以魔芋多糖為碳源的產(chǎn)聚羥基丁酸酯菌的篩選、鑒定及發(fā)酵研究
2018-07-06 12:13:32李凌凌張家銘左振宇楊忠華
武漢科技大學(xué)學(xué)報(bào) 2018年4期
關(guān)鍵詞:生長(zhǎng)
李凌凌,毛 偉,張家銘,黃 皓,左振宇,楊忠華
(1.武漢科技大學(xué)化學(xué)與化工學(xué)院,湖北 武漢,430081;2. 武漢科技大學(xué)煤轉(zhuǎn)化與新型炭材料湖北省重點(diǎn)實(shí)驗(yàn)室,湖北 武漢,430081)
魔芋多糖(又稱(chēng)魔芋葡甘露聚糖)的主要成分是甘露糖和葡萄糖以β-1,4-糖苷鍵連接起來(lái)的線(xiàn)性高分子化合物[1],可作為底物誘導(dǎo)微生物生產(chǎn)β-1,4-D-甘露聚糖酶(即β-甘露聚糖酶)[2],并在酶的作用下分解成能被人體吸收利用的魔芋低聚糖。魔芋低聚糖能有效改善腸道內(nèi)菌群結(jié)構(gòu),還可用于治療高血脂癥、預(yù)防動(dòng)脈硬化和冠心病的發(fā)生[3-4]。考慮到β-甘露聚糖酶為胞外水解酶,以魔芋多糖為碳源培養(yǎng)的微生物發(fā)酵液經(jīng)離心處理獲得含魔芋低聚糖的上清液后,會(huì)產(chǎn)生大量的菌體沉淀,如何利用這些菌體廢棄物開(kāi)發(fā)具有應(yīng)用前景的產(chǎn)品是一個(gè)重要的研究課題。
聚羥基烷酸酯(PHAs)作為一類(lèi)由微生物合成的細(xì)胞內(nèi)聚酯,是胞內(nèi)積累的碳源和能源儲(chǔ)存物,其典型代表為聚羥基丁酸酯(PHBs)。PHB具有生物降解性、熱塑性和生物相容性,并且無(wú)毒,被認(rèn)為是石油化學(xué)聚合物的替代品[5-6]。但是,與石油化工材料相比,利用微生物發(fā)酵法生產(chǎn)PHB的成本很高,嚴(yán)重制約了其大規(guī)模生產(chǎn)和應(yīng)用。研究表明,采用廉價(jià)碳源(如食品廢棄物[7]、粗甘油[8-9]、半纖維素水解液[9]、廢菜籽油[10]、皂化廢棕櫚油[11]等)培養(yǎng)微生物,可以使PHB的生產(chǎn)成本降低。目前已報(bào)道的能夠積累PHB的微生物種類(lèi)雖然不少,但仍然有許多潛在的聚羥基丁酸酯生產(chǎn)菌未被發(fā)現(xiàn),另外,不同微生物積累PHB所需的碳源也不盡相同[7,9],因此,很多學(xué)者一直致力于從各種環(huán)境中分離獲得可以利用廉價(jià)碳源合成PHB的新型細(xì)菌,從而將微生物發(fā)酵法應(yīng)用到PHB工業(yè)化生產(chǎn)中。
我國(guó)的花魔芋和白魔芋資源豐富[1],可以為微生物發(fā)酵生產(chǎn)PHB提供大量廉價(jià)的魔芋多糖作為碳源。本研究擬篩選以魔芋多糖為碳源在細(xì)胞內(nèi)合成聚羥基丁酸酯的菌株,達(dá)到降低PHB生產(chǎn)成本的目的,并且同時(shí)獲得魔芋低聚糖和生物降解材料PHB兩種高附加值產(chǎn)品。
1 試驗(yàn)材料與方法
1.1 材料
1.1.1 培養(yǎng)基
富集培養(yǎng)基:魔芋精粉2.0 g,酵母膏0.5 g,蛋白胨0.3 g,NaCl 0.5 g,加蒸餾水定容至100 mL,用于富集培養(yǎng)可產(chǎn)β-甘露聚糖酶的細(xì)菌。
固體分離培養(yǎng)基:魔芋精粉1.0g,NaNO30.3g,K2HPO40.1g,KCl 0.05g,MgSO40.05g,F(xiàn)eSO40.05 g,瓊脂1.8 g,加蒸餾水定容至100 mL,用于分離純化可產(chǎn)β-甘露聚糖酶的細(xì)菌。
LB培養(yǎng)基:按照文獻(xiàn)[12]配制,用于觀(guān)察細(xì)菌的菌落特征、液體培養(yǎng)特征、運(yùn)動(dòng)性及與氧氣的關(guān)系。
修正的M9培養(yǎng)基:Na2HPO40.678 g,KH2PO40.3 g,NaCl 0.05 g,NH4Cl 1.8 g,CaCl20.001 g,MgSO4·7H2O 0.0492 g,葡萄糖0.786 g,加蒸餾水定容至100 mL,pH調(diào)至7.0,用于優(yōu)化菌體的聚羥基丁酸酯生產(chǎn)條件。
1.1.2 酶和化學(xué)試劑
pMD18-T載體連接試劑盒為大連寶生物工程有限公司產(chǎn)品;通用型柱式基因組DNA提取試劑盒、高純度質(zhì)粒小提試劑盒、大腸桿菌DH5α感受態(tài)細(xì)胞和2×ES Taq MasterMix(含染料)為康為世紀(jì)生物科技有限公司產(chǎn)品;PCR產(chǎn)物純化回收試劑盒(Cycle-pure kit)為美國(guó)OMEGA公司產(chǎn)品;引物合成和測(cè)序工作由武漢擎科創(chuàng)新生物科技有限公司完成。
1.2 方法
1.2.1 以魔芋多糖為碳源合成PHB的菌株篩選
(1)菌株的富集及分離純化
取云南種植魔芋的農(nóng)田土壤樣品1 g加到100 mL富集培養(yǎng)基中,在30 ℃、150 r/min的恒溫?fù)u床中培養(yǎng)24 h。取1 mL富集培養(yǎng)的菌液稀釋涂布到固體分離培養(yǎng)基上,30 ℃培養(yǎng)24 h。挑取菌落在固體分離培養(yǎng)基上劃線(xiàn),待菌苔生成后鏡檢,如果菌體不純,則繼續(xù)劃線(xiàn)純化培養(yǎng),直至鏡檢下出現(xiàn)單一形態(tài)的菌體。
(2)菌株的篩選
挑取純化后的單菌落點(diǎn)接到固體分離培養(yǎng)基上,待菌落生成后,用剛果紅溶液染色0.5 h,再用生理鹽水洗去剛果紅染液,觀(guān)察菌落周?chē)袩o(wú)透明圈,作為菌株產(chǎn)β-甘露聚糖酶能力的初步檢測(cè)標(biāo)準(zhǔn)[13]。選擇透明圈大的菌株,檢測(cè)其合成PHB的能力:細(xì)菌經(jīng)蘇丹黑染色后在光學(xué)顯微鏡下觀(guān)察,若視野中60%的細(xì)菌內(nèi)有藍(lán)黑色顆粒,則為產(chǎn)PHB的菌株[14],命名為L(zhǎng)KH。
1.2.2 以魔芋多糖為碳源合成PHB的菌株鑒定
(1)菌株個(gè)體形態(tài)觀(guān)察和生理生化特征鑒定
革蘭氏染色及芽孢染色、細(xì)菌的生理生化特征鑒定均參照文獻(xiàn)[12]。
(2)菌株最適生長(zhǎng)溫度和pH值的確定
為了研究溫度對(duì)菌株生長(zhǎng)的影響,以10%的接種量接種處于對(duì)數(shù)期的菌株LKH到一組修正的M9液體培養(yǎng)基中,分別置于25、28、30、37、40 ℃的恒溫?fù)u床(200 r/min)中振蕩培養(yǎng)24 h,檢測(cè)培養(yǎng)液的OD600值。另外,以10%的接種量接種處于對(duì)數(shù)期的菌株到一組pH值分別為4.0、5.0、6.0、7.0、8.0的修正的M9液體培養(yǎng)基中,置于30 ℃恒溫?fù)u床(200 r/min)中振蕩培養(yǎng)24 h,檢測(cè)培養(yǎng)液的OD600值,以研究培養(yǎng)基的pH值對(duì)菌株生長(zhǎng)的影響。
(3)系統(tǒng)發(fā)育樹(shù)的構(gòu)建
按照通用型柱式基因組DNA提取試劑盒說(shuō)明書(shū)提取菌株LKH的基因組DNA。使用正向引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和反向引物1492R(5′-GGTTACCCTTGTTACGACTT-3′)進(jìn)行PCR擴(kuò)增。PCR反應(yīng)體系:菌體LKH的基因組DNA 1 μL,2×ES Taq MasterMix(含染料)12.5 μL,引物27F(20 μmol/L)1 μL,引物1492R(20 μmol/L)1 μL,無(wú)菌水補(bǔ)至25 μL。PCR擴(kuò)增條件:95 ℃預(yù)變性5 min,之后按如下程序(95 ℃變性1 min,58 ℃退火45 s,72 ℃延伸90 s)進(jìn)行35個(gè)循環(huán),最后一個(gè)循環(huán)72 ℃延伸10 min。根據(jù)pMD18-T載體連接試劑盒說(shuō)明書(shū),將菌株LKH的16S rDNA的PCR擴(kuò)增產(chǎn)物連接到pMD18-T載體上,構(gòu)建重組載體pMD-LKH。重組質(zhì)粒的測(cè)序結(jié)果提交到NCBI數(shù)據(jù)庫(kù)進(jìn)行Blast比對(duì),利用EBI數(shù)據(jù)庫(kù)中的Clustal Omega功能進(jìn)行多重序列比對(duì),使用軟件MEGA 5.0構(gòu)建系統(tǒng)發(fā)育樹(shù),其中根據(jù)maximum composite likelihood model計(jì)算進(jìn)化距離,采用neighbor-joining statistical method構(gòu)建進(jìn)化樹(shù)。
1.2.3 生長(zhǎng)曲線(xiàn)的繪制
取1 mL處于對(duì)數(shù)期的菌株接種到100 mL修正的M9液體培養(yǎng)基中,在30 ℃、200 r/min的恒溫?fù)u床中振蕩培養(yǎng)。每隔一定時(shí)間取樣,測(cè)定菌液的OD600值。試驗(yàn)設(shè)置3個(gè)平行組,取平均值繪制菌液的OD600隨時(shí)間的變化曲線(xiàn)(即生長(zhǎng)曲線(xiàn))。
1.2.4 PHB發(fā)酵條件的優(yōu)化
(1)PHB的提取及分析方法
采用氯仿-次氯酸鹽法提取PHB[15],參照文獻(xiàn)[16]計(jì)算PHB的含量和提取率。將PHB標(biāo)準(zhǔn)樣品和從菌株LKH中提取并純化的白色薄膜樣品經(jīng)甲酯化后進(jìn)行氣相色譜分析[16],同時(shí)兩類(lèi)樣品用KBr壓片后進(jìn)行紅外光譜檢測(cè)[17]。
(2)碳源的優(yōu)化
取1 mL處于對(duì)數(shù)期的菌株接種到100 mL添加相同質(zhì)量碳源(分別為花生油、亞油酸、亞麻酸、油酸、纖維素、魔芋精粉、葡萄糖)的修正的M9液體培養(yǎng)基中,在30 ℃、200 r/min的恒溫?fù)u床中振蕩培養(yǎng)4天,提取PHB,測(cè)定其含量(即PHB的純度),計(jì)算PHB提取率。
(3)氮源濃度的優(yōu)化
以魔芋精粉為碳源物質(zhì),配制分別含有0.1、0.2、1、5、10、20 g/L氮源NH4Cl的修正的M9液體培養(yǎng)基。取1 mL處于對(duì)數(shù)期的菌株接種到100 mL上述培養(yǎng)基中,在30 ℃、200 r/min的恒溫?fù)u床中振蕩培養(yǎng)4天,提取PHB,測(cè)定其含量,計(jì)算PHB提取率。
2 結(jié)果與分析
2.1 產(chǎn)聚羥基丁酸酯菌LKH的篩選
挑取固體分離培養(yǎng)基上剛果紅染色水解圈較大的菌株(見(jiàn)圖1(a))進(jìn)行蘇丹黑染色(見(jiàn)圖1(b)),結(jié)果呈陽(yáng)性,且胞內(nèi)的藍(lán)黑色顆粒較多,初步推斷該菌能夠分泌β-甘露聚糖酶,水解魔芋精粉以提供生長(zhǎng)所需的碳源,同時(shí)在胞內(nèi)合成PHB。

(a)剛果紅染色的水解圈
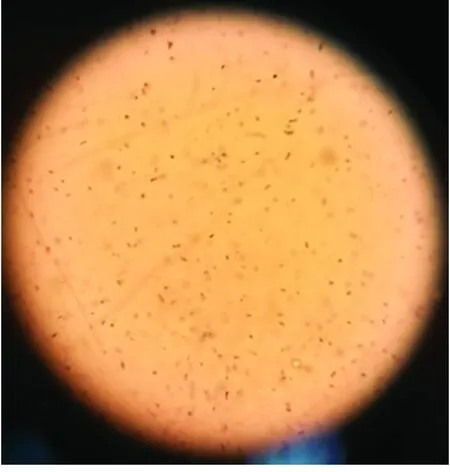
(b)蘇丹黑染色
2.2 產(chǎn)聚羥基丁酸酯菌LKH的鑒定
2.2.1 菌株LKH的個(gè)體形態(tài)特征
在光學(xué)顯微鏡下,菌株LKH呈桿狀,可游動(dòng)、旋轉(zhuǎn),革蘭氏染色為紫色,呈陽(yáng)性,芽孢染色為綠色,呈陽(yáng)性,芽孢形態(tài)為橢圓形。
2.2.2 菌株LKH的群體形態(tài)特征
菌株LKH在LB固體培養(yǎng)基上生長(zhǎng)24 h后長(zhǎng)出的菌落為乳白色,圓形,不透明,表面光滑濕潤(rùn),黏度較大,邊緣整齊且中間隆起(見(jiàn)圖2a)。菌株LKH在LB半固體培養(yǎng)基的穿刺試驗(yàn)結(jié)果如圖2(b)所示,菌體在穿刺部位的四周呈云霧狀生長(zhǎng),推斷菌株LKH兼性好氧,可運(yùn)動(dòng)。
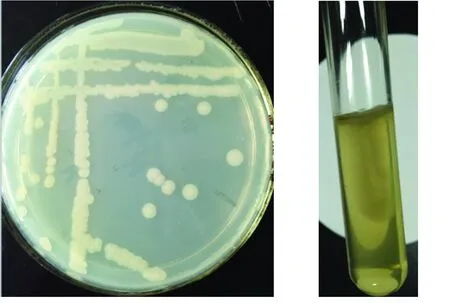
(a)菌落特征 (b)穿刺試驗(yàn)結(jié)果
圖2菌株LKH的形態(tài)特征
Fig.2MorphologicalcharacteristicsofStrainLKH
2.2.3 菌株LKH的最適生長(zhǎng)溫度和pH值
溫度和培養(yǎng)基pH值對(duì)菌株LKH生長(zhǎng)的影響如圖3所示。由圖可見(jiàn),在25~40 ℃的溫度范圍內(nèi),菌株LKH在修正的M9液體培養(yǎng)基中都可以較好地生長(zhǎng),其中在28 ℃下生長(zhǎng)得最好。另外,當(dāng)培養(yǎng)基pH值低至4.0時(shí),菌株LKH不能正常生長(zhǎng),而在pH 值為6.0~8.0的范圍內(nèi),菌株在修正的M9液體培養(yǎng)基中都可以很好地生長(zhǎng),其中菌株LKH的最適生長(zhǎng)pH值約為7.0,此時(shí)生物量最大。
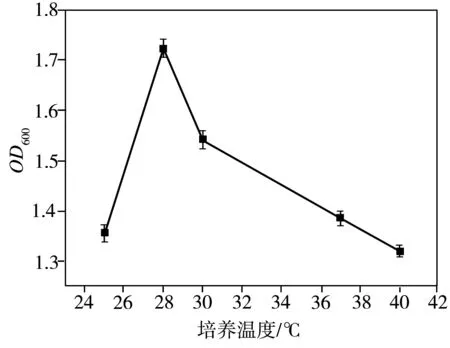
(a)溫度
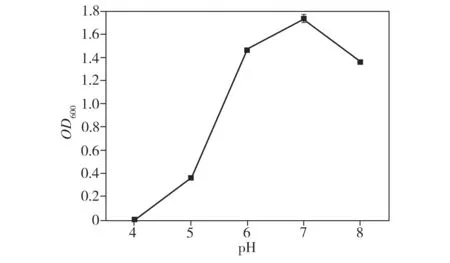
(b)培養(yǎng)基pH值
Fig.3EffectsoftemperatureandpHofculturemediumonthegrowthofStrainLKH
2.2.4 菌株LKH的生理生化特征
菌株LKH的生理生化特征如表1所示,為了便于后續(xù)的對(duì)比分析,表1中同時(shí)列出了菌株NRRL NRS-1356[18]和菌株M17[19]的生理生化特征。由表1可見(jiàn),菌株LKH的接觸酶反應(yīng)為陽(yáng)性,氧化酶反應(yīng)為陰性,可以向胞外分泌淀粉酶、脂肪酶和脲酶,能夠液化明膠。菌株LKH的IMViC試驗(yàn)結(jié)果分別為:Voges-Proskauer(V-P)反應(yīng)為陽(yáng)性,即細(xì)菌可以發(fā)酵葡萄糖生成乙酰甲基甲醇;檸檬酸試驗(yàn)呈陽(yáng)性反應(yīng),即可以利用檸檬酸鹽產(chǎn)生堿性碳酸鹽,使培養(yǎng)基pH值升高從而變成藍(lán)色;但菌株LKH的甲基紅反應(yīng)呈陰性,且不可產(chǎn)生色氨酸酶分解色氨酸產(chǎn)生吲哚。菌株LKH不能水解培養(yǎng)基中的含硫氨基酸生成H2S,也不能將硝酸鹽還原為亞硝酸鹽。在糖發(fā)酵試驗(yàn)中,菌株LKH可利用木糖、葡萄糖、半乳糖和麥芽糖產(chǎn)酸和少量的氣,而利用乳糖時(shí)不能產(chǎn)酸和氣。另外,菌株LKH可以在分別以葡萄糖、乳糖、魔芋精粉、纖維素、油酸、亞油酸、亞麻酸及花生油為唯一碳源的M9培養(yǎng)基上生長(zhǎng),但不能以正辛酸或正癸酸作為唯一碳源生長(zhǎng)。菌株LKH可以在40 ℃生長(zhǎng),但不能在4 ℃生長(zhǎng),其最適生長(zhǎng)溫度為28 ℃,最適生長(zhǎng)pH值為7.0。

表1 菌株LKH的生理生化特征
注:+代表陽(yáng)性結(jié)果;-代表陰性結(jié)果;N代表未知。
2.2.5 系統(tǒng)發(fā)育樹(shù)
對(duì)菌株LKH的16S rDNA的PCR擴(kuò)增產(chǎn)物進(jìn)行1.0%的瓊脂糖凝膠電泳檢測(cè),結(jié)果如圖4(a)所示,可見(jiàn)PCR擴(kuò)增產(chǎn)物在1500 bp左右有清晰條帶。將PCR擴(kuò)增產(chǎn)物切膠回收后,連接到pMD18-T質(zhì)粒上,構(gòu)建重組載體pMD-LKH,其PCR鑒定結(jié)果見(jiàn)圖4(b)。將重組載體送交測(cè)序,獲得一段長(zhǎng)1449 bp的核苷酸序列,利用NCBI數(shù)據(jù)庫(kù)提供的BlastN功能進(jìn)行核苷酸比對(duì),結(jié)果顯示菌株LKH與類(lèi)芽孢桿菌屬的依利諾斯類(lèi)芽孢桿菌(Paenibacillusillinoisensis)的親緣關(guān)系最接近,其中與P.illinoisensisF24菌(登錄號(hào)JQ579623.1)、NBRC 15959(登錄號(hào)NR_113828.1)、JCM 9907(登錄號(hào)NR_040884.1)的序列一致性高達(dá)99%。根據(jù)16S rDNA序列的相似性,利用Clustal Omega和Mega 5.0軟件構(gòu)建系統(tǒng)發(fā)育樹(shù),如圖5所示,圖中括號(hào)內(nèi)為基因序列的登錄號(hào)。由圖可見(jiàn),菌株LKH與P.illinoisensis的進(jìn)化關(guān)系最近。
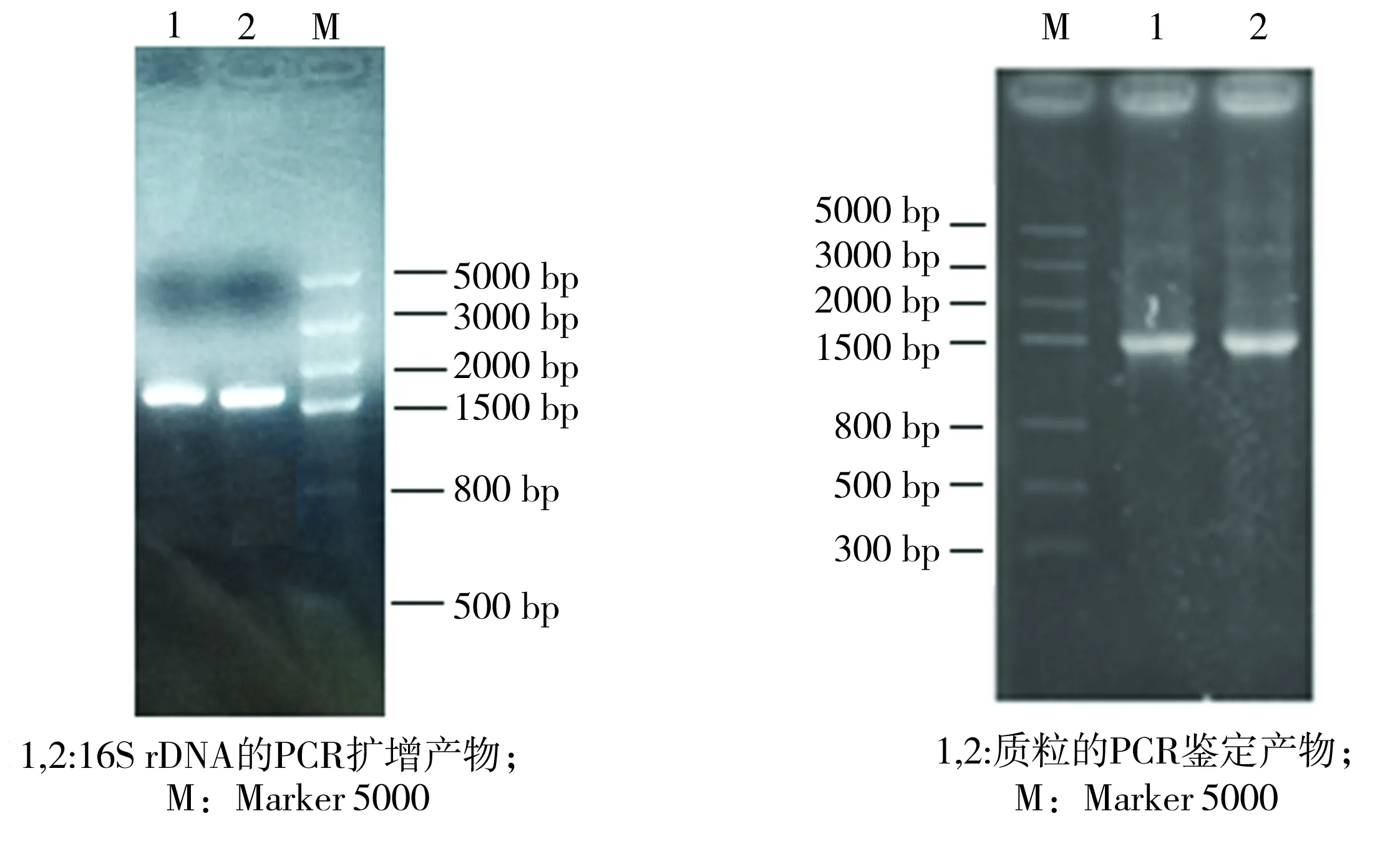
(a)16S rDNA基因的PCR擴(kuò)增產(chǎn)物 (b)質(zhì)粒pMD-LKH的PCR鑒定產(chǎn)物
圖4PCR產(chǎn)物的瓊脂糖凝膠電泳圖
Fig.4ElectrophoretogramofPCRproducts

圖5 根據(jù)16S rDNA序列的同源性構(gòu)建的系統(tǒng)發(fā)育樹(shù)
綜上所述,菌株LKH歸屬于依利諾斯類(lèi)芽孢桿菌P.illinoisensis。P.illinoisensis是1997年由Shida等從Bacilluscirculans的第6組中抽出來(lái)歸到類(lèi)芽孢桿菌屬[18]。P.illinoisensis的細(xì)胞是桿狀,大小為(0.5~0.8)μm×(3.0~5.0)μm,革蘭氏染色陽(yáng)性,周生鞭毛,可運(yùn)動(dòng),產(chǎn)橢圓形芽孢。從個(gè)體和菌落的形態(tài)及生理生化特征(見(jiàn)表1)上看,菌株LKH與依利諾斯類(lèi)芽孢桿菌及M17菌都很接近,但是這3種菌株的某些生理生化特征存在著一定的差異,譬如:菌株LKH的最適生長(zhǎng)溫度為28 ℃,V-P反應(yīng)為陽(yáng)性,能夠利用檸檬酸鹽,脲酶反應(yīng)為陽(yáng)性,吲哚反應(yīng)和硝酸鹽還原試驗(yàn)均為陰性,而菌株NRRL NRS-1356的最適生長(zhǎng)溫度為37 ℃,V-P反應(yīng)為陰性,不能利用檸檬酸鹽,菌株M17的脲酶反應(yīng)和V-P反應(yīng)為陰性,硝酸鹽還原試驗(yàn)和吲哚反應(yīng)均為陽(yáng)性。生理生化特征的差異反映了這些菌株的生存環(huán)境和自身遺傳背景的不同。
目前發(fā)現(xiàn)的可以在胞內(nèi)積累PHAs的微生物種類(lèi)很多,主要有產(chǎn)堿桿菌屬(Alcaligenes)、芽孢桿菌屬(Bacillus)、紅螺菌屬(Rhodospiriclum)、甲基營(yíng)養(yǎng)菌屬(Methylotrophs)、假單胞菌屬(Pseudomonas)、固氮菌屬(Azotobacter)、棒狀桿菌屬(Corynebacterium)、氣單胞菌屬(Aeromonas)、貪銅菌屬(Cupriavidus)、鏈霉菌屬(Streptomyces)、極端嗜鹽桿菌(Haloferax)等[5,17,20-21]。類(lèi)芽孢桿菌屬是一類(lèi)廣泛存在于自然界的好氧/兼性厭氧、可產(chǎn)生芽孢的桿狀細(xì)菌,在生物醫(yī)藥、生物修復(fù)等領(lǐng)域都有重要的應(yīng)用[22]。雖然迄今為止鮮有類(lèi)芽孢桿菌屬胞內(nèi)積累PHBs的報(bào)道,但是,考慮到多數(shù)類(lèi)芽孢桿菌屬的菌種是從芽孢桿菌屬抽出重新分類(lèi)的,相信今后有關(guān)類(lèi)芽孢桿菌屬中提取出PHBs的報(bào)道會(huì)層出不窮。
2.3 菌株LKH的生長(zhǎng)曲線(xiàn)
根據(jù)不同時(shí)間取樣的菌株LKH懸浮液的OD600繪制生長(zhǎng)曲線(xiàn),如圖6所示。菌株LKH在0~4 h內(nèi)處于生長(zhǎng)延滯期,生長(zhǎng)較為緩慢;4~24 h之間菌體進(jìn)入對(duì)數(shù)生長(zhǎng)期;24 h后,菌株開(kāi)始達(dá)到生長(zhǎng)穩(wěn)定期,此時(shí)生物量達(dá)到最大值;培養(yǎng)至96 h后,菌株開(kāi)始進(jìn)入衰亡期。由于PHB是微生物在非平衡的營(yíng)養(yǎng)條件下進(jìn)行胞內(nèi)合成的,故定于第4天提取PHB。
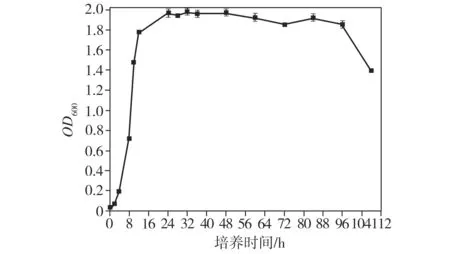
圖6 菌株LKH的生長(zhǎng)曲線(xiàn)
2.4 菌株LKH的PHB發(fā)酵條件優(yōu)化
2.4.1 碳源種類(lèi)對(duì)PHB合成的影響
不同碳源培養(yǎng)的菌株LKH中提取的PHB含量及提取率如表2所示。從表2中可以看出,菌株LKH在以葡萄糖為唯一碳源的條件下提取獲得PHB的質(zhì)量最多,含量(即純度)也最高(達(dá)到43.89%);菌株LKH以油酸為唯一碳源生長(zhǎng)時(shí)的PHB提取率最高(49.70%);菌株LKH以魔芋精粉為唯一碳源合成PHB的質(zhì)量可達(dá)0.52 g/L,含量達(dá)到40.62%,PHB提取率可以達(dá)到40.25%。
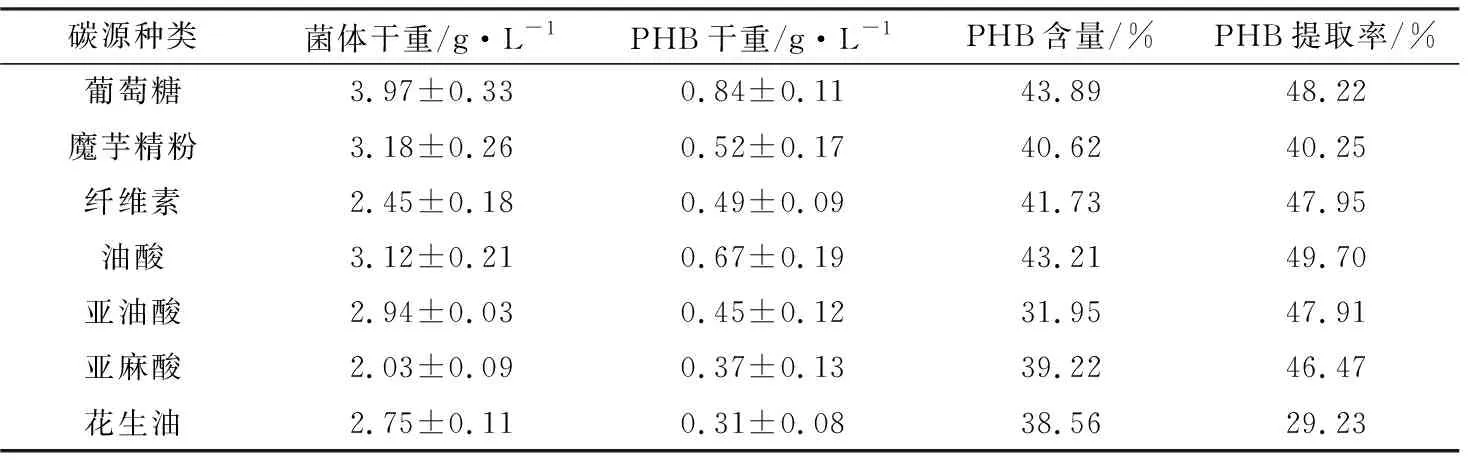
表2 不同碳源培養(yǎng)的菌株LKH中提取的PHB含量及提取率

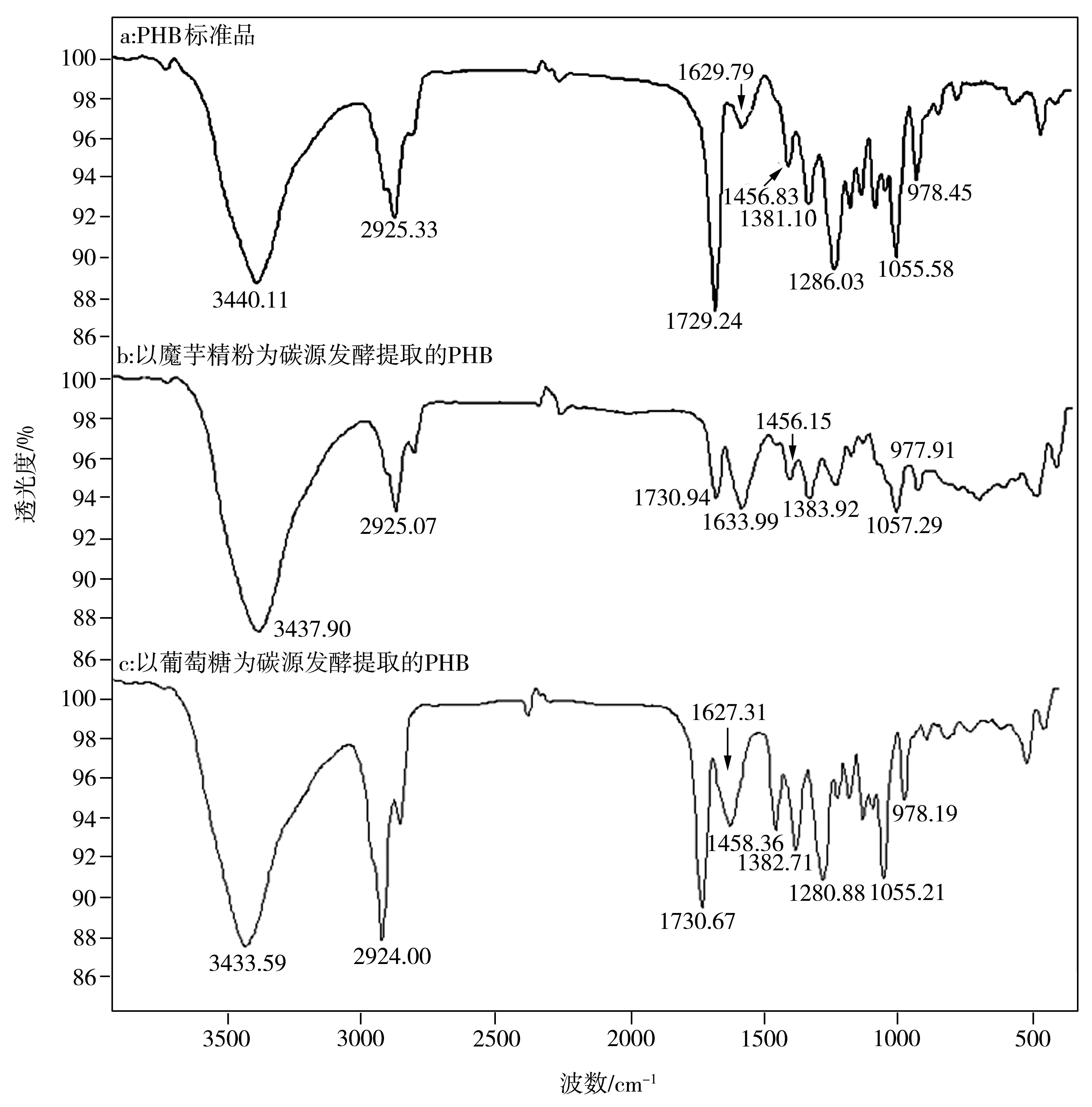
圖7 以魔芋精粉和葡萄糖為唯一碳源培養(yǎng)的菌株LKH中所提取PHB的紅外光譜
不同碳源培養(yǎng)的菌株LKH中提取出來(lái)的白色薄膜樣品經(jīng)甲酯化后進(jìn)行氣相色譜分析,其結(jié)果顯示均與PHB標(biāo)準(zhǔn)品一致。圖8為PHB標(biāo)準(zhǔn)品、分別以魔芋精粉和葡萄糖為唯一碳源培養(yǎng)的菌體中合成的白色薄膜樣品的氣相色譜。由圖可見(jiàn),提取的樣品在1.49 min附近出現(xiàn)氯仿峰,在1.67min附近出現(xiàn)甲酯化的PHB單體組分峰,在5.42min附近出現(xiàn)內(nèi)標(biāo)物苯甲酸的峰,表明提取物中含有一定量的PHB。但是,氣相色譜圖中還有很多雜峰,表明提取的PHB純度不高,這與前面所測(cè)的PHB含量約為40%的結(jié)果符合。
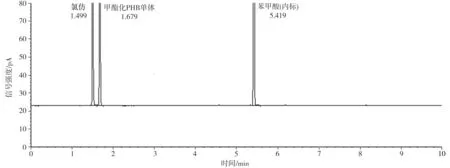
(a)PHB標(biāo)準(zhǔn)品
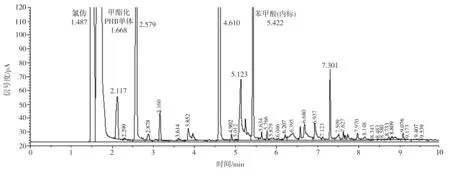
(b)以魔芋精粉為碳源發(fā)酵提取的PHB
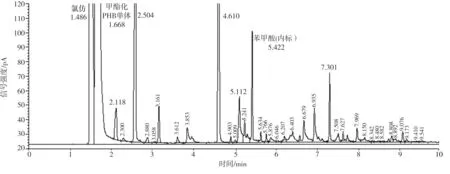
(c)以葡萄糖為碳源發(fā)酵提取的PHB
2.4.2 氮源濃度對(duì)PHB合成的影響
聚羥基烷酸酯是一類(lèi)微生物在不平衡的生長(zhǎng)條件下胞內(nèi)合成的碳源和能源貯藏物,所謂不平衡生長(zhǎng)條件為碳源相對(duì)過(guò)量,氮源等營(yíng)養(yǎng)物質(zhì)相對(duì)缺乏。因此,本試驗(yàn)在固定碳源為魔芋精粉的基礎(chǔ)上,考察了不同氮源(NH4Cl)濃度對(duì)PHB合成的影響,結(jié)果如表3所示。從表3中可知,氮源濃度越低,即C/N比越大,PHB的提取率就越高;在氮源濃度為0.2 g/L時(shí),PHB的質(zhì)量達(dá)到最高(0.85 g/L),含量高達(dá)69.55%,提取率達(dá)到93.29%。
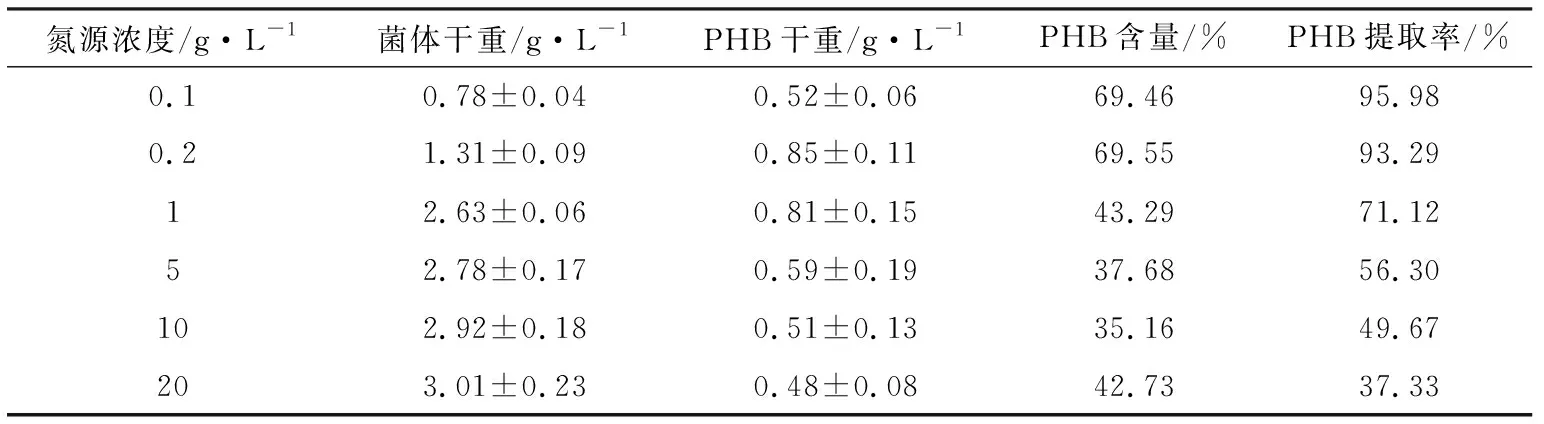
表3 不同氮源濃度下培養(yǎng)的菌株LKH中提取的PHB含量及提取率
3 結(jié)論
(1)從云南種植魔芋的土壤中分離得到了一株能以魔芋多糖為唯一碳源生長(zhǎng)的菌株LKH,該菌能夠分泌β-甘露聚糖酶水解魔芋多糖生長(zhǎng),經(jīng)蘇丹黑染色鑒定,菌株可以在胞內(nèi)積累PHB。
(2)經(jīng)形態(tài)學(xué)、生理生化鑒定和16s rDNA序列同源性分析,菌株LKH歸屬為依利諾斯類(lèi)芽孢桿菌Paenibacillusillinoisensis。

(4)在氮源濃度為0.2 g/L的條件下,菌體LKH以魔芋多糖為唯一碳源合成的PHB質(zhì)量高達(dá)0.85 g/L,含量高達(dá)69.55%,PHB提取率為93.29%。P.illinoisensisLKH在發(fā)酵魔芋多糖生產(chǎn)魔芋低聚糖的同時(shí)可胞內(nèi)合成PHB,實(shí)現(xiàn)了菌體廢棄物的二次開(kāi)發(fā)利用,有望降低微生物發(fā)酵生產(chǎn)PHB的成本。
[1] 賈成禹.魔芋及魔芋多糖[J].植物雜志,1994(5):26-27.
[2] 成莉鳳,戴小陽(yáng),馮湘沅,等.BacillussubtilisBE-91生長(zhǎng)及其胞外表達(dá)β-甘露聚糖酶的發(fā)酵條件優(yōu)化[J]. 微生物學(xué)通報(bào),2015,42(12):2300-2307.
[3] 李劍芳,鄔敏辰,程科,等.β-甘露聚糖酶制備魔芋葡甘露低聚糖的研究[J].食品與發(fā)酵工業(yè),2007,33(1):21-24.
[4] 趙梅,王春娟,董云海,等. 重組β-甘露聚糖酶制備低聚葡甘露糖工藝條件的優(yōu)化[J].食品與生物技術(shù)學(xué)報(bào),2016,35(7):704-708.
[6] 周迎鑫,楊楠,王希媛,等.聚羥基烷酸酯(PHA)改性研究進(jìn)展[J].生物工程學(xué)報(bào),2016,32(6):738-747.
[7] Nielsen C, Rahman A, Rehman A U, et al. Food waste conversion to microbial polyhydroxyalkanoates[J]. Microbial Biotechnology, 2017,10(6):1338-1352.
[9] Jiang G Z, Hill D J, Kowalczuk M, et al. Carbon sources for polyhydroxyalkanoates and an intergrated biorefinery [J]. International Journal of Molecular Sciences, 2016, 17(7):1157.
[12] 沈萍,范秀容,李廣武.微生物學(xué)實(shí)驗(yàn)[M].北京:高等教育出版社,1999.
[13] 李長(zhǎng)影,孔雯,王家昕,等.β-甘露聚糖酶產(chǎn)生菌的分離鑒定和酶學(xué)性質(zhì)[J].華中農(nóng)業(yè)大學(xué)學(xué)報(bào),2011,30(2):138-142.
[14] 李凌凌,呂早生,沈惠莉,等.產(chǎn)聚羥基烷酸的菌株分離及初步鑒定[J].武漢科技大學(xué)學(xué)報(bào):自然科學(xué)版,2007,30(5):502-505.
[15] Getachew A, Woldesenbet F. Production of biodegradable plastic by polyhydroxybutyrate (PHB) accumulating bacteria using low cost agricultural waste material[J]. BMC Research Notes, 2016, 9(1): 509-517.
[16] 呂早生,張敏,李凌凌,等.蠟狀芽孢桿菌產(chǎn)聚羥基烷酸的提取與鑒定[J].化學(xué)與生物工程,2008,25(8):52-54,66.
[17] Krishnan S, Chinnadurai G S, Perumal P. Polyhydroxybutyrate byStreptomycessp.: production and characterization[J]. International Journal of Biological Macromolecules, 2017,104(A):1165-1171.
[18] Shida O, Takagi H, Kadowaki K, et al. Emended description ofPaenibacillusamylolyticusand description ofPaenibacillusillinoisensissp. nov. andPaenibacilluschibensissp. nov.[J]. International Journal of Systematic Bacteriology, 1997, 47(2): 299-306.
[19] Jhala Y K, Vyas R V, Shelat H N, et al. Isolation and characterization of methane utilizing bacteria from wetland paddy ecosystem[J]. World Journal of Microbiology and Biotechnology, 2014, 30(6): 1845-1860.
[20] Abid S, Raza Z A, Hussain T. Production kinetics of polyhydroxyalkanoates by usingPseudomonasaeruginosagamma ray mutant strain EBN-8 cultured on soybean oil[J]. 3 Biotech, 2016, 6(2): 142.
[21] 田頻源,米鈺,尚龍安,等.聚羥基烷酸酯的研究進(jìn)展[J].化工進(jìn)展,2009,28(3):468-476.
[22] 田宇曦,閔勇,楊自文,等.多粘類(lèi)芽孢桿菌研究進(jìn)展[J].湖北農(nóng)業(yè)科學(xué),2017,56(18):3401-3404,3409.
[23] Bhuwal A K, Singh G, Aggarwal N K, et al. Isolation and screening of polyhydroxyalkanoates producing bacteria from pulp, paper, and cardboard industry wastes[J]. International Journal of Biomaterials, 2013, 2013: 752821.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
