玉米幼苗鹽脅迫條件下的生理響應機制
2018-07-09 12:48:40陳勛基常曉春足木熱木吐爾遜李建平郝曉燕高升旗黃全生
新疆農業科學 2018年4期
陳勛基,陳 果,常曉春,足木熱木·吐爾遜,李建平,郝曉燕,高升旗,黃全生
(新疆農業科學院核技術生物技術研究所,烏魯木齊 830091)
0 引 言
【研究意義】鹽堿對農作物栽培和產量的影響是多方面的。鹽堿條件下,作物根系發育受到抑制,影響水分吸收,同時營養吸收及代謝造成阻滯,從而影響產量[1, 2]。玉米是全球最重要的糧食作物之一,鹽脅迫是限制玉米生產的重要非生物逆境脅迫[3]。耐鹽性狀非常復雜,由多個調控網絡組成,屬于數量性狀位點,在鹽脅迫條件下,不同材料對脅迫的應答和適應機制很大差異[4]。玉米種質資源中擁有豐富的耐鹽變異,不同種質資源對鹽脅迫的應答具有明顯差異[5],通過分析不同自交系和雜交種對鹽脅迫的應答模式,對研究玉米耐鹽機制以及遺傳改良有著重要作用。玉米自交系和雜交種耐鹽性鑒定對于選育耐鹽玉米新品種具有重要意義。【前人研究進展】植物在受到鹽脅迫后會表現出一些共同的應激反應。主要表現為細胞脫水,從而使細胞體積發生變化,細胞膜受損,細胞質外流到胞外空間[6]。細胞膨壓的變化會產生大量的活性氧,從而被質膜上的一些通道識別和感受,激活細胞活性氧的信號[7]。通常中度的滲透和鹽堿就可導致植物根、莖、葉生長阻滯。另外,鹽脅迫的劑量也決定了對植物生長抑制程度及對下游不同調節物質的響應水平[8]。植物在經歷鹽脅迫后,主要分為根部和地上部分對鹽堿的應答。植物根部在受到持續的干旱后,其根尖分生區向下伸長,促使其吸收更多水分[9];植物地上部分會迅速積累大量的可溶性物質,從而對其進行滲透調節[10]。同樣,在鹽脅迫條件下,植物通過增加根部表面積從而隔離對毒害離子的吸收[11]。在玉米自交系中也有大量對鹽脅迫較為耐受的材料如GEMS46和對鹽脅迫敏感的自交系如B73,大部分自交系屬于輕度耐鹽材料,不同學者對不同雜交種進行了耐鹽生理研究,篩選了大量耐鹽的雜交種[12]。【本研究切入點】對自交系耐鹽性雜種優勢研究還相對較少[13]。通過分析玉米自交系和雜交種對鹽脅迫條件下的應答模式,研究脅迫條件下玉米關鍵生理指標的變化趨勢,分析耐鹽脅迫條件下自交系及其雜交種耐鹽性,比較親本和雜交種對滲透脅迫和鹽離子毒害應答模式,確定穩定的耐鹽鑒定條件。【擬解決的關鍵問題】分析玉米自交系和雜交種在鹽脅迫條件下干重、鮮重、電導率變化和Na+、K+離子積累變化,研究典型自交系和雜交種對鹽脅迫的應答速率,自交系和雜交種離子毒害機制,為選育、鑒定耐鹽玉米自交系和雜交種選配提供理論支持。
1 材料與方法
1.1 材 料
玉米自交系B73和GEMS46種子為實驗室保藏,B73хGEMS46 F1(以下簡稱BхG)為實驗室配制的雜交種。成熟一致的玉米種子用10%次氯酸(v/v)鈉漩渦震蕩,表面消毒20 min,用無菌水沖6次后,將種子發芽紙上卷紙發芽,于28℃培養箱中暗培養48 h,選取長勢一致的幼苗播在裝有蛭石和營養土(1∶1)的花盆中,28℃,16 h 光照/8 h 黑暗條件下正常生長,隔3 d澆一次Hoagland營養液。
1.2 方 法
1.2.1 幼苗鹽脅迫處理
幼苗長到3葉1心期時,對照和處理分別開始進行營養液和鹽溶液處理。對照植株正常澆營養液,鹽處理植株每隔2 d澆灌150 mmol的NaCl溶液,每次澆完后倒去多余的液體,確保花盆底部不積水。分別在鹽處理開始第0、3、5和7 d,剪下地上部分莖葉用于測量各項生理指標。
1.2.2 幼苗Na+、K+離子含量測定
將幼苗從花盆中取出,清洗根部基質,置于85℃烘箱24 h,取出樣品稱重。將樣品置于消化液(濃硝酸∶高氯酸=4∶1)中,馬沸爐中175℃消化3 h。當液體變為清亮透明時,按照不同濃度稀釋樣品。稀釋后的樣品在原子吸收分光光度計上測定Na+、K+離子含量。
1.2.3 玉米葉片H2O2含量測定
稱取玉米葉片0.5 g,置于液氮中迅速打磨樣品,按材料與提取劑1∶1的比例加入4℃下預冷的丙酮,輕輕震蕩混勻,離心機上3 000 r/min,離心10 min,吸取上清即為樣品提取液。用移液管吸取樣品提取液1 mL,按表中體積加入5%硫酸鈦和濃氨水,待沉淀形成后3 000 r/min離心10 min,棄去上清液。沉淀用丙酮反復洗滌2~3次,去除植物色素。向洗滌后的沉淀中加入5 mL 2 mol/L的硫酸溶液,待完全溶解后,比色測定吸光值。植物組織中H2O2含量 (μmol/g Fw) =(C×Vt) / (FW×V1),式中:C:標準曲線上查得樣品中H2O2濃度(μmol);Vt:樣品提取液總體積(mL);V1:測定時用樣品提取液體積(mL);FW:植物組織鮮重(g)。
1.2.4 玉米葉片電導率測定
將幼苗葉片在去離子水中浸泡沖洗3次,再將幼苗置于20 mL去離子水中的離心管,封口浸泡8 h,用電導儀測定電導率,將離心管在沸水中煮沸20 min,待離心管降至室溫,再次測定總電導率。相對電導率計算公式為:相對電導率(RE)=滲出電導值/總電導值。
2 結果與分析
2.1 鹽脅迫條件下干鮮重變化
玉米自交系和雜交種在收到鹽脅迫處理之后,對脅迫表現出不同的耐受水平,前期主要表現為滲透脅迫,后期為離子毒害[14]。在高滲透脅迫下植株表現吸水困難,甚至失水。從植株鮮重變化可以看出在正常條件下,自交系B73和GEMS46鮮重隨著時間推移鮮重不斷增加,雜交種B×G鮮重增加量顯著高于自交系。在鹽脅迫處理下,雜交種和自交系鮮重都呈現下降趨勢,B73下降最明顯鮮重從3.94 g下降到2.92 g。雜交種B×G下降趨勢較小,從處理前的4.21 g下降到3.64 g,說明鹽脅迫處理后,造成滲透脅迫,B73作為敏感自交系對滲透脅迫應答比較明顯,而雜交種對滲透脅迫適應能力較兩個親本更強。圖1
同幼苗鮮重一樣,植株干重變化可以看出在正常條件下,自交系B73和GEMS46干重隨著時間推移鮮重不斷增加,雜交種B×G干重增加量顯著高于自交系。而在鹽脅迫處理下,B73下降最明顯,干重從1.88 g下降到1.13 g。雜交種B×G下降趨勢較小,從處理前的2.23 g下降到1.64 g,說明鹽脅迫處理后,幼苗生長發育受到阻滯,干物質積累減少,B×G雜交種由于表現出雜種優勢,干物質積累能力較強,表現出對鹽脅迫較強的適應能力。圖2
2.2 鹽脅迫條件下相對電導率變化
植株受到鹽脅迫后使質膜的選擇透性發生改變或喪失, 導致溶質外滲, 因而質膜的透性變化可顯示細胞膜結構和功能的受損程度。研究表明, 正常培養條件下,自交系和雜交種電導率變化趨勢不明顯,而在鹽脅迫條件下,隨著脅迫時間的延長, 自交系和雜交種相對電導率都呈現增長趨勢,B73電導率增加趨勢非常明顯,在處理第7 d后達到71%,GEMS46次之,雜交種增長趨勢相對平緩,處理第7 d相對電導率為48%。B73在鹽脅迫條件下,離子滲出較多,質膜受損嚴重,GEMS46離子滲出較輕,雜交種與自交系相比其離子滲出較低,從而表現出一定的耐鹽性。 圖3
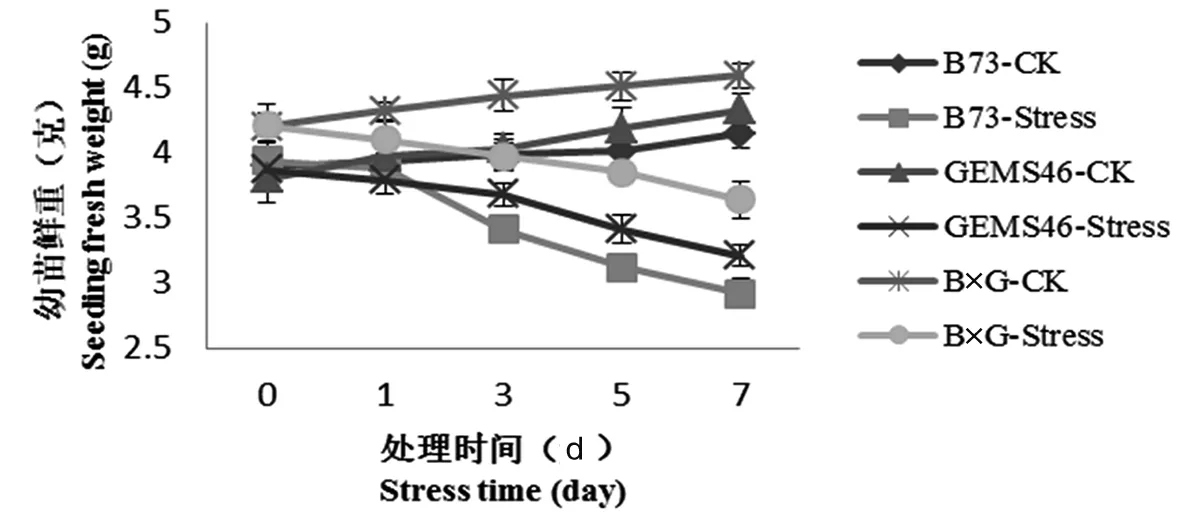
圖1 鹽脅迫處理后植株鮮重
Fig.1 The fresh weight of seeding under salt stress
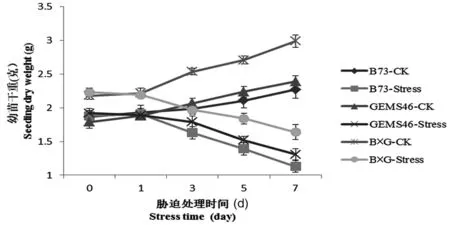
圖2 鹽脅迫處理后植株干重
Fig.2 Seedling dry weight after under salt stress
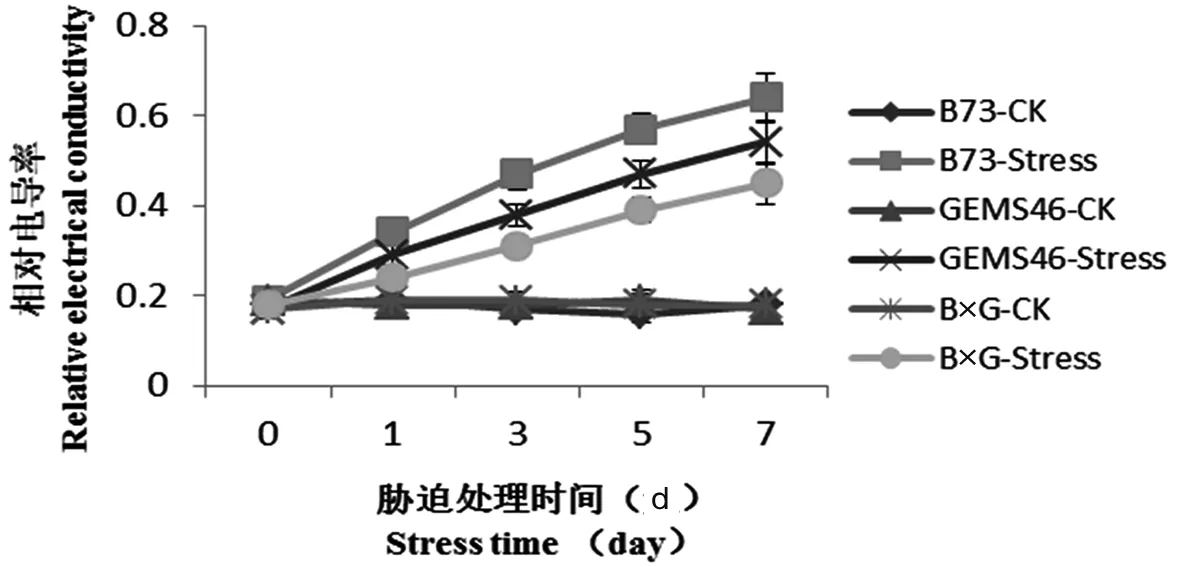
圖3 植株相對電導率
Fig.3 Seedling relative electrical conductivity
2.3 鹽脅迫條件下H2O2含量變化
自交系和雜交種經200 mmol NaCl脅迫后,葉片H2O2的含量變化明顯。研究表明,在正常條件下,B73、GEMS46和B×G的H2O2含量差異不顯著, 脅迫7 d后H2O2的含量變化明顯,B73幼苗脅迫7 d后H2O2含量是正常條件下的2.4倍,GEMS46幼苗脅迫7 d后H2O2含量達到10.03 μmol/g,雜交種脅迫7 d后,H2O2含量達到9.11 μmol/g,其增長趨勢較為平緩。在NaCl脅迫下,雜交種B×G和自交系GEMS46膜脂過氧化程度低于B73。自交系GEMS46在維持膜的功能具有突出優勢。圖4
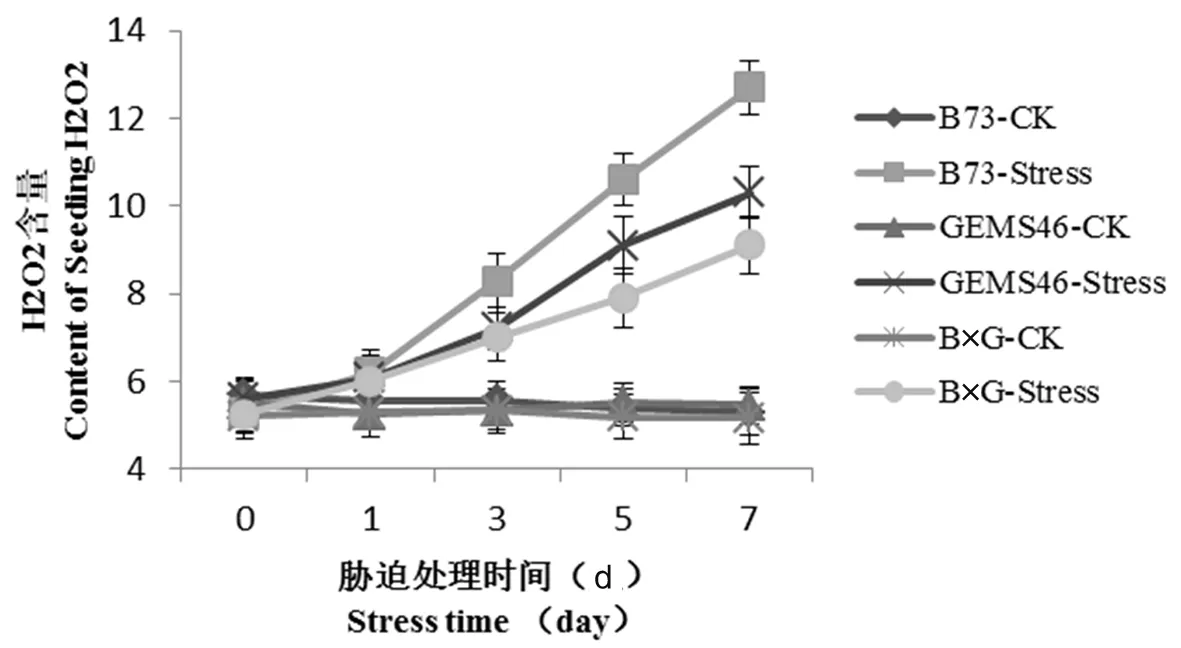
圖4 鹽脅迫后植株H2O2含量
2.4 鹽脅迫條件下Na+、K+離子含量變化
高濃度的Na+導致胞內K+、磷和有機溶質的外滲。于此同時,細胞K+/Na+比率下降,抑制胞質中的H+跨液泡膜運輸,導致液泡堿化,進一步阻礙Na+在液泡內積累。研究表明,鹽脅迫7 d后,自交系B73的Na+離子含量是正常對照的3.42倍,達到65 mg/g,雜交種B×G的Na+離子含量是正常對照的2.7倍,達到50 mg/g。在無鹽脅迫下,K+和Na+含量在自交系和雜交種之間無顯著差異。圖5
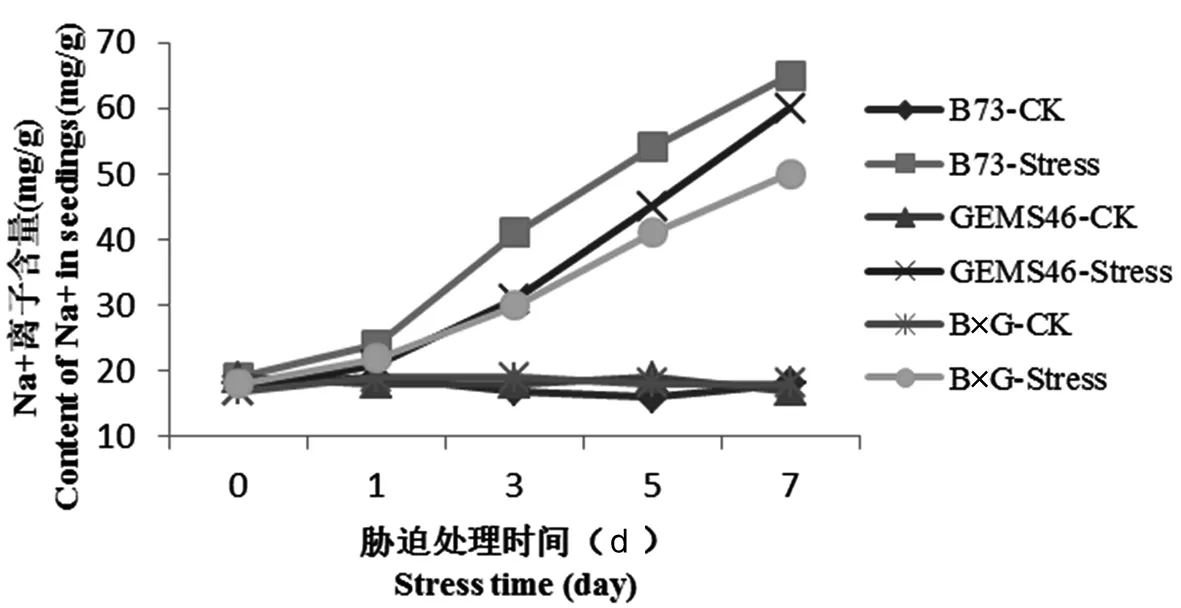
圖5 植株葉片Na+離子含量
脅迫處理7 d后,B73的K+離子含量從58 mg/g下降到31 mg/g,GEMS46從54 mg/g下降到36 mg/g,而雜交種在鹽脅迫下B×G的K+離子含量從55下降到39 mg/g,下降趨勢較自交系更為平緩。雜交種和自交系在Na+和K+含量以及K/Na比存在顯著差異。圖6
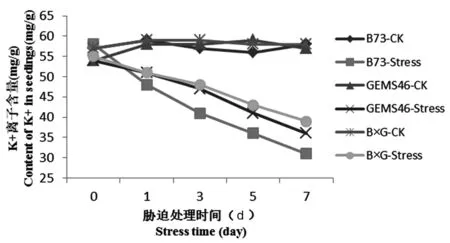
圖6 植株葉片K+離子含量
3 討 論
鹽脅迫可以導致水分虧缺,從而影響植物的代謝過程,這一過程是一個連續變化的過程[15]。在低鹽脅迫下,玉米幼苗鮮重和干重并沒有明顯的變化。當較高濃度NaCl持續脅迫處理后,幼苗植株生長量受到抑制,干物質積累減少[15-18]。試驗中,選用前期鑒定的鹽敏感自交系B73和相對耐鹽自交系GEMS46為親本,同時配制B×G雜交種進行比較,B73在150 mmol/L NaCl處理后第3 d就表現為鹽敏感,而GEMS46為相對耐鹽自交系,它及其雜交種在150 mmol/L NaCl 處理5~7 d表現出鹽離子毒害,不同品種在不同自交系和雜交種的耐鹽啟動機制及應對策略可能存在差異[18]。在玉米幼苗第15 d(3葉1心)時,生長發育迅速、生長勢較強,外源的非生物脅迫對其這一發育時期影響較大[19],由此可知在玉米3葉1心期是玉米對鹽分較為敏感時期,這一時期150 mmol/L NaCl即可造成鹽離子毒害。
玉米的耐鹽性是受多基因控制,能夠反映耐鹽性的生理指標和形態指標有很多[13, 20]。經過持續的鹽脅迫處理后,幼苗生長隨著脅迫處理加劇,植株膜系統受到損傷,導致離子外泄,而在植株體內Na+離子富集產生離子毒害并引起體內營養水平失衡而造成幼苗生長受到嚴重抑制[20]。同時,植株活性氧迅速增加,植株生長被完全抑制,造成嚴重的葉緣焦枯[21]。單一濃度或單一指標并不能對植物耐鹽性進行全面、有效的評價,綜合幾個濃度和幾個指標進行評價是必要的[22]。鹽處理后地上鮮重、地上干重、和Na+、K+離子含量的指標與玉米耐鹽性綜合評價結果顯著相關[23]。研究利用鮮重、干重、相對電導率、H2O2含量、Na+、K+離子含量作為衡量耐鹽性指標,對自交系和雜交種進行了耐鹽性綜合評價,為有效篩選玉米耐鹽性鑒定指標提供了依據。該方法具有操作性強、周期短、操作簡單、效率高等優點,且鑒定結果較準確,適宜大量材料的耐鹽性篩選。可作為玉米苗期耐鹽鑒定的首選指標,這與前人的研究結果一致[24]。
選用高耐鹽自交系為親本材料,通過輪回改良方式來改良敏感自交系的方法是可行的,研究中,B73是相對鹽敏感自交系,由于其對鹽離子脅迫響應不如高抗自交系,其體內Na+、K+離子積累較多,而雜交種和另一親本GEMS46對離子毒害響應較快,Na+、K+離子積累較少,說明其耐鹽性貢獻主要來自于耐鹽親本。自交系的耐鹽性強,則組配的玉米雜交種耐鹽性也較強,這與前人的研究結果基本一致[25]。近年來,許多玉米育種家對玉米耐鹽性進行了深入的研究,也取得了一定的進展。但是,我國玉米資源相對還比較匱乏,影響了耐鹽種質研究。隨著分子測序技術應用,越來越多的功能基因被發現,這將有利于從功能基因組方面對耐鹽性進行挖掘和利用。進而通過基因編輯手段應用到玉米遺傳改良中,這必將為高效耐鹽玉米的培育和玉米高產、穩產新品種選育提供理論基礎。
4 結 論
在3葉1心時用鹽處理玉米幼苗,連續處理第5 d后植株耐鹽性差異較為顯著,150 mmol/L NaCl處理植株是一個比較合理濃度;利用鹽脅迫后植株的鮮重和干重以及植株Na+、K+離子含量是衡量植株耐鹽性的較好的指標。在鹽處理后,耐鹽自交系的相對電導率、H2O2含量與雜交種變化趨勢相同,耐鹽自交系和雜交種對鹽脅迫的響應較快,植株體內積累Na+離子較小,K+/Na+比例變化范圍較小,說明雜交種的耐鹽性主要貢獻來自于耐鹽自交系。自交系的耐鹽性,選擇耐鹽自交系作為親本,配制雜交組合是選育耐鹽玉米新品種的必要方法。
參考文獻(References)
[1] 楊曉杰, 李旭業, 王海艷, 等. 玉米自交系耐鹽種質的篩選及耐鹽性評價[J]. 玉米科學, 2014,22(4):19-25.
YANG Xiao-jie, LI Xu-ye, WANG Hai-yan. (2014). Screening of Maize Inbred LIne Varieties with Salt Tolearance and the Evaluation [J].JournalofMaizeSciences, 22(4):19-25. (in Chinese)
[2]Dorothea Bartels, & Ramanjulu Sunkar. (2005). Drought and salt tolerance in plants.CriticalReviewsinPlantSciences, 24(1):23-58.
[3]馮艷, 周蓉, 付蓉, 等. 玉米遺傳多樣性分析及耐鹽種質篩選[J]. 江蘇農業科學, 2015,43(7):59-61.
FENG Yan, ZHOU Rong, FU Rong, et al. (2015). Genetic Diversity Analysis of Maize and screening of Salt-tolerant Germplasm (2015). [J].JiangsuAgriculturalSciences, 43(7):59-61. (in Chinese)
[4]雷震, 裴玉賀, 趙美愛, 等. 鹽脅迫下玉米葉片差異蛋白的雙向電泳分析[J]. 玉米科學, 2013, 21(3):82-86.
LEI Zhen, PEI Yu-he, ZHAO Mei-ai, et al. (2013). Analysis of soluble maize leaf proteins under salinity stress by two-demensional electronphoresis [J].JournalofMaizeSciences, 21(3):82-86. (in Chinese)
[5]Benke, A., Urbany, C., Marsian, J., Shi, R., Nv, W., & Stich, B. (2014). The genetic basis of natural variation for iron homeostasis in the maize ibm population.BmcPlantBiology, 14(1): 12.
[6]楊愛芳, 張可煒, 尹小燕, 等. 轉基因耐鹽玉米自交系的農藝性狀及雜種優勢表現的分析[J]. 中國農業科學, 2007,(12):2 895-2 902.
YANG Ai-fang, ZHANG Ke-wei, YIN Xiao-yan, et al. (2007). Agronomic Traits and Combining Ability Analysis of Salt Enduring Transgenic Maize [J].ScientiaAgriculturaSinica, (12):2,895-2,902. (in Chinese)
[7] Blumwald, E. (2000). Sodium transport and salt tolerance in plants.CurrentOpinioninCellBiology, 12(4): 431-434.
[8]Fujita, M., Fujita, Y., Noutoshi, Y., Takahashi, F., Narusaka, Y., & Yamaguchishinozaki, K., et al. (2006). Crosstalk between abiotic and biotic stress responses: a current view from the points of convergence in the stress signaling networks.CurrentOpinioninPlantBiology, 9(4): 436-442.
[9]李慧. 玉米鹽脅迫下組蛋白乙酰化介導的細胞壁相關基因的表達與功能[D]. 武漢大學, 2014.
Li Hui. (2014). Histone acetylation mediated expression of the cell wall related genes is involved in maize salt stress response[D].Wuhan University.(in Chinese)
[10]賈丹莉, 楊治平, 郭軍玲, 等. 6種玉米品種耐鹽性篩選[J]. 中國農學通報, 2017,33(11):1-8.
JIA Dan-li, YANG Zhi-ping, GUO Jun-ling, et al. (2017). Salt Tolerance Screening of Six Maize Varieties [J].ChineseAgriculturalScienceBulletin, 33(11):1-8. (in Chinese)
[11]李亮, 高明波, 于清濤. 耐鹽玉米自交系的鑒定[J]. 黑龍江農業科學, 2017,(2):14-17.
LI Liang, GAO Ming-bo, YU Qing-tao. (2017). Salt Tolerance Identification of Maize Inbred Lines [J].HelongjiangAgriculturalSciences, (2):14-17. (in Chinese)
[12]陳陽松. 玉米苗期耐鹽性全基因組關聯分析[D]. 北京:中國農業科學院碩士學位論文, 2017.
CHEN Yang-song. (2017).Commonwideassociationstudyofsalttoleranceatmaizeseedlingstage[D]. Master Dissertation. Chinese Academy of Agriculutlral Sciences, Beijing. (in Chinese)
[13]劉春榮, 張國新, 王秀萍. 主成分分析及隸屬函數法綜合評價玉米苗期耐鹽性[J]. 安徽農業科學, 2015,(28):13-14.
LIU Chun-rong, ZHANG Guo-xin, WANG Xiu-ping. (2015). Principal Component Analysis and Membership Function Method to Evaluate Salt Tolerance in Seedling Stage of Corn [J].JournalofAnhuiAgriculturalSciences, (28): 13-14. (in Chinese)
[14]張紅雪, 史振聲. 玉米耐鹽性研究進展[J]. 種子, 2015,34(10):47-51.
ZHANG Hong-xue, SHI Zhen-sheng. (2015). Research Progress of Maize Salt Tolerance [J].Seed, 34(10):47-51. (in Chinese)
[15]Farooq, M., Hussain, M., Wakeel, A., & Siddique, K. H. M. (2015). Salt stress in maize: effects, resistance mechanisms, and management. a review.AgronomyforSustainableDevelopment, 35(2):461-481.
[16]姚正培, 孟君, 李冠. 玉米自交系芽苗期耐鹽性的鑒定與篩選[J]. 華北農學報, 2007, (5):27-30.
YAO Zheng-pei, MENG Jun, LI Guan. (2007). Salinity Tolerance Identification and Screening of Maize Inbreeds in Seedling Emergence Stage [J].ActaAgriculturaeBoreali-Sinica, (5):27-30. (in Chinese)
[17]Galvanampudia, C. S., & Testerink, C. (2011). Salt stress signals shape the plant root.CurrentOpinioninPlantBiology, 14(3): 296-302.
[18]Zhu, J. K. (2002). Salt and drought stress signal transduction in plants.AnnualReviewofPlantBiology, 53(53): 247-273.
[19]Gruwel, M. L. H., Rauw, V. L., Loewen, M., & Abrams, S. R. (2001). Effects of sodium chloride on plant cells; a 31 p and 23 na nmr system to study salt tolerance.PlantScience, 160(5): 785-794.
[20]Golldack, D., Lüking, I., & Yang, O. (2011). Plant tolerance to drought and salinity: stress regulating transcription factors and their functional significance in the cellular transcriptional network.PlantCellReports,30(8):1,383-1,391.
[21]Shabala, S., & Cuin, T. A. (2008). Potassium transport and plant salt tolerance.PhysiologiaPlantarum, 133(4): 651-669.
[22]Agarie, S., & Cushman, J. C. (2007). Salt tolerance, salt accumulation, and ionic homeostasis in an epidermal bladder-cell-less mutant of the common ice plant mesembryanthemum crystallinum.JournalofExperimentalBotany, 58(8):1,957-1,967.
[23]劉學, 周璇, 曾興, 等. 玉米芽期和苗期耐鹽性鑒定方法的比較分析[J]. 玉米科學, 2015, 23(1):115-121.
LIU Xue, ZHOU Xuan, ZENG Xing, et al. 2015). Comparison of identification method of maize salt tolerance in Germination and seedling state [J].JournalofMaizeSciences, 23(1):115-121. (in Chinese)
[24]杜錦. 鹽脅迫下不同基因型玉米的生物學特性及其遺傳規律的研究[D]. 天津:天津農學院, 2010.
Du Jing.(2010).Study on Biological Characteristics and Genetic Regularity of Different Genotypes Maize(Zea mays L.) under salt stress[D].Tianjin Agricultural University. (in Chinese)
[25]商學芳. 不同基因型玉米對鹽脅迫的敏感性及耐鹽機理研究[D]. 濟南:山東農業大學, 2007.
Shang Xue-fang. (2007). Study on Sensitivity of Salt Stress and Salt-Tolerant Mechanism of Different Genotypes Maize (Zea mays L.). Shangdong Agricultural University. (in Chinese)
