山西沁河上游植物種間關系及功能群劃分
2018-06-19 11:49:50田益瑆郭東罡
山西農業科學 2018年6期
田益瑆,郭東罡
(山西大學環境與資源學院,山西 太原 030006)
沁河屬于黃河一級支流,是黃河三門峽至花園口區間兩大支流之一,全長為456 km,流域面積為12 900 km2,包括山西省和河南省16個縣(區),流域內煤炭資源豐富,人類活動歷時悠久。沁河發源于山西省太岳山山區,上游流經縣域均為我國的產煤大縣,其生態系統服務功能尤為重要,但是關于沁河上游濕地植被群落生態學的系統研究并不多見。
本研究在樣方調查的基礎上,采用χ2檢驗、聯結系數(AC)和共同出現百分率(PC),研究了沁河上游濕地植物群落的種間關聯性,并對種間相關的測定結果進行了生態分析[1-3],旨在為保護沁河源頭水環境質量、濕地生物多樣性和科學評價當地生態本底等提供科學數據。
1 研究區域概況
沁河上游主要位于山西省長治市境內,長度為86.40km,處于東經 112°3′12.5552″~112°24′46.4175″,北緯 36°22′42.7105″~36°52′28.2353″。屬典型的暖溫帶大陸性季風氣候,年均氣溫5.5~7.8℃,1月份平均氣溫 -6.8~-10.4℃,7月份平均氣溫17.4~20.9℃;通常的無霜期是90~150 d,年均降水量656.7 mm;年平均日照時數2 519 h。土壤主要為沖積土、沼澤土和草甸土等。
2 研究方法
2.1 樣地設置與調查
在踏查的基礎上,采用系統隨機取樣法,在沁河上游沿岸,從源頭和主要水源補給區順流而下,盡量避開人為干擾,在河岸兩側設置了7塊樣地,在每塊樣地內,平行于河岸設置6條1 m×10 m樣線,河岸兩側各設置3條,每條樣線間隔5 m,在每條樣線上取10個1 m×1 m的樣方,共計樣方數420個。在每個樣方內記錄植物種名、蓋度、高度、多度、物候期、生活型和生活力等,同時記錄相關環境因子情況[4]。各樣地基本概況列于表1。
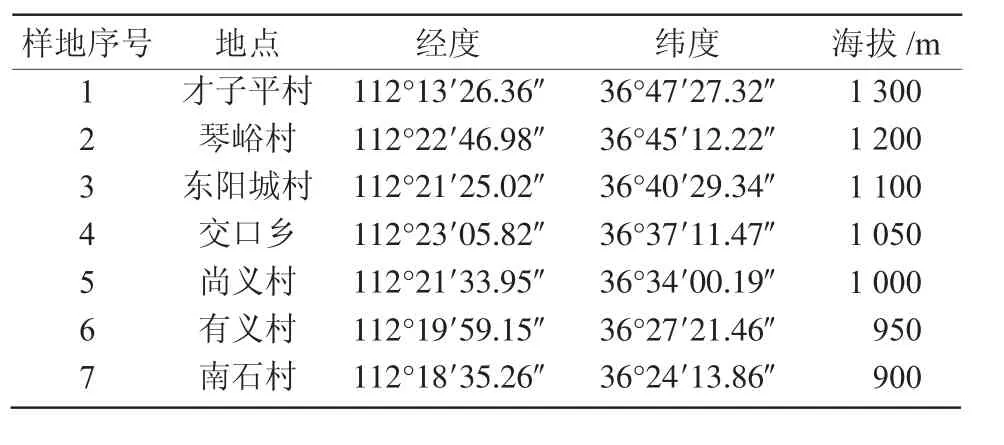
表1 樣地基本概況
2.2 數據處理
種間聯結性測定,χ2檢驗是運用二元定性數據測定種間聯結,將40個高頻種和420個樣方組成40×420的種-樣方矩陣,以這些數據為基礎,得出這40個高頻種780個種對應的2×2列聯表,再以此表為基礎對所列出的種對之間的種間關聯系數進行計算,即對a,b,c與d值進行計算。然后利用χ2檢驗,與AC和PC相結合對物種間聯結性進行測定。
2.2.1 常用的用于確定物種間是否存在關聯關系的χ2檢驗方法 依據所列出的聯表中的χ2統計量,通過如下公式進行檢驗。
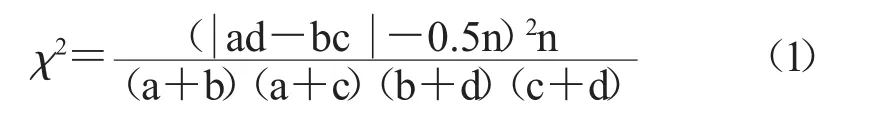
其中,n指的是樣方總數,a指的是2個物種均出現的樣方數量,b指的是A物種出現而B物種不出現的樣方數量,c指的是B物種出現而A物種不出現的樣方數量,d指的是A物種和B物種都不出現的樣方數量。當ad>bc時為正聯結,當ad<bc時為負聯結。當χ2>χ20.05(1)(此時χ2對應的自由度是1,顯著性P對應的值為0.05)說明種間的關聯性顯著;χ2>χ20.01(1)(此時 χ2的值對應的自由度為1,顯著性P值為0.01)則說明是極顯著的,反之則是不顯著。χ20.05(1)=3.841,χ20.01(1)=6.635,即當3.841<χ2<6.635 為關聯顯著,χ2>6.635 為極顯著,χ2<3.841為關聯不顯著。
2.2.2 聯結系數AC AC系數可以用來對χ2檢驗結果進行檢驗,并表示種間聯結程度。
若 bc>ad,且 d≥a,則有下式。

若 bc>ad,且 d<a,則有下式。

若ad≥bc,則有下式。
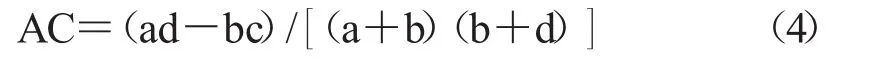
AC的值域為[-1,1]。當AC所對應的值越趨近于1的情況下,說明對應的物種間的正聯結性越強;反之,AC值更加趨近于-1,則說明物種間的負聯結性越強;當AC值為0時,物種間完全獨立。
2.2.3 共同出現百分率PC PC值可以表示物種間的正聯結程度。

式中,PC的值域為[0,1]。當PC值越靠近1時,說明物種之間的正聯結程度越強[5-7]。
3 結果與分析
3.1 群落組成
本研究選取了研究區域內頻度>5%的物種為高頻草本物種,結果顯示,共有40個高頻種,屬于15科35屬,其中,水生植物與濕生植物有8種,中生植物與旱生植物有32種。主要高頻種為狗尾草(Setaria viridis)、節節草(Commelina diffusa)、棒頭草(Polypogon fugax)、青蒿(Artemisia carvifolia)、堿茅(Puccinellia distans)、早開堇菜(Viola prionantha)、蘆葦(Phragmites australis)、花苜蓿(Medicago ruthenica)、車前(Plantago asiatica)、旋覆花(Inula japonica)等。
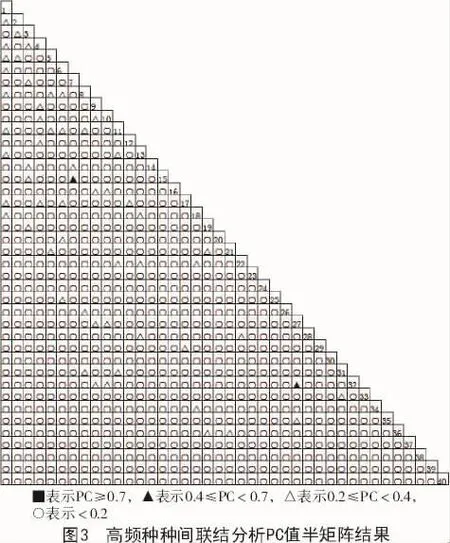
3.2 種間聯結測定的χ2值、AC值和PC值特征分析
根據表2以及圖1,2,3可知,由于所測指標性質不同,χ2,AC值和PC值3個指數有所差異。χ2能夠體現種間聯結的顯著狀況,但它只能體現種間聯結程度是否顯著,對于種間聯結程度不顯著的種對,不能準確體現種對間的聯結程度。AC和PC更能夠客觀地對通過χ2檢驗結果為不顯著聯結種對間所對應的聯結程度進行檢驗[7-9]。聯結系數AC和共同出現百分率PC雖然能反映種對間聯結程度強弱,但缺乏統一的標準劃分種間聯結程度。假若物種出現頻率低,特別是在a值是0的時候,AC和PC對應的值對物種間關聯程度有放大作用,甚至有時候會得到相反的結果。如果葎草、柳葉菜、地榆、荊三棱等物種的出現頻率較低,次數較少,這些物種和其他高頻種一起出現的樣方數的a值等于0的概率較大。極顯著負關聯(AC<0.60)高達265對,正是這種原因所造成的。當物種出現次數較多時,共同出現百分率PC值能夠避免由于2個物種均未出現的樣方數d偏高而造成聯結系數AC值偏高的問題,但PC所對應的值只能對種對之間的正聯結程度做出較為準確的反應。發生此種情況的原因是由于在a值比較小的情況下,會對d值進行忽略,對a,b,c的作用值進行夸大,同時夸大了負聯結程度。這種現象在出現次數較少物種出現時更為明顯。即物種的分布較為均勻時,AC和PC的數值是準確的,而在沁河上游河岸帶生態系統里,由于受到人為或自然因素的影響,河岸帶生態系統受到破壞,物種的分布均勻度較差[10-12]。
總之,選擇40個草本高頻種。經過χ2檢驗的同時,把AC和PC所對應的值作為用于檢驗的輔助參數,對沁河河岸帶生態系統里面存在的主要高頻種種間存在的關聯性進行分析[13-14]。

表2 χ2檢驗值、AC值、PC值
3.3 種間聯結性
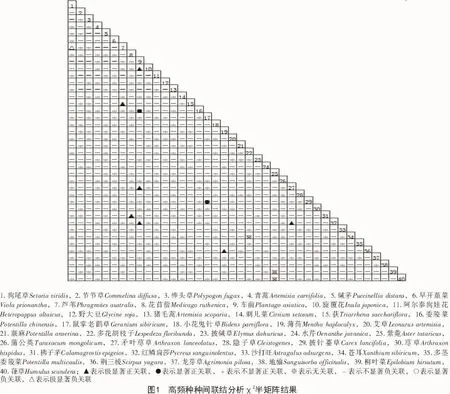
χ2檢驗結果說明,在全部的40個草本高頻種所進行組成的780個種對中,表現為正聯結一共有319對,占總量的40.90%,表現為負聯結的有458對,占總量的58.72%,表現為無關聯的種對為3對,占總種對數的0.38%。正負聯結種對數之比為0.70∶1,由此可知,沁河上游河岸帶草本群落種對間正負聯結差異較大,河岸帶群落不夠穩定,易于受到外部因素干擾。種對間呈顯著聯結以及呈極顯著聯結的種對數較少,種對間顯著正聯結以及極顯著正聯結的種對有9對,占總種對數的1.15%,種對間顯著負聯結以及極顯著負聯結的種對有1對,占總種對數的0.13%,表明沁河上游河岸帶草本群落種對間聯結性較差,種間的獨立性強,生態位分離明顯,在生長過程中,物種之間相互影響較小,容易受到人為或自然因素影響,河岸帶草本群落穩定性差。呈極顯著正聯結的種對為:蘆葦和荻,花苜蓿和拂子茅,車前和旋覆花,薄荷和荊三棱,矛葉藎草和紅鱗扁莎等;呈顯著正聯結的種對為:車前和委陵菜,鼠掌老鸛草和披針薹草;呈極顯著負聯結的種對為狗尾草和蘆葦;無關聯種對為:多花胡枝子和蒼耳,紫菀和柳葉菜,紫菀和葎草。
AC值結果表明,沁河上游河岸帶草本群落中40個高頻種種對間呈極顯著正聯結性(AC≥0.6)的種對有0對,顯著正聯結(0.2≤AC<0.6)種對有35對,占總對數的4.49%,不顯著正聯結(0.05≤AC<0.2)種對有147對,占總對數的18.85%;無聯結(-0.05≤AC<0.05)的種對有159對,占總種對數的20.38%;不顯著負聯結(-0.2≤AC<-0.05)的種對有49對,占總對數的6.28%,顯著負聯結(-0.6≤AC<-0.2)的種對有125對,占總對數的16.3%,其中,表現為極顯著負聯結(AC<-0.6)的一共有265對,占33.97%。由此可知,該區域草本植物物種間聯結性較弱,各物種偏向獨立分布。
PC值結果表明,沁河上游河岸帶草本群落中40個高頻種種對間極顯著正聯結性(PC≥0.7)的種對有0對;顯著正聯結(0.4≤PC<0.7)種對有2對,占總對數的0.26%;不顯著正聯結(0.2≤PC<0.4)的種對有55對,占總對數的7.05%;無聯結(PC<0.2)的種對有723對,占總種對數的92.69%。PC值可以對物種間的正聯結存在的強弱關系進行較為準確地表示。
綜上所述,種間聯結測定的χ2值、AC值和PC值3種方法的結果均表明,沁河上游河岸帶草本群落物種間正聯結性弱,物種之間的相互影響程度不強,群落不夠穩定,易受到人為或自然因素的干擾[15]。
3.4 功能群劃分及分析
在群落內各個物種生態習性各不相同,種群之間的關聯揭示了由于生境的影響不同群落的物種之間空間分布的關系[16-18]。本研究基于種間正聯結性為基礎、負聯結性為界限以及草本高頻種對水分因子的響應,對草本植物功能群進行了劃分[19-20]。各功能群間的物種間存在正相關關系,這是由物種間的同一資源利用模式以及功能群內同類生態位所決定的[21]。功能群的劃分不僅需要基于種間關系,還需要基于水分因子和其他環境因子[22-23]。

山西沁河上游草本植物群落的40個高頻種可分為4個功能群:Ⅰ.水生植物功能群:包括蘆葦、水芹以及荊三棱。該功能群物種對水分條件要求高,常見于地下水位較深、水分充足的環境中,廣泛分布在沁河濕地的河道以及常年積水的沼澤等地區。Ⅱ.濕生植物功能群:包括旋覆花、薄荷、拂子茅、紅鱗扁莎、柳葉菜。種間關系大多數為顯著正關聯以及正關聯,生態習性相近。該功能群物種對水分條件要求較高,常見于地下水位較淺的地方,抗旱屬性差,不能長時間在缺水環境下生長,廣泛分布在沁河季節性積水的沼澤地和河漫灘等地區。Ⅲ.中生植物功能群:包括節節草、棒頭草、青蒿、堿茅、野大豆、刺菜、蕨麻、紫菀、蒲公英、龍牙草以及地榆。該功能群物種主要分布在沁河河岸邊以及沼澤地邊等河灘一級階地等潮濕地區。Ⅳ.中旱生植物功能群:包括狗尾草、早開堇菜、花苜蓿、車前、阿爾泰狗娃花、豬毛蒿、荻、委陵菜、鼠掌老鸛草、小花鬼針草、艾草、多花胡枝子、披堿草、矛葉藎草、隱子草、披針薹草、藎草、沙打旺、蒼耳、多莖委陵菜以及葎草。該功能群物種主要分布在沁河河灘二級階地以及濕地附近的路旁、林下、田邊和河漫灘地勢較高處地區。
通過功能群的劃分可以分析得出,不同的功能群對生境具有不同的趨同性適應。而水分因子成為沁河上游濕地草本植物劃分功能群的重要環境因子,起著決定性的作用。功能群的不同也反映了草本植物群落隨著水分梯度的變化而產生明顯的演替規律。
4 討論與結論
利用高頻種種間關系和水分條件劃分植物功能群的方法,是對濕地生態系統功能群進行分類的一種嘗試。高頻種在生態系統中控制群落結構,能較好地反映一個區域的生態系統狀況,因此,以高頻種的選擇為研究對象對植物功能群進行劃分。沁河上游濕地在不同樣地草本植物高頻種的變化,揭示了高頻種對水分條件變化的不同反應。例如,中旱生植物功能群廣泛分布在濕地附近的路旁、林下、田邊和河漫灘地勢較高處地區,而水生植物功能群廣泛分布在沁河濕地的河道以及常年積水的沼澤等地區。
對植物功能群的研究已經成為研究群落動態穩定性、植被分布格局、生態系統總體生產力的重要手段[12],所以,對于功能群劃分的研究有著重要意義。濕地植物功能群的劃分,將植物功能群中的物種作為一個整體研究其對水分因子以及外界干擾響應的規律,要比單個物種的研究更容易歸納出規律性變化[20]。功能群的劃分要以種間關系為基礎,結合其他因子,例如海拔[23]、物種生態習性等因子,具體劃分方法要根據研究地區實際情況決定。
以高頻種種間關系為基礎,結合水分條件對沁河上游草本植物群落劃分功能群符合生態學原理,符合當地實際生態現狀。如果只是以物種間相關性為依據劃分功能群,蘆葦與荻為顯著正相關,但2種物種并不同屬一個功能群,蘆葦常年生長在沁河河道以及常年積水的沼澤、灘涂,為水生植物功能群,而荻廣泛分布于沁河濕地兩旁的田邊、林下等地勢較高處,為中旱生植物功能群,二者生態習性大相徑庭,并不能劃分為同一個功能群。沁河上游濕地的草本植物在長期自然演替規律下,具有相近生態習性的物種會共同組成一個功能群,在同屬一個功能群內物種間具有正聯結性,經常結伴出現,對水分條件具有共同的反映趨勢,生態位相近,在群落中相互利用、相互促進生長;而表現為負相關性的物種,對生境的反映趨勢不同,很少共同出現,成為劃分功能群的界限。在本研究中,沁河濕地水分生態功能群隨著水分梯度的增加,土壤水分增加,水生植物與濕生植物的比例隨之增加,在這種生境下,中旱生植物生長得到抑制,而水生植物與濕生植物的競爭力得到加強,因此,在群落中獲得絕對優勢;相反,水分條件減少,土壤水分降低,水分不充足時,耐旱的中旱生植物為優勢物種。
沁河上游濕地草本植物物種間呈極顯著以及顯著相關的種對較少,整體物種間相關性較弱,這是由于所選取樣地生境差異較大,有沁河濕地的河道以及常年積水的沼澤、季節性積水的沼澤地和河漫灘、沁河河岸邊、濕地附近的路旁、林下、田邊和河漫灘地勢較高處等環境,而且物種生態學特性差異較大[13]。同時,40個高頻種種對間正相關種對數小于負相關種對數。這是由于沁河上游附近農業的發展,人口的增長,沁河上游濕地附近地區成為人類活動區域,人為干擾因素增加,對濕地植物群落造成影響,使得植物群落不夠穩定,景觀破碎化加劇。沁河上游濕地生態系統雖然物種組成比較豐富,但也非常脆弱,人為因素以及自然因素的影響使得其生物、環境均受到一定程度的威脅。因此,應以沁河上游濕地草本植物功能群劃分研究為基礎,加強沁河上游的污染防治,并采取人工措施恢復該地區環境,使物種具有多樣性,促進沁河上游濕地的健康發展[24]。
[1]KLEYER M.Validation of plant functional types across two contrasting landscapes[J].Journal of Vegetation Science,2002,13(2):167-178.
[2] SMITH T M,WOODWARD F I,SHUGART H H.Plant function types[M].Cambridge:Cambridge University Press,1997.
[3]李榮平,劉志民,蔣德明,等.植物功能型及其研究方法[J].生態學雜志,2004,23(1):102-106.
[4]賈慧君,李帥,郝婧,等.黃河中游(龍門-汾河入黃口)水分因子與濕地植物多樣性的相關關系研究 [J].山西農業科學,2017,45(8):1325-1330.
[5]WOODWARDF I,CRAMER W.Plant functional types and climatic changes:introduction[J].Journal of Vegetation Science,1996,7(3):306-308.
[6]胡楠,范玉龍,丁圣彥,等.陸地生態系統植物功能群研究進展[J].生態學報,2008,28(7):3302-3311.
[7]郭屹立,盧訓令,丁圣彥.伊洛河河岸帶生態系統草本植物功能群劃分[J].生態學報,2012,32(14):146-154.
[8] GRAFF P,AGUIAR M R,CHANETON E.Shifts in positive and negative plant interactions along a grazing intensity gradient[J].E-cology,2007,88:188-199.
[9]嚴慧玲,劉東明,王俊,等.河北邢汾高速公路沿線植物群落優勢種的種間關系[J].河南大學學報(自然科學版),2015,45(4):76-83.
[10]王燁,秦浩,張峰,等.濁漳河干流濕地草本植物群落優勢種種間關系及功能群劃分 [J].生態學雜 志,2015,34(8):2109-2114.
[11]房飛,胡玉昆,張偉,等.高寒草原植物群落種間關系的數量分析[J].生態學報,2012,32(6):1898-1907.
[12]段后浪,趙安,姚忠.鄱陽湖區典型草洲植物種間關聯性研究[J].濕地科學,2017,15(1):107-113.
[13]BROOKER R W,MAESTRE F T,CALLAWAY R M.Facilitation inplant communities:The past,the present,and the future[J].Journal of Ecology,2008,96:18-34.
[14]王建良,趙成章,張偉濤,等.秦王川濕地鹽角草和鹽地堿蓬種群的空間格局及其關聯性 [J].生態學雜志,2017,36(9):2494-2500.
[15]劉亞琦,劉加珍,陳永金,等.孔雀河下游斷流河道的環境特征及物種間關系[J].生態學報,2017,37(8):2706-2718.
[16]范曉,張峰,龐春花,等.山西汾河流域下游草本植物功能群[J].生態學雜志,2013,32(10):2716-2720.
[17]李潮,謝應忠,許冬梅,李俠.寧夏荒漠草原植物群落的種間關系[J].草業科學,2013,30(11):1801-1807.
[18]李蘭蘭,許諾,李洪遠,等.七里海濕地植物種間關系的數量分析[J].水土保持通報,2014,34(4):70-75.
[19]程雷星,陳克龍,蘇旭,等.青海湖流域小泊湖濕地植物種間關系研究[J].干旱區地理,2014,37(5):1005-1011.
[20]吳華,張建利,喻理飛,等.草海流域水源功能區植物持水功能群劃分[J].水土保持研究,2014,21(2):138-143.
[21]劉玨宏,高慧,張麗紅,等.內蒙古錫林郭勒草原大針茅-克氏針茅群落的種間相關特征分析 [J].植物生態學報,2010,34(9):1016-1024.
[22]秦曉娟,高璐,鄧永利,等.山西平陸黃河濕地植功能群劃分[J].山西大學學報(自然科學版),2014,37(3):454-460.
[23]趙艷云,劉京濤,陸兆華.渤海灣貝殼堤濕地蘆葦種群與蒙古蒿種群空間分布格局和種間關系 [J].濕地科學,2017,15(2):187-193.
[24]王思元,牛萌.濕地系統的生態功能與濕地的生態恢復[J].山西農業科學,2009,37(7):55-57.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中華詩詞(2022年6期)2022-12-31 06:41:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
