庫爾勒香梨果實可溶性糖積累及代謝相關酶活性變化
2018-07-09 12:48:40艾沙江買買提張校立王繼勛
新疆農業科學 2018年4期
艾沙江·買買提,張校立,梅 闖,馬 凱,閆 鵬,王繼勛
(新疆農業科學院園藝作物研究所/農業部新疆地區果樹科學觀測試驗站,烏魯木齊 830091)
0 引 言
【研究意義】庫爾勒香梨(以下簡稱“香梨”)是新疆“名、特、優”水果之一,香梨已成為新疆庫爾勒市農業增效,農民增收的支柱產業。但目前生產上出現香梨果肉變粗,口感多渣,石細胞含量增高,導致果實品質降低的問題。果實中糖的代謝和積累是影響果實品質的重要因素。研究果實中糖分的代謝及積累對提高香梨品質有實際意義。【前人研究進展】梨屬于木本薔薇科果樹, 葉片光合產物以山梨醇為主要形態, 果實糖的積累類型屬于中間轉化型,果實發育早中期以積累山梨醇和淀粉為主, 至果實發育后期果糖、蔗糖和葡萄糖含量逐漸增多[1-2]。不同栽培種梨果實中糖的含量不同,果糖和葡萄糖是最主要的可溶性糖,果實中果糖含量最高, 占總糖比例的42.22%~57.02%。不同品種之間, 葡萄糖和果糖的含量相對穩定, 而山梨醇和蔗糖含量變化較大[3-9]。目前對于梨果實中糖代謝的研究,主要集中在套袋、疏花疏果等栽培措施對梨果實糖積累的影響以及酶的活性與糖積累的關系上,另外,果實糖積累過程中糖代謝關鍵酶的作用機制以及糖運輸蛋白等方面也有一些報道[10-17]。【本研究切入點】雖然已經明確梨果實的主要糖組分及含量水平差異,但是對不同品種糖積累差異及其產生機制的研究較少,尤其相關代謝酶在庫爾勒香梨果實的糖積累過程中起到作用尚不明確。研究庫爾勒香梨果實發育過程中糖積累特性及相關酶活性的變化規律。【擬解決的關鍵問題】以庫爾勒香梨為試材,分析庫爾勒香梨果實發育過程中不同糖分的積累規律以及糖代謝相關酶活性的變化,研究不同發育時期果實糖分變化與酶活性之間的關系,為香梨糖分積累的分子機制研究和品質改良提供技術參數。
1 材料與方法
1.1 材 料
材料為發育過程中的庫爾勒香梨(PyrusbrestschneideriRehd)果實,果實采自新疆農業科學院輪臺果樹資源圃15年生香梨樹。選擇樹勢健壯、管理水平一致的果樹10株,于2016年4月初香梨梨花蕾期,分別在樹冠的中層南部選擇花蕾發育期及大小基本一致的短果枝掛牌,每短果枝保留兩個果實。于花后15 d開始取樣,前期每10 d取樣一次,每次取大小均勻的果實30個,后期每15 d取樣一次,每次取大小均勻的果實10個冷藏帶回實驗室,用于相關指標的測定。
1.2 方 法
參照趙世杰[18]的方法,采用葸酮比色法測定果實中可溶性糖和淀粉含量,山梨醇、葡萄糖、果糖和蔗糖的測定參考姚改芳的方法[4]采用高效液相色譜法(HPLC)。酸性轉化酶(AI)、堿性轉化酶(NI)、蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)的活性參照許讓偉的方法[10]進行測定;山梨醇脫氫酶(SDH)和山梨醇氧化酶(SOX) 的活性測定參考Yamaguchi的方法[19]。
1.3 數據處理
所有數據采用Excel 2010和 SPSS 17.0 軟件進行處理,顯著水平P<0.05。采用Excel進行作圖。
2 結果與分析
2.1 庫爾勒香梨果實的發育動態變化規律
研究表明,香梨果實發育需要130 d左右,果實縱、橫徑生長曲線類似“S”型。根據果實發育期鮮重和縱橫徑的生長動態,香梨果實發育呈“慢-快-慢”的趨勢。發育前期果實縱徑生長較快,后期橫徑生長較快,香梨在花后70 d進入快速膨大期,坐果后75~120 d是果實快速膨大的關鍵階段,至成熟時,果實增重減緩。
香梨單果重變化趨勢同果實縱橫徑一樣,在發育期呈“S”型逐漸增加的趨勢,花后15~60 d為遲滯期,果實重量增加緩慢;花后51~120 d為對數增長期,果實重量增加迅速;花后 120 d以后為穩定期,果實重量增加減慢,并逐漸穩定。圖1
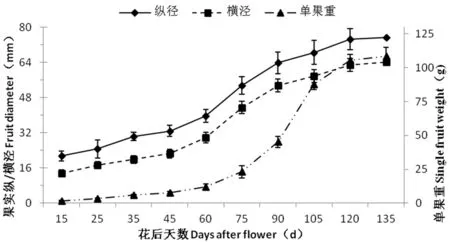
圖1 庫爾勒香梨果實發育過程中縱/橫經、單果重變化
Fig.1 Changes of single fruit weight and fruit diameters during development of Korla fragrant pear
2.2 庫爾勒香梨果實的糖積累動態變化規律
研究表明,隨著果實的不斷生長發育,可溶性總糖含量逐步上升,各糖組分都呈現出升高趨勢。果實發育前期(花后15~60 d)果糖、葡萄糖和蔗糖含量較低并呈現出緩慢增大趨勢,同時期果實中山梨醇和淀粉含量較高,其上升速度較快。花后75 d開始,香梨果實中果糖、葡萄糖和蔗糖的含量呈現出快速增長,而淀粉含量開始迅速下降,山梨醇含量的繼續上升,在果實成熟期(花后105 d )山梨醇含量變化較平穩。從其構成來看,果糖為香梨果實中含量最高的可溶性糖,香梨花后135 d果實的可溶性糖為16.16%,其中果糖、山梨醇、葡萄糖和蔗糖含量分別為6.25%、3.99%、3.54%、2.35%。從糖積累最快的時期來看(花后90~135 d),花后90 d開始,香梨果實中果糖、葡萄糖和蔗糖含量快速上升, 果糖上升幅度最大,其次為葡萄糖、蔗糖的上升幅度最小,山梨醇含量卻開始下降。圖2
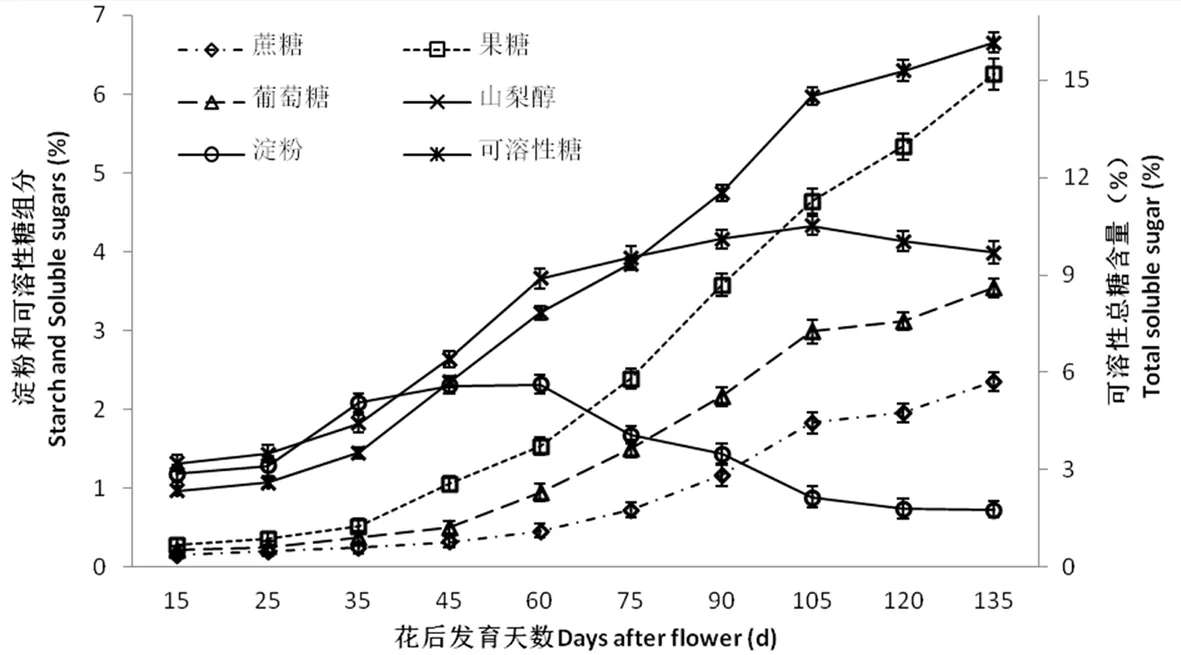
圖2 庫爾勒香梨果實發育過程中不同類型可溶性糖含量變化
Fig.2 Changes in different soluble sugar during fruit development in Korla fragrant pear
2.3 香梨果實發育過程中不同糖組分所占比例變化
研究表明,各組分可溶性糖在總糖中所在的比例不同。在果實發育早期,梨果實中山梨醇的含量占總糖中的比例50%以上。隨著果實的發育,山梨醇所占的比例呈下降趨勢,相反,果糖、葡萄糖和蔗糖所占的比例緩慢上升。果實發育早期(花后60 d以內),山梨醇的比例保持較高水平,下降速度緩慢(由51.1%降到42.1%),相應的,同時期,果糖、葡萄糖和蔗糖占總糖的比例較低,緩慢上升,果糖占總糖的比例由11.9%升至19.6%,葡萄糖占總糖的比例由8.9%升至12.2%,蔗糖占總糖的比例變化幅度很少。果實快速膨大階段(花后70~120 d),山梨醇的比例迅速下降(由42.1%降至24.7%),果糖、葡萄糖和蔗糖所占的比例迅速上升,果實成熟期,各組分在總糖中所占的比重分別為24.7%、38.7%、22.0%、14.5%。圖3
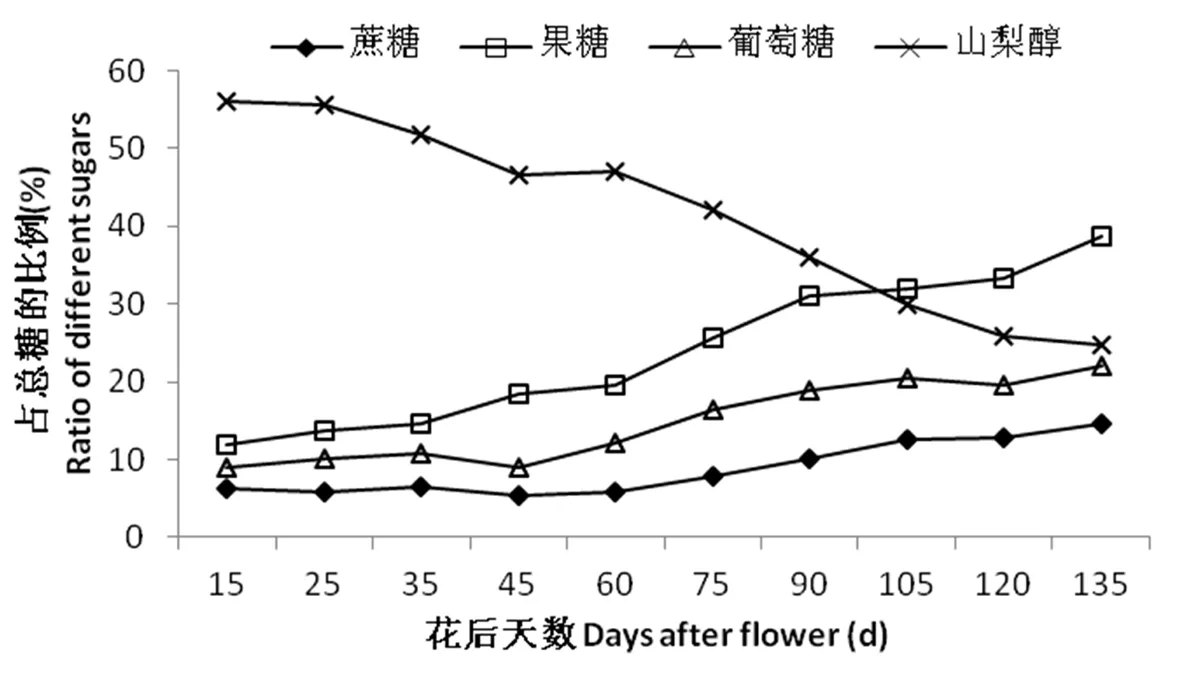
圖3 香梨果實發育過程中不同糖組分所占比例變化
Fig.3 Changes in different soluble sugars ratio during fruit development in Korla fragrant pear
2.4 庫爾勒香梨果實的糖積累動態變化規律
對香梨果實單果重與糖含量間相關性分析。研究表明,香梨果實大小與可溶性糖之間呈極顯著的正相關,香梨果實與糖積累之間具有必然的內在聯系。單果重與蔗糖、果糖、葡萄糖、山梨醇、可溶性總糖之間相關系數r分別為0.981**、0.994**、0.997**、0.860**、0.988**,均達到極顯著水平。不同糖組分都與單果重高度正相關。正是由于各糖組分都與單果重高度正相關,導致各糖組分之間高度正相關。偏相關分析表明,香梨果實發育過程中,果實蔗糖含量與果糖和葡萄糖含量和極顯著正相關,相關系數分別為0.836**和0.845**,與山梨醇含量顯著負相關(r=-0.737*)。山梨醇是香梨果實中的重要可溶性糖,果實發育中,山梨醇含量與蔗糖、果糖和葡萄糖含量呈現負相關關系,而與總糖含量極顯著正相關,說明果實發育過程中,總糖含量的增加,主要貢獻是山梨醇含量的增加,果實山梨醇含量提高有利于香梨總糖的積累。表1
表1 香梨單果重與果實可溶性糖及組分相關性
Table 1 Correlation analysis of single fruit weight and soluble sugar contents of in Korla fragrant pear
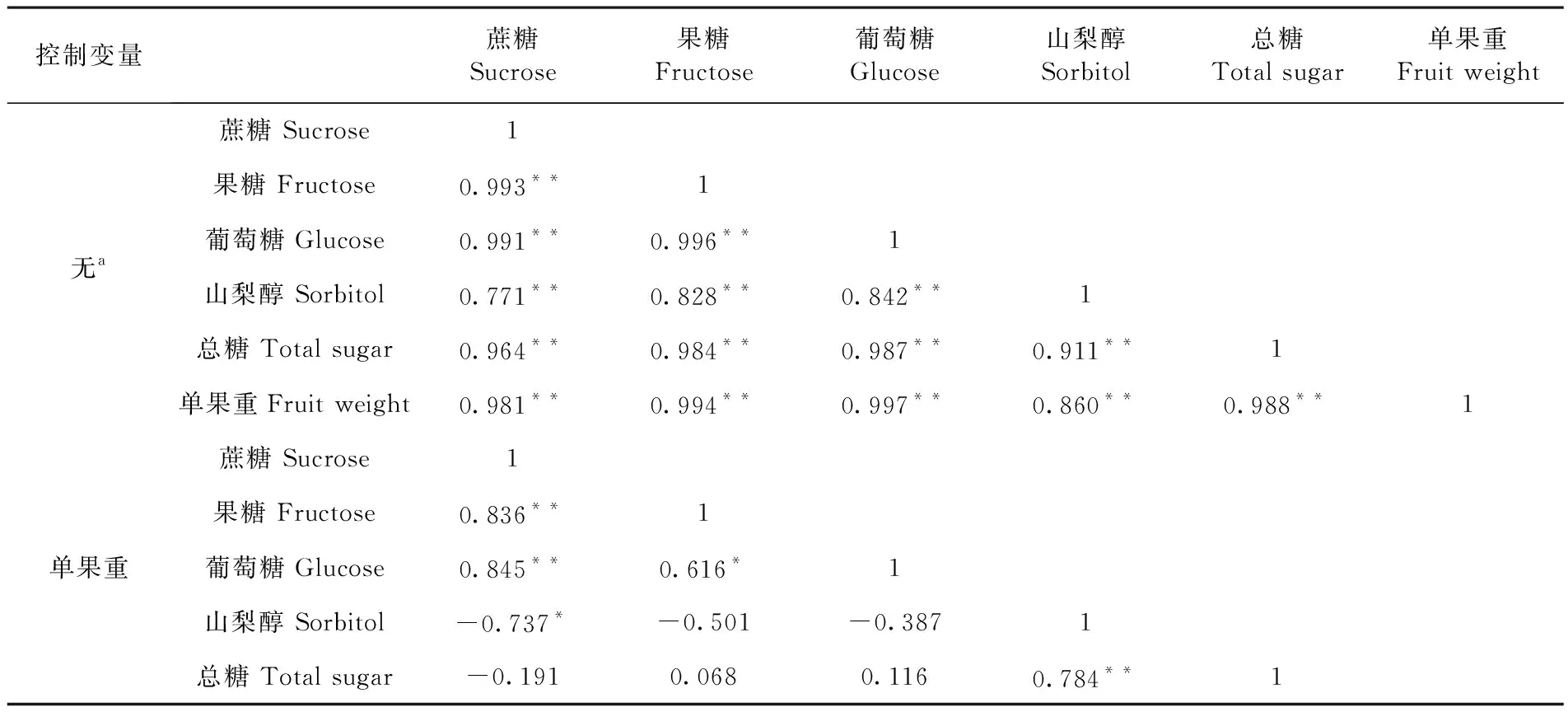
控制變量蔗糖Sucrose果糖Fructose葡萄糖Glucose山梨醇Sorbitol總糖Total sugar單果重Fruit weight無a蔗糖 Sucrose1果糖 Fructose0.993**1葡萄糖 Glucose0.991**0.996**1山梨醇 Sorbitol0.771**0.828**0.842**1總糖 Total sugar0.964**0.984**0.987**0.911**1單果重Fruit weight0.981**0.994**0.997**0.860**0.988**1單果重蔗糖 Sucrose1果糖 Fructose0.836**1葡萄糖 Glucose0.845**0.616*1山梨醇 Sorbitol-0.737*-0.501-0.3871總糖 Total sugar-0.1910.0680.1160.784**1
a. 單元格包含零階 (Pearson) 相關,顯著水平:P<0.05(*),P<0.01(**)
Significant atP<0.05(*),P<0.01(**)
2.5 香梨果實發育過程中SDH和SOX活性的變化
研究表明,在果實發育早期,香梨果實中的山梨醇脫氫酶活性均較高,隨著果實的發育,NAD-SDH 活性先急劇下降,花后60~70 d出現一個低谷(分別為7.10 μmol NAD/(h·g)和7.25 μmol NAD/(h·g)),花后75 d開始逐漸升高,在果實的后期保持較高的水平。SOX的變化趨勢與SDH活性趨勢有類似之處,也是隨著果實的發育先下降后升高。在整個果實發育階段,香梨果實SHD和SOX酶活性變化趨勢雖一致,但前者的變化幅度大,且其活性水平始終顯著高于后者,這可能是香梨果實中果糖含量顯著高于葡萄糖含量的原因之一。相關性分析結果表明,SDH酶活性與蔗糖含量變化顯著正相關(r=0.715*),而與山梨醇和可溶性總糖含量的變化極顯著負相關(r= -0.941**和r= -0.767**)。SOX酶活性只與山梨醇含量的變化極顯著負相關(r= -0.777**)。表2,圖4
2.6 香梨果實發育過程中SS和SPS活性變化
研究表明,果實SPS活性隨果實發育逐步上升。幼果階段(花后15~75 d)果實中SPS 活性較低且保持相對穩定,花后75 d起大幅上升,果實成熟時(花后135 d),SPS活性達到最高,為48.47 μmol/(h·g),SPS活性增高將近3倍左右。SS活性在果實生長發育初期較高,為38.03 μmol/(h·g),隨后呈下降的趨勢,花后60 d降到低谷(18.70 μmol/(h·g)),之后略有上升,花后75 d后SS活性又呈下降趨勢,花后105 d出現第二低谷,之后隨著果實的生長發育活性逐漸升高。相關性分析表明,SPS酶活性與蔗糖和果糖含量變化顯著正相關(相關系數分別為r= -0.821**和r= -0.807**),而與山梨醇含量的變化極顯著負相關(r= -0.838**)。SS酶活性只與可溶性總糖含量的變化顯著負相關(r= -0.663**)。圖5,表2
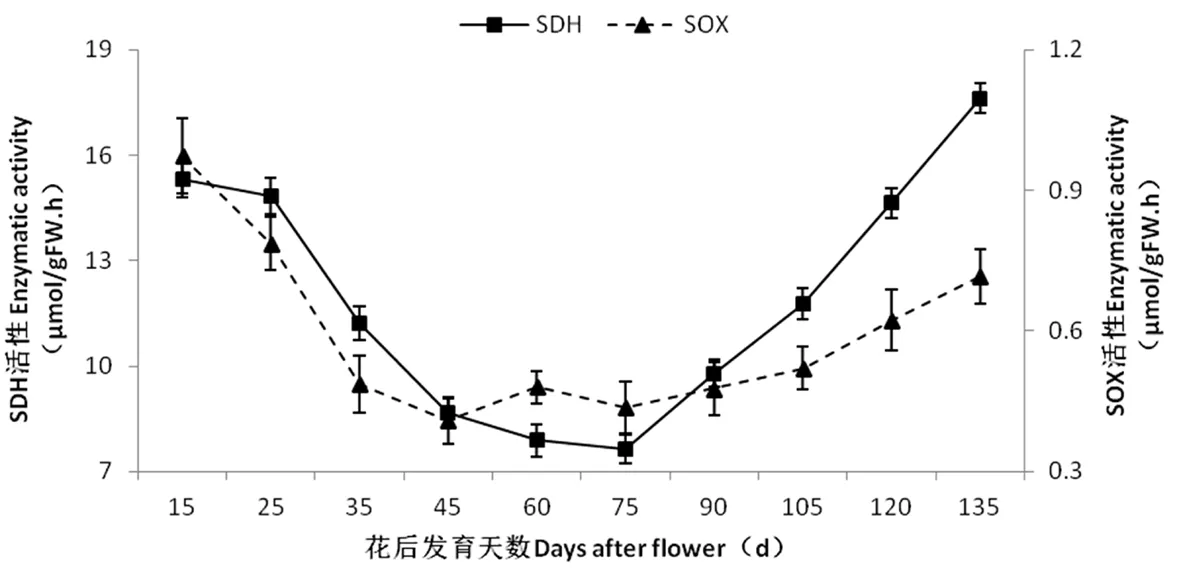
圖4 庫爾勒香梨果實發育過程中山梨醇脫氫酶(SDH)和山梨醇氧化酶(SOX)活性變化
Fig.4 Changes in SDH and SOX activities during the fruit development in Korla fragrant pear
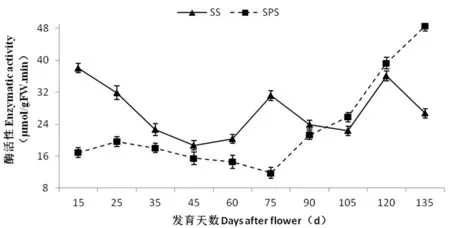
圖5 庫爾勒香梨果實發育過程中蔗糖合酶(SS)和蔗糖磷酸合酶(SPS)活性變化
Fig.5 Changes in SPS and SS activities during the fruit development in Korla fragrant pear
表2 香梨果實中糖組分與相關酶活性相關性
Table 2 Correlation analysis of soluble sugars and related enzymes activity in Korla fragrant pear
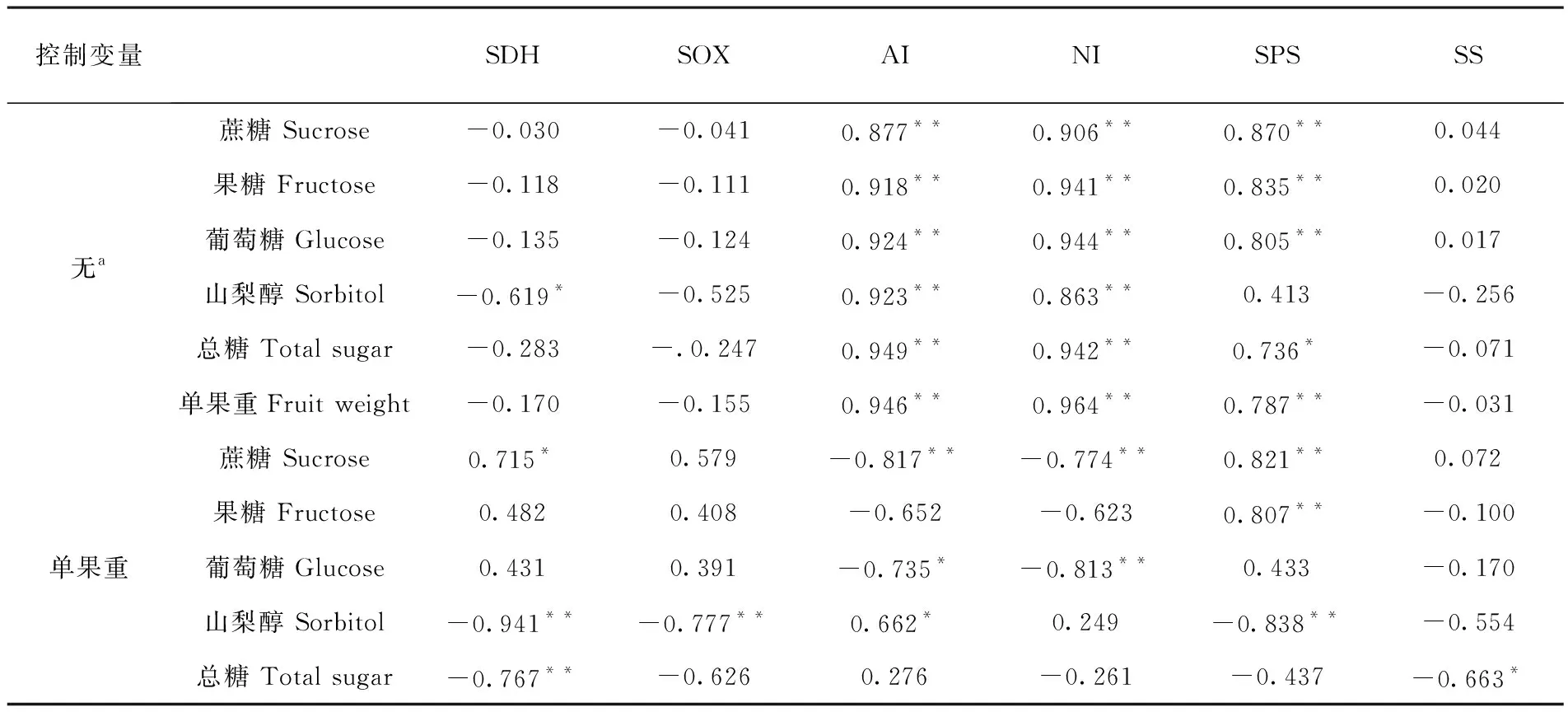
控制變量SDHSOXAINISPSSS無a蔗糖 Sucrose-0.030-0.0410.877**0.906**0.870**0.044果糖 Fructose-0.118-0.1110.918**0.941**0.835**0.020葡萄糖 Glucose-0.135-0.1240.924**0.944**0.805**0.017山梨醇 Sorbitol-0.619*-0.5250.923**0.863**0.413-0.256總糖 Total sugar-0.283-.0.2470.949**0.942**0.736*-0.071單果重Fruit weight-0.170-0.1550.946**0.964**0.787**-0.031單果重蔗糖 Sucrose0.715*0.579-0.817**-0.774**0.821**0.072果糖 Fructose0.4820.408-0.652-0.6230.807**-0.100葡萄糖 Glucose0.4310.391-0.735*-0.813**0.433-0.170山梨醇 Sorbitol-0.941**-0.777**0.662*0.249-0.838**-0.554總糖 Total sugar-0.767**-0.6260.276-0.261-0.437-0.663*
注:a. 單元格包含零階 (Pearson) 相關,顯著水平:P<0.05(*),P<0.01(**)
Note: Significant atP<0.05(*),P<0.01(**)
2.7 庫爾勒香梨果實發育過程中AI和NI活性變化
研究表明,香梨果實中AI和NI活性變化趨勢一致,且兩者活性水平沒有顯著差異,隨著果實發育而上升,pH變化均是在細胞分裂前期(花后 15~45 d)低且變化幅度較小,花后60 d起急劇上升,至花后90 d分別達到峰值5.69 μmol/(h·g)和5.50 μmol/(h·g)。在后期(從花后105 d至花后135 d)略微上升后即趨于平緩。香梨果實中的AI活性在整個發育期間約增高了4倍左右(從花后15 d的1.48 μmol/(h·g)增至花后135 d的5.85 μmol/(h·g)),NI活性在整個發育期間約增高了3倍左右(從花后15 d的2.24 μmol/(h·g)上升至花后135 d的5.78 μmol/(h·g))。相關性分析表明,AI酶活性與蔗糖和葡萄糖含量的變化顯著負相關(相關系數分別為:r= -0.817**和r= -0.735*),而與山梨醇含量變化顯著正相關(r=0.662*)。NI酶活性只與蔗糖和葡萄糖含量變化極顯著負相關(相關系數分別為:r= -0.774**和r= -0.813*)。圖6,表2
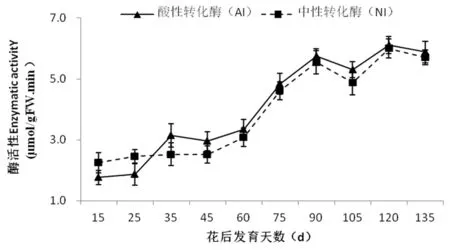
圖6 庫爾勒香梨果實發育過程中酸性轉化酶(AI)和中性轉化酶(NI)活性變化
Fig.6 Changes in AI and NI activities during the fruit development in Korla fragrant pear
3 討 論
果實品質在很大程度上取決于所含糖的種類和數量,而其數量取決于各種糖在果實發育過程中的消長結果。不同植物果實糖的組成與積累模式不盡相同。梨等薔薇科果樹果實糖積累特征為光合產物進入果實中,在果實發育的中期同化物轉換為淀粉進行積累,隨后至果實發育后期同化物以糖的形式直接進入,此時,由于淀粉開始水解使果實可溶性糖含量上升[20-21]。研究發現,庫爾勒香梨果實發育早中期以積累淀粉和山梨醇為主, 后期果糖、葡萄糖和蔗糖含量逐漸增多, 其中果糖含量上升最為突出并超過山梨醇成為果實中的主要糖,蔗糖含量最低,葡萄糖和山梨醇含量較接近,這與前人的研究結果基本一致[4,5,22]。果實發育早期,果糖、葡萄糖和蔗糖含量較低,可能是因為果實發育早期需要的能量和碳骨架增加,將輸入的光合產物一部分用于糖酵解和三羧酸循環及隨后的氨基酸和蛋白質合成,一部分轉化為淀粉進行積累[23],該時期香梨果實中的淀粉含量逐漸增高,達到一個峰值;至果實發育后期, 山梨醇含量變化平緩,而其余可溶性糖含量快速增長,一部分光合產物以糖的形式直接輸入, 同時淀粉也開始水解進入糖代謝, 果實中果糖、葡萄糖和蔗糖等可溶性糖含量上升。
山梨醇是梨的同化物運輸的主要形式,進入果實后,大部分經過SDH和SOX 的分解作用形成葡萄糖和果糖,未被分解的積累在果實中,是影響果實品質形成的重要組分。研究中香梨果實發育90 d左右可溶性糖以山梨醇為主,說明庫爾勒香梨幼果期可溶性糖以山梨醇為主,其次為果糖,葡萄糖和蔗糖含量較少;這與李馨玥等[24]在南果梨中的研究結果相同。幼果期山梨醇相關代謝酶的活性都較高這與許讓偉[10]的研究結論一致,這樣可以使運進的山梨醇快速地代謝以滿足果實發育初期細胞分裂和生長所需要的能量和中間物。隨著果實的發育SDH和SOX酶活性逐漸降低,花后60 d左右達到低谷,該時期果實中的山梨醇和淀粉含量迅速上升,淀粉含量達到最大值,而果糖和葡萄糖含量很低,變化平緩。大部分山梨醇不能轉化成果糖和葡萄糖;可能該時期果實的發育需要的能量和碳骨架較少,運進的糖只有一小部分用于糖酵解和三羧酸循環及隨后的氨基酸和蛋白質合成。于是果糖和葡萄糖開始在液泡中積累,淀粉合成上調,運進的糖以淀粉的形式貯存在質體中[23]。在整個果實發育階段,香梨果實SHD和SOX酶活性變化趨勢雖一致,但前者的變化幅度大,且其活性水平始終顯著高于后者,以 SDH 催化山梨醇生成果糖的能力較強,對果糖積累的貢獻可能相對更高[25]。這可能是香梨果實中果糖含量顯著高于葡萄糖含量的原因之一。另外,淀粉開始大量降解之前,葡萄糖主要來自蔗糖的水解。山梨醇轉運蛋白受到果糖和葡萄糖的競爭性抑制[26-27],為了避免山梨醇在質外體的過度積累,經過轉化酶水解的蔗糖應該是少量的。由此看來,果實發育早期,蔗糖的水解對葡萄糖的積累貢獻不大。試驗結果表明,香梨果實發育早期葡萄糖和蔗糖含量很低,分別占總可溶性糖的8.9%和6.2%,而山梨醇的含量占總糖中的比例50%以上,該時期NI和AI活性的較低,說明葡萄糖的再生減少。果實成熟時淀粉大量的降解,葡萄糖僅僅是微量的增加,可能與該時期蔗糖的再生有關,因為果實成熟時SPS活性大增,而SPS與蔗糖的積累有顯著關系。
SPS活性和蔗糖積累的趨勢相一致,在蔗糖代謝中起關鍵作用,而SS活性對蔗糖的積累作用較小[28]。研究發現,庫爾勒香梨果實發育前期蔗糖的變化幅度很少,到成熟期蔗糖含量快速上升,香梨具有在果實近成熟期積累蔗糖的特性。與其相應,果實發育前期SPS酶活性較低,隨著果實的長大逐漸上升,而果實發育早期SS活性高,之后逐漸下降,到后期又上升,SPS活性和蔗糖積累的趨勢相一致,這與常尚連的結論一致。香梨果實發育前期AI和NI的活性較低,隨著果實發育逐漸升高,這與陳露露等[29]在對翠冠梨的研究結果相反,這可能是香梨果實中蔗糖含量始終保持較低的原因。果實將要成熟時,蔗糖含量的持續上升是和SPS酶活性上調一致。淀粉降解之前,香梨果實中的蔗糖含量保持較低水平,這一方面,蔗糖可能是果實發育前期淀粉合成與積累的來源;其次,蔗糖作為代謝調控的信號,從而調控蔗糖/淀粉代謝轉化方向和強度[16, 30-34]。因此,香梨果實發育前期沒有蔗糖的明顯積累,可能與低含量的蔗糖僅作為單糖/多糖代謝之間的路徑連接信號作用有關。果實成熟階段蔗糖含量的增加主要是由于蔗糖的重新合成引起的。
4 結 論
庫爾勒香梨果實發育早中期以積累淀粉和山梨醇為主(占可溶性總糖中的比例50%以上), 后期果糖、葡萄糖和蔗糖含量逐漸增多, 其中果糖含量上升最為突出并超過山梨醇成為果實中的主要糖(占可溶性總糖中的比例38.7%)。果實發育過程中可溶性糖的積累主要受山梨醇的影響,受蔗糖、葡萄糖和果糖積累差異的影響較小。
參考文獻(References)
[1] 伍濤,陶書田,張虎平,等. 疏果對梨果實糖積累及葉片光合特性的影響[J]. 園藝學報,2011,38(11):2 041-2 048.
WU Tao,TAO Shu-tian,ZHANG Hu-ping,et al. (2011). Effects of Fruit Thinning on Fruit Sugar Accumulation and Leaf Photosynthetic Characteristics of Pear [J].ActaHorticulturaeSinica, 38(11): 2,041-2,048. (in Chinese)
[2] 黃艷花, 曾明. 梨果實糖代謝及調控因子的研究進展[J]. 植物生理學報,2013, 49(8): 709-714.
HUANG Yan-hua, ZENG Ming. (2013). Research advances in sugar metabolism and regulatory factors in pear fruits plant [J].PhysiologyJournal, 49(8): 709-714. (in Chinese)
[3] 霍月青. 砂梨品種資源糖酸及石細胞含量特點研究[D].武漢: 華中農業大學碩士學位論文,2007.
HUO Yue-qing. (2007).Studyoncharacteristicsofsugars,acidsandstonecellindifferentsandpearsfromgermplasmresources[D]. Master Dissertation: Huazhong Agricultural University, Wuhan.(in Chinese)
[4] 姚改芳,張紹鈴,曹玉芬,等. 不同栽培種梨果實中可溶性糖組分及含量特征[J]. 中國農業科學,2010,43(20): 4 229-4 237.
YAO Gai-fang, ZHANG Shao-ling, CAO Yu-fen, et al. (2010). Characteristics of components and contents of soluble sugars in pear fruits from different species [J].ActaHorticulturaeSinica, 43(20): 4,229-4,237. (in Chinese)
[5] 熊碧玲,汪志輝,劉燕. 鮮黃梨果實糖積累及山梨醇轉化相關酶活性的變化[J]. 中國南方果樹, 2011,40(4): 24-26.
XIONG Bi-ling,WANG Zhi-hui, LIU Yan. (2011). Changes in sugar accumulation and enzymatic activities of sorbitol metabolism in developing fruit of pyrus pyrifolia cv. Sunhwang [J].SouthChinaFruits, 40 (4): 24-26. (in Chinese)
[6] 崔艷波,陳 慧,樂文全,等. 京白梨與鴨梨正反交后代果實性狀遺傳傾向研究[J]. 園藝學報,2011,38(2):215-224.
CUI Yan-bo,CHEN Hui,YUE Wen-quan,et al. (2011). Studies on genetic tendency of fruit characters in reciprocal crosses generation between Jingbaili and Yalipear cultivars [J].ActaHorticulturaeSinica,38(2):215-224. (in Chinese)
[7] 楊志軍,樂文全,張紹鈴,等. 鴨梨×京白梨雜交后代果實可溶性糖積累差異以及相關酶活性研究[J]. 園藝學報,2012,39(6):1 055-1 063.
YANG Zhi-jun,YUE Wen-quan,ZHANG Shao-ling,et al. (2012). The Difference of Soluble Sugar Accumulation and Related Enzymes Activities in Pear Fruit from Hybrid Offspring [J].ActaHorticulturaeSinica, 39(6):1,055-1,063. (in Chinese)
[8] 李芳芳,何子順,陶書田,等. 套袋對庫爾勒香梨果實發育過程中可溶性糖含量的影響[J]. 果樹學報,2014,31(6): 1 072-1 078.
LI Fang-fang,HE Zi-shun,TAO Shu-tian,et al. (2014). Effects of bagging on soluble sugars contents during fruit development of Korla fragrant pear [J].JournalofFruitScience, 31(6): 1,072-1,078. (in Chinese)
[9] 王德孚,楊志軍,孫江妹,等,吳俊梨不同品種果實可溶性糖積累差異及代謝相關酶活性[J]. 果樹學報,2014,31(1):30-38.
WANG De -fu,YANG Zhi -jun,SUN Jiang -mei,et al. (2014). Difference in soluble sugar accumulation and related enzyme activity of fruits among different pear cultivars [J].JournalofFruitScience, 31(1): 30-38. (in Chinese)
[10] 許讓偉. 砂梨果實和葉片中糖積累及代謝相關酶活性變化研究[D]. 武漢: 華中農業大學碩士論文,2009.
XU Rang-wei. (2009).Studiesonchangesofsugaraccumulatiomandrelatedenzymesactivitiesinsandpearfruitsandleaves[D]. Master Dissertation: Huazhong Agricultural University, Wuhan.(in Chinese)
[11] 于年文,李俊才,王家振,等. 南國梨果實糖積累過程中相關酶的作用[J]. 西南農業學報,2011,24(3):919-923.
YU Nian-wen,LI Jun-cai,WANG Jia-zhen,et al. (2011). Roles of related enzymes in accumulation of sugars in Nanguoli pear fruit[J].SouthwestChinaJournal, 24(3): 919-923. (in Chinese)
[12] 劉松忠,劉軍,朱青青,等. 肥料種類對不同采收期黃金梨糖酸含量和風味的影響[J]. 果樹學報,2012,29(5): 804-808.
LIU Song-zhong,LIU Jun,ZHU Qing-qing,et al. (2012). Effects of manure types on sugar and acid contents and flavor of pear(Pyrus pyrifolia Hawangkumbae)at different mature stages [J].JournalofFruitScience, 29(5): 804-808.(in Chinese)
[13] Li, M., Feng, F., & Cheng, L. (2012). Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development.PlosOne, 7(3): e33055.
[14] Ito, A., Sakamoto, D., & Moriguchi, T. (2012). Carbohydrate metabolism and its possible roles in endodormancy transition in japanese pear.ScientiaHorticulturae, 144(3): 187-194.
[15] 戴美松,徐飛,施澤彬,等. 砂梨山梨醇轉運蛋白(SOT)基因家族成員表達特性及在果實糖積累中的作用初探[J]. 園藝學報,2015,42 (8):1 457-1 466.
DAI Mei-song,XU Fei,SHI Ze-bin,et al. (2015). Preliminary Study on Expression Characteristics of Sorbitol Transporter(SOT)Gene Family and the Role in Sugar Accumulation in Pyrus pyrifolia Fruits[J].ActaHorticulturaeSinica, 42 (8):1,457-1,466. (in Chinese)
[16] 潘儼,孟新濤,車鳳斌,等. 庫爾勒香梨果實發育成熟的糖代謝和呼吸代謝響應特征[J]. 中國農業科學,2016, 49(17):3 391-3 412.
PAN Yan, MENG Xin-tao, CHE Feng-bin, et al. (2016).Metabolic Profiles of Sugar Metabolism and Respiratory Metabolism of Korla Pear (Pyrus sinkiangensis Yu) Throughout Fruit Development and Ripening[J].ScientiaAgriculturaSinica, 49(17):3,391-3,412.(in Chinese)
[17] 袁暉,韋云,李馨玥,等. '南果梨'及其芽變'南紅梨'果實中糖分積累與相關基因表達差異分析[J]. 果樹學報, 2017,34(5):534-540.
YUAN Hui, WEI Yun, LI Xin-yue, et al. (2017). Differences in sugar accumulation and the related gene expression infruit development between'Nanguo'and its mutant'Nanhong'pears[J].JournalofFruitScience, 34(5): 534-540. (in Chinese)
[18] 趙世杰. 植物生理學實驗指導[M].北京:中國農業科技出版社,1998.
ZHAO Shi-jie (1998).Experimentalguidanceofplantphysiology[M]. Beijing: China Agricultural Science and Technology Press. (in Chinese)
[19] Yamaguchi, H. (1996). Changes in the amounts of the nad-dependent sorbitol dehydrogenase and its involvement in the development of apple fruit.AmericanSocietyforHorticulturalScience, 121(5): 848-852.
[20] 王永章, 張大鵬. '紅富士'蘋果果實蔗糖代謝與酸性轉化酶和蔗糖合酶關系的研究[J]. 園藝學報, 2001,28(3): 259-261.
WANG Yong-zhang, ZHANG Da-peng. (2001). Study on the relationships between acid invertase, sucrose synthase and sucrose metabolism in red Fuji apple fruit[J].ActaHorticulturaeSinica, 28(3): 259-261. (in Chinese)
[21] 魏建梅. 蘋果(MalusdomesticaBorkh.) 果實質地品質發育及采后調控的生理和分子基礎[D]. 楊凌: 西北農林科技大學博士論文,2009.
WEI Jian-mei. (2009).Studyonthephysiologicalandmelecularmechanismoffruittexturedevelopmentandpost-harvestregulationofapple(MalusdomesticaBorkh.) [D]. PhD Dissertation. Northwest A&F University, Yangling. (in Chinese)
[22] 柯凡君, 張虎平, 陶書田, 等. 套袋對梨果實發育過程中糖組分及其相關酶活性的影響[J]. 西北植物學報, 2011,31(7):1 422-1 427.
KE Fan-jun, ZHANG Hu-ping, TAO Shu-tian , et al. (2011). Sugar Component Contents and Metabol ism-related Enzyme Activities in Developing Pear Fruits after Bagging[J].ActaBot.Boreal.-Occident.Sin. ,31 (7):1,422-1,427. (in Chinese)
[23] Rolland, F., & Sheen, J. (2002). Sugar sensing and signaling in plants.PlantCell, 14(Suppl): s185.
[24] 李馨玥,李通,袁暉,等. 南果梨果實發育過程中糖分積累與相關基因表達分析[J]. 果樹學報,2016,33(增刊):59-64.
LI Xin-yue,LI Tong,YUAN Hui,et al. (2016). Sugar accumulation and expression of related genes in'Nanguo'pear fruit[J].JournalofFruitScience, 33(supplement):59-64. (in Chinese)
[25] 梁東. 蘋果山梨醇代謝相關基因的分子特性研究[D]. 楊凌: 西北農林科技大學博士論文,2010.
LIANG Dong (2010).Molecularcharacteristicsofgenesinvolvedinsorbitolmetabolisminapple[D] PhD Dissertation. Northwest A&F Universit, Yangling. (in Chinese)
[26] Gao, Z., Maurousset, L., Lemoine, R., Yoo, S. D., Van, N. S., & Loescher, W. (2003). Cloning, expression, and characterization of sorbitol transporters from developing sour cherry fruit and leaf sink tissues.PlantPhysiology, 131(4): 1,566-1,575.
[27] Zhang, H. P., Wu, J. Y., Tao, S. T., Wu, T., Qi, K. J., & Zhang, S. J., et al. (2014). Evidence for apoplasmic phloem unloading in pear fruit.PlantMolecularBiologyReporter, 32(4):931-939.
[28] 常尚連, 于賢昌, 于喜艷. 西瓜果實發育過程中糖分積累與相關酶活性的變化[J]. 西北農業學報,2006,15(3): 138-141.
CHANG Shang-lian, YU Xian-chang, YU Xi-yan. (2006). Change of sugar accumulation and related enzyme activity during fruit development of watermelon [J].ActaAgriculturaeBoreali-occidentalisSinica, 15(3): 138-141. (in Chinese)
[29] 陳露露,王濤,黃雪燕,等. 大棚翠冠梨果實糖積累及蔗糖代謝相關酶活性的研究[J]. 果樹學報, 2011,28(3): 400-405.
CHEN Lu -lu,WANG Tao,HUANG Xue -yan,et al. (2011). Study on enzyme activities involved in sugar accumulation and sucrose metabolization in Cuiguan pear under plastic tunnel culture [J].JournalofFruitScience, 28(3): 400-405. (in Chinese)
[30] 賈海鋒. 蔗糖及茉莉酸信號在草莓果實發育中的作用及其機理分析[D]. 北京: 中國農業大學博士論文,2013.
JIA Hai-feng. (2013).Rolesandmechanmismsofsucroseandjasmonicacidsignalsintheregulationofstrawberryfruitdevelopment[D]. PhD Dissertation. China Agricultural University, Beijing. (in Chinese)
[31] 陳俊偉, 張上隆, 張良誠. 果實中糖的運輸、代謝與積累及其調控[J]. 植物生理與分子生物學學報, 2004,30(1): 1-10.
CHEN Jun-wei, ZHANG Shang-long, ZHANG Liang-cheng. (2004). Sugar transport, metabolism, accumulation and their regulation in fruits [J].JournalofPlantPhysiologyandMolecularBiology, 30(1): 1-10. (in Chinese)
[32] Schultz J A, Juvik J A. (2004). Current models for starch synthesis and the sugary enhancer1 (se1) mutation in Zea mays[J].PlantPhysiolBiochem, 42(6):457-464.
[33] 張紅. 風味甜瓜果實酸味形成機理及糖酸遺傳和積累模擬研究[D]. 楊凌: 西北農林科技大學博士論文,2009.
ZHANG Hong.(2009).MechanismoforganicacidformationandgeneticsofsugarandacidandsimulationofAccumulationinflavormelonfruit[D]. PhD Dissertation. Northwest A&F University, Yangling. (in Chinese)
[34] 郝敬虹, 李天來, 孟思達, 等, 孫利萍夜間低溫對薄皮甜瓜果實糖積累及代謝相關酶活性的影響[J]. 中國農業科學, 2009,42(10): 3 592-3 599.
HAO Jing-hong, LI Tian-lai, MENG Si-da, et al. (2009). Effects of night low temperature on sugar accumulation and sugar-metabolizing enzyme activities in melon fruit [J].ScientiaAgticulturaSinica, 42(10): 3,592-3,599. (in Chinese)
