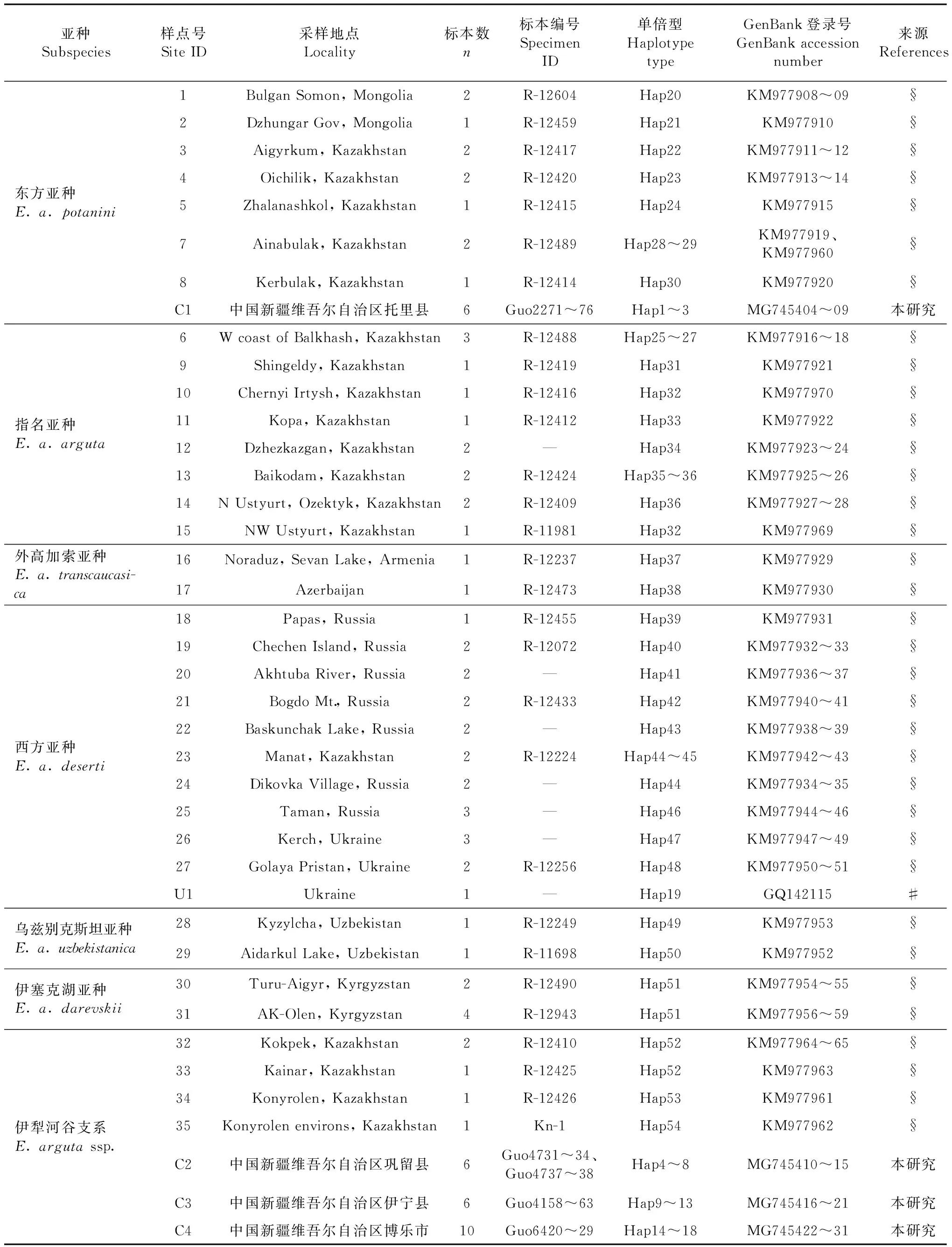
表1 敏麻蜥樣品信息和序列GenBank登錄號Table 1 Samples and sequences information of Eremias arguta
注 Notes: # Pavlicev & Mayer, 2009; § Poyarkovetal., 2014
1.2.3數據分析首先使用DNAStar(DNAStar Inc.,Madison,WI,USA)中的SeqmanⅡ查看所測得的DNA序列峰圖并進行正反鏈校對和編輯,手工去除序列兩端的引物,獲得Cytb基因全序列1 143 bp。將獲得的序列在NCBI中用BLAST(Altschuletal.,1997)進行序列同源性搜索,確保所得的序列是目標基因。在Seaview 4.2.5(Gouyetal.,2010)中進行序列多重比對,并輔以手工校正。在MEGA7(Kumaretal.,2016)中進行序列組成和特征分析,并用脊椎動物線粒體遺傳密碼子將DNA序列翻譯成氨基酸,以確認蛋白編碼基因是否具有功能。在序列中沒有發現提前終止密碼子,提示所獲得的序列不是細胞核線粒體假基因拷貝(Songetal.,2008)。整合從GenBank下載的序列,共計89條序列一起分析。使用DnaSP 5.0(Librado & Rozas,2009)識別單倍型,再用所有的單倍型序列以及外群序列重建系統發育樹。
根據Poyarkov等(2014)對敏麻蜥系統發育的研究,以快步麻蜥Eremiasvelox(GenBank登錄號:KF999319)和中介麻蜥Eremiasintermedia(GenBank登錄號:MG745432)為外群,并以草甸蜥蜴Darevskiapraticola(GenBank登錄號:KM977968)賦根。采用最大簡約法(maximum parsiomony,MP)、最大似然法(maximum likelihood,ML)和貝葉斯推斷法(Bayesian inference,BI)構建單倍型的系統發育樹。在PAUP*4.0b10(Swofford,2003)中進行最大簡約分析,采用啟發式搜索(heuristic search)、10次隨機序列加入重復、樹二等分再連接選項(tree-bisection reconnection),通過Bootstrap方法經1 000次疊加重復估計節點支持率(bootstrap support,BP)。使用RAxMLHPC 8.1.20(Stamatakis,2006)進行最大似然分析,對密碼子一、二、三位點進行分割,選擇GTR+G為核苷酸替代模型,對每個分區單獨進行模型參數推斷,通過Bootstrap方法經1 000次疊加重復估計節點支持率。采用Partitionfinder 2.1.1(Lanfearetal.,2017)基于貝葉斯信息標準(Bayesian information criterion;Schwarz,1978)選取用于BI分析的最優核苷酸替代模型,在MrBayes 3.2(Ronquistetal.,2012)中重建BI樹。密碼子一、二、三位點分別采用K80+G、HKY+I、GTR+G模型。起始樹為隨機樹,以3條熱鏈、1條冷鏈的馬爾可夫鏈蒙特卡羅(Markov Chain Monte Carlo)方法運行2 000萬代,每1 000代抽樣1次,最初5 000棵樹作為老化樣本(burn-in)被舍棄,余下的15 001棵樹用于估算50%多數一致樹,分支節點的可信度以貝葉斯后驗概率(posterior probability,PP)表示。
采用貝葉斯因子(Bayes factor,BF;Kass & Raftery,1995)檢驗相互競爭的系統發育假設。2個不同假設H0和H1的貝葉斯因子等于H0和H1邊際似然值(marginal likelihood)的比值。由于邊際似然值很難直接計算,可用調和平均數(harmonic-mean)來代替(Newton & Raftery,1994)。采用Suchard等(2001)改進的方法,在Tracer 1.5.0(Rambaut & Drummond,2009)中估算調和平均數。因此,BF可表示為2種不同系統發育假設之間調和平均數的比值。本研究檢驗了5種關于敏麻蜥伊犁河谷種群的分類地位的相關假設:(1)伊犁河谷支系E.argutassp.位于進化樹的基礎位置,即與其余支系(亞種)形成姐妹群關系;(2)伊犁河谷支系屬于指名亞種(趙爾宓等,1999);(3)伊犁河谷支系屬于東方亞種(Szczerbaketal.,1993);(4)伊犁河谷支系屬于烏茲別克斯坦亞種E.a.uzbekistanica(Chernov,1934);(5)指名亞種和東方亞種聚成一支,但各自均不形成單系(Poyarkovetal.,2014)。將這些假設均設定為“硬性”約束,在MrBayes 3.2中分別進行貝葉斯系統發育分析,然后分別和“非約束”的貝葉斯系統發育樹(零假設)進行比較。如果2lnBF≥10,則認為有很強的證據(顯著)拒絕備擇假說(Kass & Raftery,1995)。
2 結果
2.1 形態學比較
2.1.1形態特征標本外觀見圖2,外形度量見表2。體型粗壯而稍平扁,尾部短粗,尾長不超過頭體長的1.5倍;頭短寬,顳部稍膨大;吻圓鈍,吻部背面有一縱溝從吻端向后延伸至額鱗中部;吻端有1枚大吻鱗,后接2枚左右上鼻鱗;左右鼻孔各圍有鼻鱗3枚;額鼻鱗1枚;前額鱗2枚;額鱗1枚;大形眶上鱗2枚;額頂鱗2枚;頂鱗2枚;頂間鱗1枚,較小;無枕鱗;眶下鱗不楔入上唇鱗之間;頸部與頭寬相等;趾外側無櫛緣;尾部第9環或第10環鱗片數大于18枚;尾形短圓,尾部鱗片呈棱狀,平滑或輕微向后凸起,有的鱗末端鈍尖;指(趾)短,指(趾)長的順序均為4-3-2-5-1,爪細尖,呈灰白色;大腿兩側的股孔列均未到達膝關節處,其間距至少有1~2枚鱗片;兩側股孔列于肛門前,相距較窄,其間隔7~9枚細鱗;腹鱗行數37~38,腹部一橫列鱗16~17枚;背部黑灰色,有8~9縱列醒目的黑緣白斑,呈不規則分布,繁殖季節雄性成體在背部側面會呈現出明亮的藍色眼點,雄性亞成體則呈淡黃色偏綠色的眼點;而繁殖季節時的雌性成體背部側面為白色眼點,亞成體背部側面有明亮的黃色眼點。

圖2 新疆博樂敏麻蜥成體,雌性(左)、雄性(右,顯示體側藍綠色眼點)(龔雄 攝)Fig. 2 General view of adult Eremias arguta in Bole city, Xinjiang Uyghur Autonomous Region, China,female (left) and male (right, with bluish-greenish ocelli)(photo by GONG Xiong)
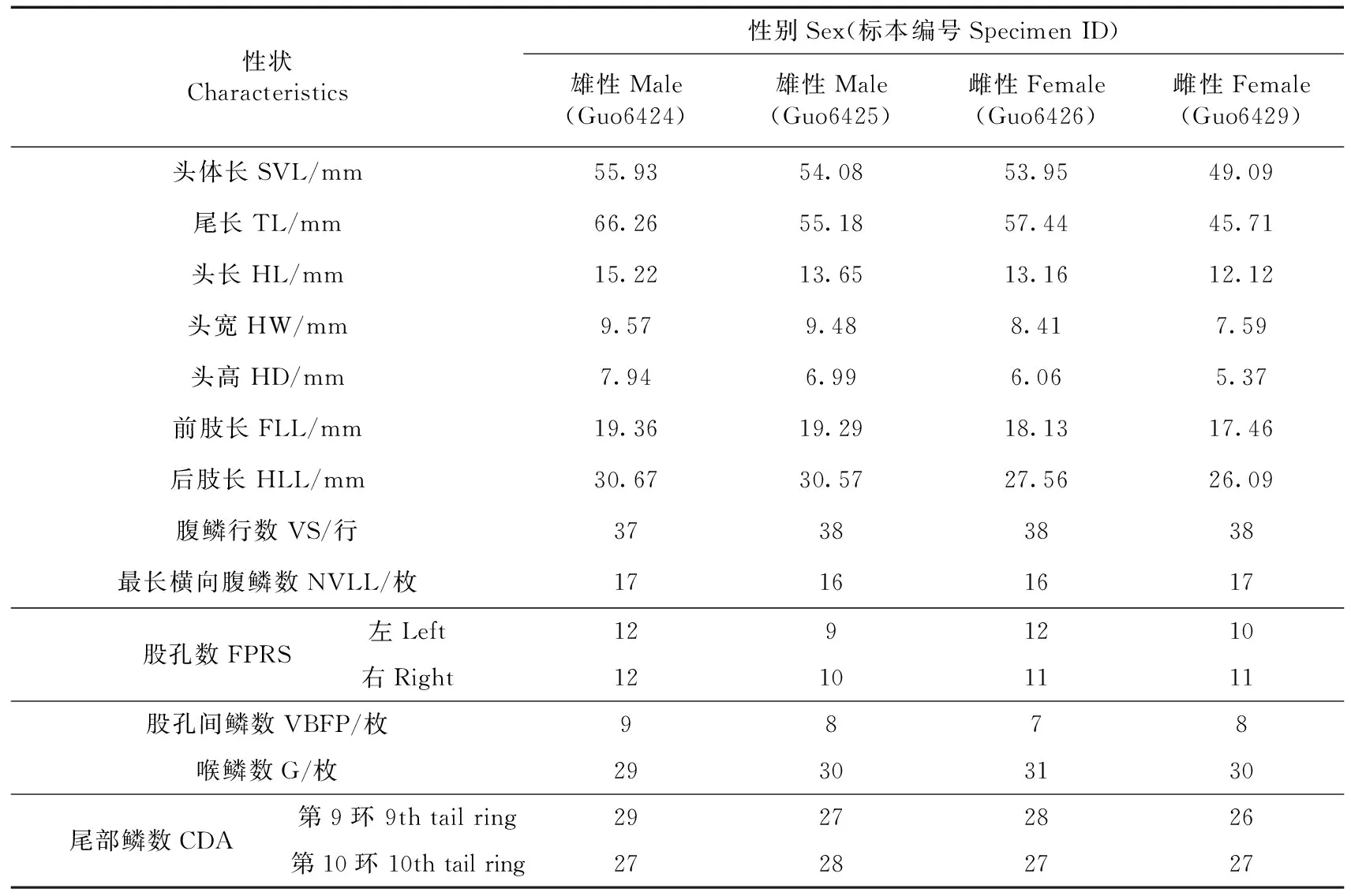
性狀Characteristics性別Sex(標本編號Specimen ID)雄性Male(Guo6424)雄性Male(Guo6425)雌性Female(Guo6426)雌性Female(Guo6429)頭體長SVL/mm55.9354.0853.9549.09尾長TL/mm66.2655.1857.4445.71頭長HL/mm15.2213.6513.1612.12頭寬HW/mm9.579.488.417.59頭高HD/mm7.946.996.065.37前肢長FLL/mm19.3619.2918.1317.46后肢長HLL/mm30.6730.5727.5626.09腹鱗行數VS/行37383838最長橫向腹鱗數NVLL/枚17161617股孔數FPRS左Left1291210右Right12101111股孔間鱗數VBFP/枚9878喉鱗數G/枚29303130尾部鱗數CDA第9環 9th tail ring29272826第10環 10th tail ring27282727
2.1.2形態比較敏麻蜥分布區東緣3個亞種或支系的形態特征比較見表3。東方亞種的體型更為粗壯,成體個體更大,尾部更粗短,背面灰黃色,飾有6~9行縱列斑帶,為黑緣白斑橫斷及由此聯結所成的橫紋,但是呈更大的拉伸和橫斷的延長,大腿兩側的股孔列均未到達膝關節處,中間有1~2枚鱗片,2列股孔數多為9~11枚;指名亞種背斑為不規則的黑色橫斷的斑點和條紋,而伊犁河谷敏麻蜥多為縱列的黑緣白斑(Szczerbak,1974,2003;趙爾宓等,1999)。于博樂采集的敏麻蜥,外部形態特征和度量數據與準噶爾盆地分布的東方亞種相比,在體型、背斑和股孔數等方面差別很大;與指名亞種相比,主要為背斑差異和在繁殖季節體側出現了特殊的眼點,而與伊犁河谷分布的敏麻蜥的描述相符(Dujsebayevaetal.,2007)。
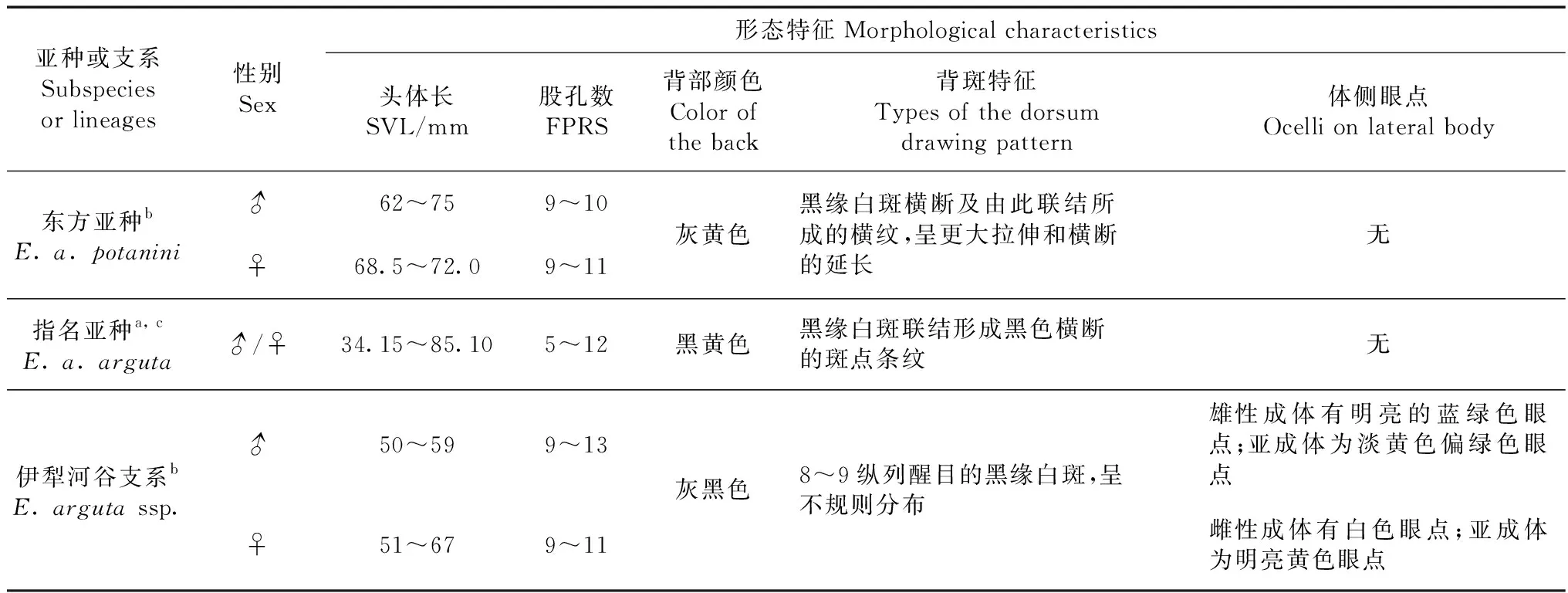
表3 敏麻蜥分布區東緣3個亞種或支系的形態特征比較Table 3 Comparisons of the morphological characteristics of the 3 subspecies or lineages of Eremias arguta in the eastern periphery of distribution area
注:aSzczerbak等, 1993;b趙爾宓等, 1999;cSzczerbak, 2003
Note:aSzczerbaketal., 1993;bZhaoetal., 1999;cSzczerbak, 2003
2.2 生境比較
新疆博樂發現的敏麻蜥棲息地為天山以北一片荒漠戈壁(圖3),生境單調,地表土壤主要為沙土礫石,動物種類較少,除同棲分布的沙蜥和偶見的天山黃鼠Citellusrelictus和蛇類的洞穴外,幾乎無其他脊椎動物。分布的步甲科Carabidae和擬步甲科Tenebrionidae甲蟲、蜘蛛、黑螞蟻和一些蝴蝶的幼蟲等為敏麻蜥食物來源。主要植被包括北疆錦雞兒Caraganacamilli-schneideri、芨芨草Achnatherumsplendens和一些低矮的草本植物。敏麻蜥主要在低矮的北疆錦雞兒叢中及其周圍活動,其洞穴也大多分布在較密的北疆錦雞兒叢和一些較大的巖石底部。
2.3 序列變異和系統發育關系
PCR擴增獲得敏麻蜥Cytb序列(1 143 bp)28條,識別出18個單倍型;從GenBank下載Pavlicev和Mayer(2009)發表的同源序列1條(1 143 bp)和Poyarkov等(2014)發表的同源序列60條(778 bp)。聯合3條外群序列比對生成的數據矩陣,識別出57個單倍型,1 143個同源位點中有變異位點446個,簡約信息位點296個,單一多態位點150個,轉換與顛換之比平均為3.15。
采用3種方法所得到的系統發育樹的拓撲結構基本一致,特別是ML樹和BI樹的拓撲結構幾乎完全一致(圖4、圖5)。3種方法所得到的進化樹均支持敏麻蜥形成一個單系(MP/ML/BI:100%/88%/1.00),并識別出7個明顯的分支(Clade Ⅰ~Ⅶ),分別與基于形態劃分的亞種相對應:敏麻蜥東方亞種、指名亞種、外高加索亞種E.a.transcaucasica、西方亞種E.a.deserti、烏茲別克斯坦亞種、伊塞克湖亞種E.a.darevskii和伊犁河谷支系。其中,采自博樂的敏麻蜥E.argutassp.-Bole的單倍型序列均聚在伊犁河谷支系所在的CladeⅦ(MP/ML/BI:100%/100%/1.00);且與準噶爾盆地分布的東方亞種和哈薩克斯坦中部分布的指名亞種沒有構成姐妹群,二者親緣關系較遠。
圖3 新疆博樂敏麻蜥采集地生境(龔雄 攝)
Fig. 3 Biotope ofEremiasargutain Bole city, Xinjiang Uyghur Autonomous Region, China (photo by GONG Xiong)
MP分析得到了1 001棵同等簡約樹,樹長(tree length)=922,一致性指數(consisteney index,CI)=0.637,留存指數(retention index,RI)=0.856,尺度化一致性指數(rescaled consistency,RC)=0.545。其中,Clade Ⅰ(東方亞種)與Clade Ⅱ(指名亞種)聚成一支(BP=98%),但二者各自形成單系的支持率均很低(BP<50%);Clade Ⅶ(伊犁河谷支系)位于樹的基礎位置,與其余支系形成姐妹群關系,但支持率不高(BP=73%)。BI樹和ML樹中,Clade Ⅰ與Clade Ⅱ聚成一支(PP/BP=1.00/96%),二者形成交互單系(Clade Ⅰ:PP/BP=0.98/53%,Clade Ⅱ:PP/BP=0.80/53%);Clade Ⅴ與Clade Ⅵ聚成一支(PP/BP=0.99/76%),再和Clade Ⅶ形成姐妹群關系,盡管支持率不高(PP/BP=0.88/55%)。
2.4 貝葉斯因子檢驗
采用貝葉斯因子檢驗幾種相互競爭的拓撲結構的結果見表4。伊犁河谷支系屬于指名亞種或東方亞種或烏茲別克斯坦亞種的3種備擇假設均被本研究的分子數據極強地拒絕;而伊犁河谷支系位于系統發育樹的基礎位置,即與其余支系形成姐妹群關系的假設沒有被顯著拒絕。本研究的MP樹(圖4)與BI樹(圖5)間的差異并不顯著(2lnBF=5.94<10)。指名亞種和東方亞種不形成交互單系的假設(Poyarkvaetal.,2014),與本研究BI樹差異不顯著(2lnBF=8.184<10),沒有被很強的證據拒絕。
3 討論
敏麻蜥為麻蜥屬中分布范圍最廣的物種之一,其分布區橫貫歐亞大陸,包括歐洲黑海地區的羅馬尼亞到俄羅斯的北高加索,外高加索(格魯吉亞,亞美尼亞,阿塞拜疆)的東部,中亞五國以及向東一直到中國新疆西北部(伊犁河谷、準噶爾盆地)和蒙古國的西部戈壁地區等(Szczerbak,1974,2003;趙爾宓等,1999;Sindacoetal.,2008)。敏麻蜥為典型的耐干旱物種,因其分布廣泛,生境異質性高,地理種群變異很大,導致種下分類復雜。傳統的形態學研究將敏麻蜥分為敏麻蜥指名亞種、西方亞種、外高加索亞種、烏茲別克斯坦亞種、伊塞克湖亞種和東方亞種6個亞種(Szczerbak,1974,2003;Sindacoetal.,2008;郭憲光等,2010)。
圖4 基于線粒體Cytb基因序列采用PAUP*4.0b10重建的最大簡約嚴格一致樹
Fig. 4 Maximum parsiomony strict consensus tree based onCytbgene sequences using PAUP*4.0b10
節點上的數值為1 000次重復抽樣分析所得到的自展支持率(僅顯示>50%); 樹長=922, 一致性指數=0.637, 留存指數=0.856, 尺度化一致性指數=0.545
Numbers above the nodes represent bootstrap support with 1 000 pseudoreplicates (only shown BP>50%); tree length=922, consistercy index=0.637, retention index=0.856, rescaled consistercy=0.545
節點上的數值代表貝葉斯后驗概率/最大似然自展支持率(僅顯示PP>0.50或BP>50%), — 表示節點的自展支持率小于50%或此節點不存在
Bayesian posterior probabilities (>0.50) and maximum likelihood bootstrap values (>50%) are shown, respectively; — represents node with bootstrap value lower than 50% or node not existed
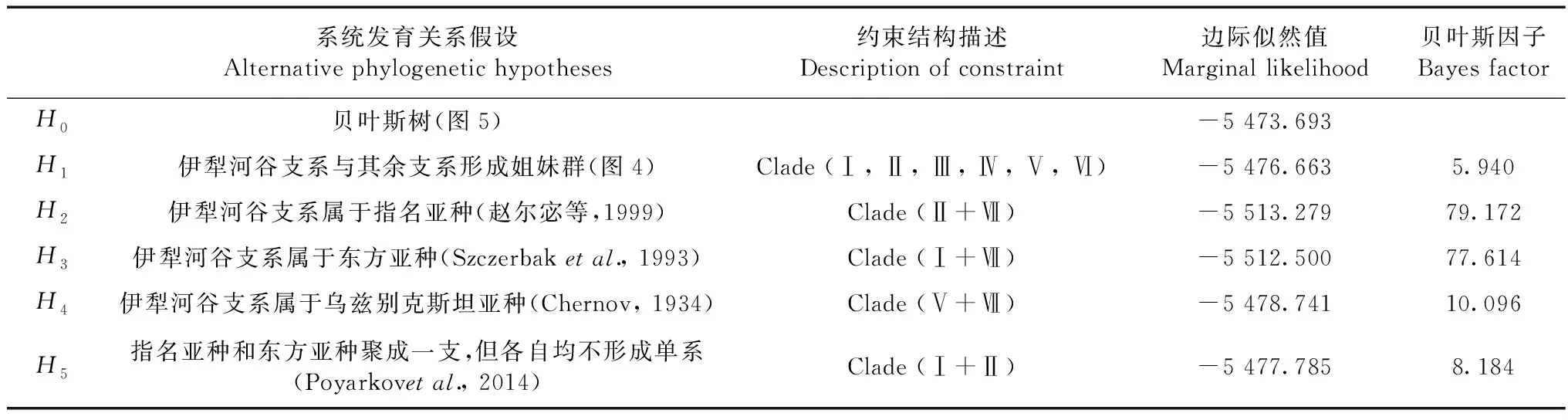
表4 各個備擇系統發育假設的貝葉斯因子檢驗Table 4 Bayes factors for comparisons of alternative phylogenetic hypotheses
3.1 博樂種群的分類地位
根據《中國動物志 爬行綱(第二卷) 有鱗目 蜥蜴亞目》(趙爾宓等,1999)記載,新疆博樂有敏麻蜥分布,但志書并未對博樂種群進行分類討論,其種下分類地位不明確。戴鑫等(2006)對中國8種麻蜥開展了多變量形態比較,其中敏麻蜥包括博樂的2號標本,未提及其亞種地位。本研究重新檢視了這2號標本:采集號625134(館藏號CIB3531)、625135(館藏號CIB3532),標簽上記錄分別于1958年8月15日和29日采自博樂城郊草壩。這2號標本經福爾馬林固定,由于年代相對較久遠,一些關鍵特征不易辨識,難于鑒定到亞種,館藏記錄僅鑒定至種級水平,推測編寫志書時,沒有進一步討論其種下分類地位。志書在相關部分將中國的敏麻蜥歸為指名亞種(分布于尼勒克、新源、伊寧等地)和東方亞種(塔城-額敏地區自西向東直到富蘊-青河一帶)。
然而,此次在博樂采集的敏麻蜥標本,其形態特征與準噶爾盆地分布的東方亞種相比,在體型、背斑和股孔數等方面差別很大;與指名亞種相比,主要為背斑差異和在繁殖季節出現了特殊的眼點,而與伊犁河谷地區的敏麻蜥描述(Dujsebayevaetal.,2007)相符。在生境方面,此次博樂敏麻蜥的采集地生境與趙爾宓等(1999)描述的伊犁河谷敏麻蜥生境和Dujsebayeva等(2007)在哈薩克斯坦東南部伊犁河谷地區發現的敏麻蜥生境相似。而敏麻蜥東方亞種的棲息地主要為干枯河床的礫石堆下(趙爾宓等,1999),與此次博樂敏麻蜥的采集地生境有較大區別。基于Cytb基因的分子系統發育分析表明,MP、ML和BI樹均一致地支持其與伊犁河谷樣本聚成一支。因此,此次于新疆博樂發現的敏麻蜥應劃分為伊犁河谷支系,隸屬于敏麻蜥中未描述的一個新亞種(Poyarkovetal.,2014)。這一發現暗示,阿拉套山、科爾古琴山和博羅科努山可能并未對伊犁河谷和博樂的敏麻蜥種群的基因流構成明顯的地理阻隔。這是敏麻蜥伊犁河谷支系在準噶爾盆地有分布的新紀錄,推測準噶爾盆地還有更多的分布點。此外,有趣的是,博樂的敏麻蜥種群似乎與東方亞種的分布區相連,伊犁河谷支系與東方亞種是否有同域分布的種群,有待進一步考察研究。
3.2 伊犁河谷支系的系統發育位置
長期以來,伊犁河谷敏麻蜥種群的分類地位主要有4種觀點:Chernov(1934)認為屬于烏茲別克斯坦亞種;Yakovleva(1964)認為屬于烏茲別克斯坦亞種和指名亞種中間類型;Sczcerbak等(1993)將其修訂為東方亞種;趙爾宓等(1999)將其劃為指名亞種。Dujsebayeva等(2007)發現,伊犁河谷敏麻蜥雄性成體在繁殖季節體側有醒目的藍色眼點,而敏麻蜥其他亞種均無這一特征,這暗示了伊犁河谷的敏麻蜥種群可能處于一個獨特的分類地位。Orlova等(2012)通過多變量判別分析,發現伊犁河谷的敏麻蜥種群聚成單獨的一簇,可與其他幾個亞種分開。Guo等(2011)基于16SrRNA基因片段探討了麻蜥屬部分物種的分子系統發育和分歧時間,但并未涉及敏麻蜥種內關系的探討。Poyarkov等(2014)基于Cytb基因片段,首次從分子上發現伊犁河谷種群不是指名亞種、烏茲別克斯坦亞種或東方亞種間的過渡形式,而是形成一個獨立的支系,與其余支系形成姐妹群關系,盡管支持率不高(MP/ML/BI:71%/65%/0.93),可能是一個未描述的新亞種。遺憾的是,Poyarkov等(2014)的研究未包含中國分布敏麻蜥的代表樣本。
本研究在Poyarkov等(2014)的基礎上,增加中國新疆博樂、伊寧和鞏留的敏麻蜥種群以及塔城盆地(托里縣)東方亞種種群的代表樣品。重建的MP樹重現了Poyarkov等(2014)關于伊犁河谷支系的系統發育位置,即位于樹的基礎位置,與其余支系形成姐妹群關系(BP=73%);而ML和BI樹中,伊犁河谷支系與烏茲別克斯坦亞種和伊塞克湖亞種形成的分支構成姐妹群關系(ML/BI:0.88/55%)。可見,伊犁河谷支系的系統發育位置尚未解決,下一步研究有必要增加樣品和利用多基因位點來檢驗這2種相互競爭的假設,并在整合分類學(Padialetal.,2010)研究的基礎上,來描述這一新分類亞種單元。
3.3 東方亞種的有效性
Orlova等(2012)發現敏麻蜥整個分布范圍的東部,背部從斑點狀的“arguta”-type到帶狀的“potanini”-type表現出由西至東的漸變。Poyarkov等(2014)發現,敏麻蜥指名亞種和東方亞種聚成一支,但二者并未形成交互單系,東方亞種散布在指名亞種形成的支系中。但本研究的MP、ML和BI樹均支持這2個亞種形成交互單系,盡管MP樹的自展支持率低于50%。通過BF檢驗(2lnBF=8.184<10),發現Poyarkov等(2014)的假設不能被當前的Cytb基因數據很強地拒絕。當然,如果放寬標準,按傳統的6<2lnBF<10來理解(Kass & Raferty,1995),還是有強的證據拒絕“指名亞種和東方亞種不形成交互單系”的假設。也就是說,本研究中,東方亞種的有效性得到了分子數據一定程度上的支持。因此,綜合形態數據的結果,東方亞種是否有效尚不確定,有必要增加更多的數據,引入整合分類學的方法(Padialetal.,2010;Torstrometal.,2014)。鑒于對生物多樣性的認識和保護目的,建議當前保留此亞種,不宜將其視為指名亞種的次定同物異名。
致謝:四川大學華西基礎醫學與法醫學院陳達麗協助新疆伊犁河谷的標本采集,何金蕾和陳晗在標本整理過程中提供了幫助,審稿專家對稿件提出了寶貴意見,在此一并致謝!
