低溫脅迫對高山杜鵑生理特性影響的研究
2018-05-16 13:03:53李小玲華智銳賀廣祥
江西農業學報 2018年5期
關鍵詞:植物
李小玲,華智銳,祝 穎,賀廣祥
(商洛學院 生物醫藥與食品工程學院,陜西 商洛 726000)
高山杜鵑(Rhododendrondelavayi)是杜鵑花科杜鵑花屬植物的總稱。它既是世界三大高山花卉之一,也是中國十大名花之一[1]。高山杜鵑原種的70%早在100多年前從中國引入歐洲,歐洲人經過上百年的研究,培育出了中國杜鵑品種與歐美杜鵑品種的雜交種[2]。高山杜鵑耐寒,喜陰涼和濕潤的環境,多生長在海拔1000 m以上的高山、草甸、林緣地區[3]。陜西秦嶺野生杜鵑花屬植物以其豐富的種類、鮮艷多姿的花型、四季常青的特點博得了人們的喜愛,它不僅具有一定的觀賞價值,而且其枝葉可以用來入藥,因此具有較高的開發利用價值[4]。鎮安木王國家森林公園,地處陜西省商洛市鎮安縣楊泗鄉,享有“千山杜鵑”、“杜鵑故里”的美譽。該地區杜鵑花種類主要包括秀雅杜鵑、美容杜鵑、頭花杜鵑、照山白等,其中分布數量最大,面積最廣的為美容杜鵑[5]。
溫度是植物正常生理代謝的主要生態因子之一,極端溫度不僅會影響植物的正常生長發育、引起植物代謝紊亂,而且嚴重時還會造成植株的死亡。低溫是植物生長發育過程中經常遭受的非生物脅迫因子之一[6],低溫可作為環境信號啟動細胞內信號轉導途徑,引起植物體內酶活性的改變及基因表達,進而影響植物重要的生理生化反應[7-8]。
本研究以秦嶺高山杜鵑幼苗為材料,通過探討低溫脅迫對高山杜鵑丙二醛(MDA)含量、相對電導率、游離脯氨酸含量、可溶性蛋白含量、超氧化物歧化酶(SOD)活性、過氧化物酶(POD)活性、過氧化氫酶(CAT)活性的影響,從理論上揭示高山杜鵑對低溫脅迫的生理生態適應機制,為高山杜鵑休眠狀態的解除和抗寒性品種的選育提供理論依據和實驗指導。
1 材料與方法
1.1 試驗材料
試驗所用高山杜鵑幼苗于2017年4月上旬采自商洛市鎮安縣木王國家森林公園海拔1500 m左右的茨溝杜鵑花景區。
1.2 試驗方法
1.2.1 材料預培養 將采集的株高10 cm的高山杜鵑幼苗移栽到直徑為15 cm的花盆中,每盆種植1株,盆栽基質為腐葉土∶松針土=1∶2。培養期間于每天傍晚噴灑0.1%~0.5%的硫酸亞鐵水溶液以保持盆土濕潤。低溫脅迫前將所栽的高山杜鵑幼苗先放入常溫下連續培養10 d,然后選取生長一致的幼苗用于試驗。
1.2.2 試驗設計 本試驗分別設置0、5、10、25 ℃這4個溫度梯度處理高山杜鵑幼苗,其中以25 ℃作為對照溫度。然后將試驗材料分別放入設有不同溫度的人工氣候箱內,相對濕度為72%,每天光照處理12 h,光照強度約為1000 lx,黑暗處理12 h,連續培養6 d。經過低溫脅迫后,分別采第1天、第2天、第3天、第4天、第5天的高山杜鵑葉片并進行處理,每個處理溫度稱取1.5~3.0 g除去主脈、健康生長無病害的葉片,分別測定高山杜鵑相關的生理指標,每次試驗重復3次。
1.2.3 各項生理指標的測定 相對電導率測定參照陳建勛等[9]的電導儀法;游離脯氨酸含量測定參照許維岸[10]的茚三酮法;MDA含量的測定參照張志安等[11]的硫代巴比妥酸法;SOD活性的測定參照李合生[12]的氮藍四唑(NBT)光還原法;POD活性的測定參照郝再彬等[13]的愈創木酚顯色法;CAT活性的測定參照高俊鳳[14]的紫外光譜法;可溶性蛋白含量的測定參照高俊鳳[14]的考馬斯亮藍G-250法。
1.3 數據處理
所有處理每次測定重復3次,取3次測定值的平均值,用Excel 2010進行數據統計,用SPSS Statistics 17.0軟件進行數據分析。
2 結果與分析
2.1 低溫脅迫對高山杜鵑相對電導率的影響
由圖1可知,與對照溫度相比,其它溫度處理下相對電導率均明顯高于對照組,且溫度越低增加幅度越大。而不同溫度處理下,除對照組溫度條件下電導率相對平穩外,隨著時間延長相對電導率均呈現上升趨勢,到處理第5天時,0、5、10 ℃下電導率與對照組25 ℃時相比,增加幅度分別達到90.11%、49.79%和27.07%,差異達顯著水平(P<0.05)。
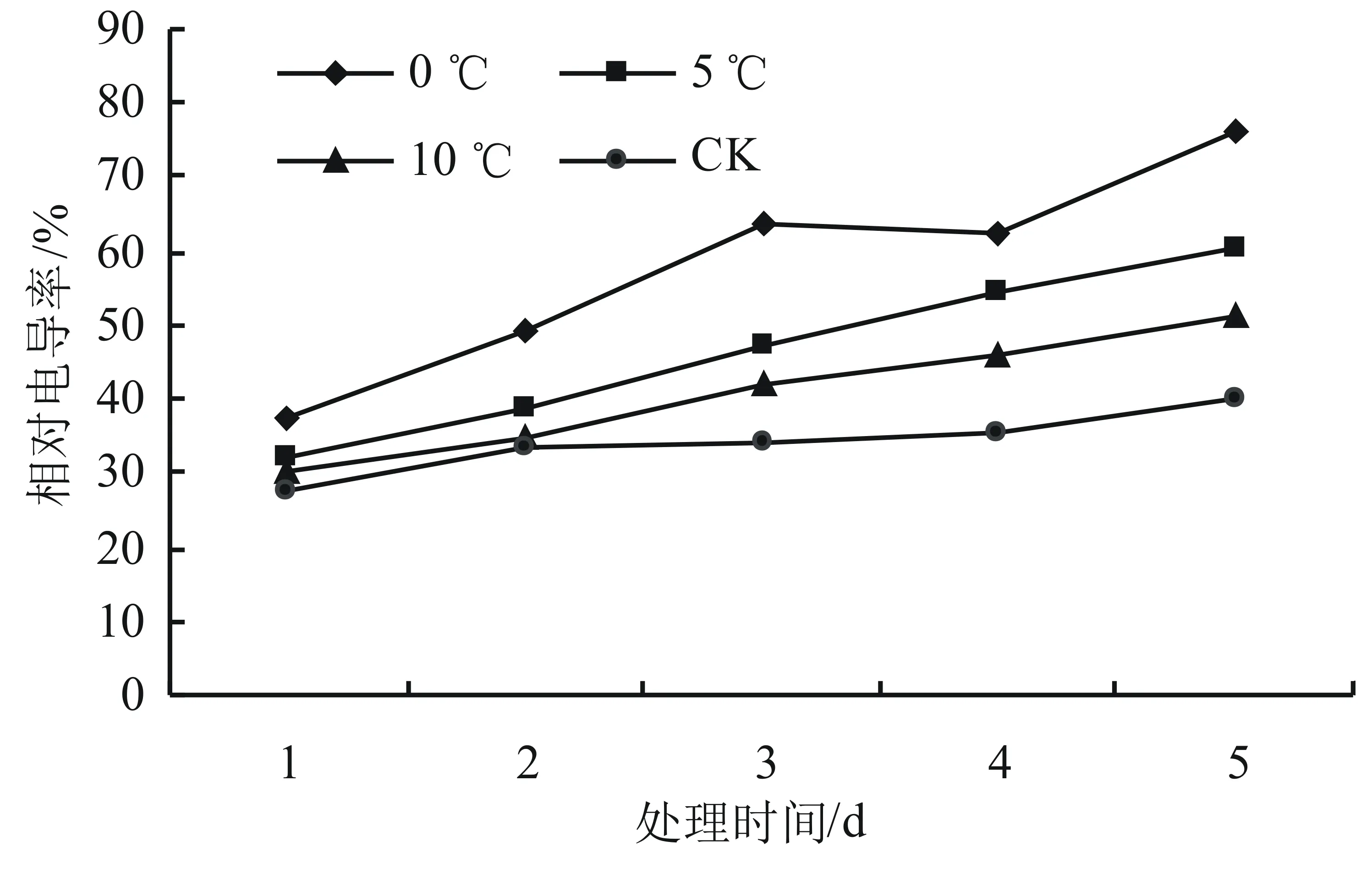
圖1 低溫脅迫對高山杜鵑相對電導率的影響
2.2 低溫脅迫對高山杜鵑MDA含量的影響
由圖2可知,高山杜鵑幼苗的MDA含量在相同溫度下隨著時間的增加而增加。而在同一時間的不同溫度下,溫度越低,MDA含量越高。0 ℃時MDA含量的變化最大,上升最快。與對照溫度處理下的MDA含量相比,10 ℃處理下的MDA含量變化不顯著(P>0.05);5 ℃處理下的MDA含量則顯著增加(P<0.05),比對照增長了68.32%;而0 ℃處理下的MDA含量差異最為顯著(P<0.05),與對照相比增加幅度為88.12%。
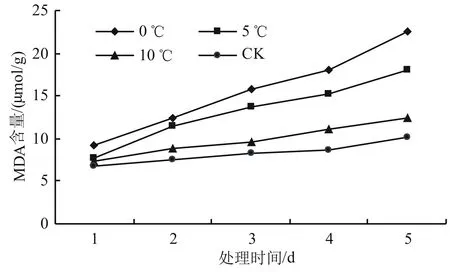
圖2 低溫脅迫對高山杜鵑MDA含量的影響
2.3 低溫脅迫對高山杜鵑SOD、POD和CAT活性的影響
由圖3可以看出,隨著低溫脅迫時間的延長,SOD的活性呈先升后降再上升的趨勢,總體上呈現上升的趨勢。在相同時間內,隨著溫度的降低,高山杜鵑幼苗葉片的SOD活性上升,第5天時0、5、10 ℃處理下杜鵑幼苗葉片的SOD活性顯著高于室溫處理下的SOD活性(P<0.05),與對照相比增加幅度分別為84.87%、54.38%和35.35%。由圖4可以看出,POD活性總體呈上升趨勢。對照溫度下POD活性在第1天最高,0 ℃最低。而在第2天時,0 ℃時的POD活性增長最快,比第1天增長了43.21%,達到同一天不同溫度處理下POD活性的最高值。總體上,在試驗期間0 ℃時POD活性增長最快,比第一天增長了86.73%,差異顯著(P<0.05);5 ℃和10 ℃下POD活性的增長趨勢較為接近,對照溫度下POD活性總體趨于穩定。由圖5可以看出,隨著溫度的降低,CAT的活性增加,對照溫度CAT活性變化比較穩定,在10 ℃時CAT活性的增加量較少,從第2天開始總體趨于穩定。0 ℃和5 ℃下CAT活性的變化趨勢比較明顯,增長較快,在第5天時比第1天時分別增長了78.45%和86.23%。

圖3 低溫脅迫對高山杜鵑SOD活性的影響
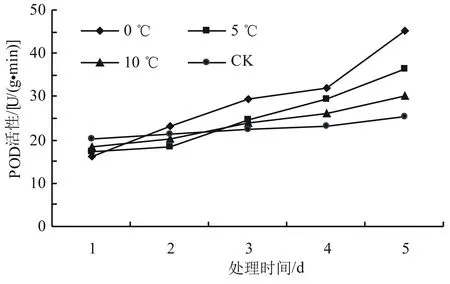
圖4 低溫脅迫對高山杜鵑POD活性的影響
2.4 低溫脅迫對高山杜鵑可溶性蛋白含量的影響
由圖6可知,在低溫脅迫下,隨著時間的延長,高山杜鵑幼苗的可溶性蛋白含量呈現上升趨勢。就同一天來說,隨著溫度的降低,可溶性蛋白含量呈現上升的趨勢,其中,低溫0 ℃處理下效果最為顯著(P<0.05),比對照增長了89.43%,在第2天至第3天上升最快,第3天時相對于對照組升高幅度最大,達86.51%。在10 ℃以下時,可溶性蛋白含量顯著上升(P<0.05),5 ℃和0 ℃分別比對照增長了84.40%和89.43%。在對照溫度下可溶性蛋白含量趨于穩定。

圖5 低溫脅迫對高山杜鵑CAT活性的影響
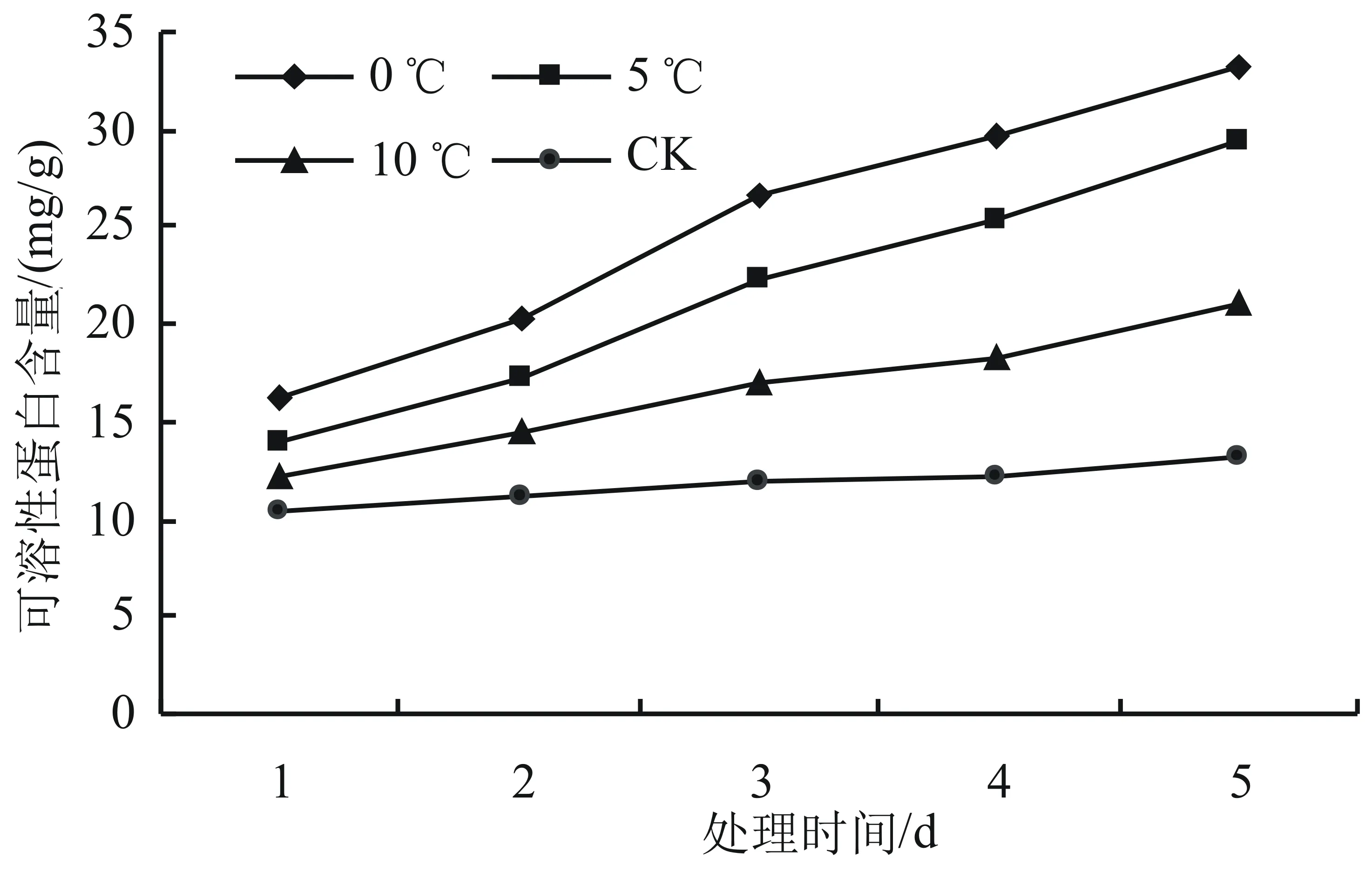
圖6 低溫脅迫對高山杜鵑可溶性蛋白含量的影響
2.5 低溫脅迫對高山杜鵑游離脯氨酸含量的影響
由圖7可知,低溫脅迫下,游離脯氨酸含量總體呈現上升的趨勢,但各溫度下存在一定的差異。隨著脅迫溫度的降低,高山杜鵑幼苗游離脯氨酸含量增加,但增加量不顯著。在相同溫度下,隨著脅迫時間的延長,游離脯氨酸含量緩慢增加,變化不明顯。低溫脅迫起始時,各低溫處理與對照組的游離脯氨酸含量水平較低,5 d后0、5、10 ℃處理組的游離脯氨酸含量分別上升至190.1、170.8、153.4 μg/g,與對照相比增加幅度分別為50.51%、35.23%和21.46%,其中0 ℃效果最明顯,與對照組差異最顯著(P<0.05)。
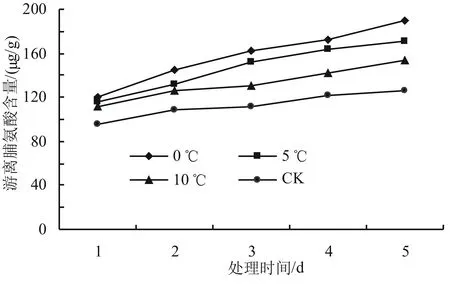
圖7 低溫脅迫對高山杜鵑游離脯氨酸含量的影響
3 討論
低溫作為植物生長發育的非生物脅迫因子之一,嚴重影響著植物體內的一系列生理生化過程,而植物則通過產生相應的抗逆性和耐逆性來適應外界環境的變化[15]。眾多植物抗寒研究報道指出,在低溫脅迫下,植物體內的可溶性蛋白含量、游離脯氨酸含量、保護酶活性等變化與植物抗寒性密切相關[16-18]。
細胞質膜是細胞最外面的一層界膜,對植物起到一定的保護作用,可以防止外界不良環境對植物組織結構的損傷。在低溫脅迫下,植物細胞膜的選擇透性遭到破壞,細胞內液外滲,進而導致相對電導率增大,且相對電導率值的大小與低溫脅迫程度呈正相關,與植物抗寒能力的強弱呈負相關[19],本試驗也進一步證實了此結論。高山杜鵑在對照溫度(25 ℃)時相對電導率變化較為穩定,隨著溫度的降低,相對電導率變化程度增大。本試驗中,在0 ℃時高山杜鵑相對電導率增長速度最快,同時也達到了最大值,原因是0 ℃低溫造成了高山杜鵑幼苗細胞膜和原生質體的損傷,而且其損傷程度會隨低溫脅迫時間的延長而加重,進而造成高山杜鵑膜結構的破壞及膜功能的喪失。
丙二醛(MDA)是膜脂過氧化作用的主要產物之一。植物體內MDA含量的大小可作為判斷膜系統損傷程度和植物抗逆性強弱的重要指標[20]。本試驗中,隨著低溫脅迫溫度的降低,高山杜鵑體內MDA含量呈上升趨勢,在0 ℃時MDA含量增加幅度最大;在相同處理溫度下隨著時間的延長MDA含量升高,在第5天時高山杜鵑MDA含量達到最大值,與前人的研究結果[21]一致。MDA的含量在低溫處理前兩天增長比較緩慢,與對照相差不大,但是隨著時間的延長,細胞膜脂質過氧化程度加深,導致膜結構和膜脂功能的損傷加劇,高山杜鵑體內MDA含量快速增長。
低溫脅迫下,植物體內會積累過多的超氧自由基,從而造成植物體內膜脂過氧化作用增強、生物膜透性增加等傷害。植物體內的天然超氧自由基清除劑,主要包括超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT),它們可以清除逆境下植物體內積累的自由基,從而維持自由基的動態平衡[22]。總體來說,高山杜鵑體內SOD活性表現為先上升后降低再上升的趨勢,低溫脅迫初期,高山杜鵑體內SOD活性顯著升高,隨著脅迫程度的加強,SOD活性開始逐漸下降,而在第4、5天時SOD活性又繼續上升。在整個試驗過程中,SOD活性隨著時間的延長而增加,在第5天時達到最大值。SOD活性隨著溫度的降低而升高,在0 ℃時增加幅度最大。過氧化物酶(POD)和過氧化氫酶(CAT)能夠提高植物對低溫環境的抗逆性,是因為POD和CAT能夠催化過氧化物分解,降低對細胞膜的傷害作用。隨著低溫脅迫時間的延長,高山杜鵑體內POD和CAT的活性總體上呈現出逐漸上升的趨勢。溫度越低,POD和CAT活性越高,在0 ℃處理下,雖然高山杜鵑體內SOD、POD和CAT活性都有了較為明顯的提高,但MDA含量的急劇上升仍會對高山杜鵑的正常生理代謝活動產生不良影響。而在5 ℃和10 ℃處理下,隨著低溫脅迫時間的延長,高山杜鵑體內MDA含量上升幅度較小,因此對植株的正常生長發育產生的傷害也較小。
可溶性蛋白在植物體內起著維持滲透調節及防止原生質體脫水的作用。在不同低溫脅迫條件下,植物體內可溶性蛋白含量的高低與植物的抗逆性強弱呈正比[23]。本試驗中,隨著低溫脅迫時間的延長高山杜鵑體內可溶性蛋白含量總體呈現上升趨勢,10 ℃時可溶性蛋白含量隨脅迫時間的延長增幅較小,0 ℃和5 ℃增幅較大,與熊燕等[23]的研究結果一致。
游離脯氨酸是植物體內重要的滲透調節物質之一。低溫脅迫導致了植物原生質體脫水從而引發游離脯氨酸的累積,游離脯氨酸含量的升高可降低冰點,有效防止細胞脫水,穩定原生質體及組織內的代謝過程,從而保護植物細胞免受低溫的傷害[24]。本試驗中,隨著低溫脅迫時間的延長,高山杜鵑體內游離脯氨酸含量逐漸升高;隨著脅迫溫度的降低,高山杜鵑體內游離脯氨酸含量升高,在0 ℃時達到最大值,但是總體變化不大。
綜上所述,秦嶺高山杜鵑幼苗在不同低溫脅迫條件下體內各項重要生理指標的變化結果表明,在5 ℃和10 ℃條件下其生長受到了不同程度的抑制,但仍具有一定抗性;0 ℃低溫條件下生長受到嚴重影響,不利于植株正常生長發育;在25 ℃下生長較好,但具體適于高山杜鵑生長的溫度還有待于進一步研究。
參考文獻:
[1] 李璟琦,蘇真,張曉靜.秦嶺杜鵑花屬植物資源及其利用研究[J].中國農學通報,2012,28(22):303-307.
[2] 鮮小林,陳睿.溫度與光強對高山杜鵑催花期間花芽營養物質積累的影響[J].西北植物學報,2015,35(5):991-997.
[3] 汪源,鞠波.中國野生杜鵑資源開發利用探討[J].生物學雜志,2006,23(1):43-44.
[4] 江福英,李延,翁伯琦.植物低溫脅迫及其抗性生理[J].福建農業學報,2002,17(3):190-195.
[5] 趙冰,杜宇科,付玉梅,等.鎮安木王國家森林公園野生杜鵑花資源調查[J]. Journal of Landscape Research, 2010, 2(1): 54-56,61.
[6] 李淑順,李倩中,唐玲,等.低溫對高山杜鵑內源營養動態變化及休眠解除的影響[J].江蘇農業學報,2011,27(4):853-857.
[7] 華智銳,李小玲.鹽旱交叉脅迫對小麥種子萌發及幼苗保護酶活性的影響[J].江西農業學報,2017,29(2):18-23.
[8] Mohamed H B, Vadel A M, Geuns J M C, et al. Biochemical changes in dormant grapevine shoot tissues in response to chilling: possible role in dormancy release[J]. Scientia Horticulturae(Amsterdam), 2010, 124(4): 440-447.
[9] 陳建勛,王曉峰.植物生理試驗指導[M].廣州:華南理工出版社,2013:78-80.
[10] 許維岸.生物學綜合實驗指導[M].蘇州:蘇州大學出版社,2010:115-116.
[11] 張治安.植物生理學實驗技術[M].長春:吉林大學出版社,2008:192-193.
[12] 李合生.植物生理生化實驗原理與技術[M].北京:高等教育出版社,2000:167-169.
[13] 郝再彬,蒼晶.植物生理實驗[M].哈爾濱:哈爾濱工業大學出版社,2004:110-112.
[14] 高俊鳳.植物生理學實驗指導[M].北京:高等教育出版社,2006:228-231.
[15] 楊東,張紅,陳麗萍,等.溫度脅迫對10種菊科雜草丙二醛和可溶性糖的影響[J].四川師范大學學報:自然科學版,2007,30(3):391-394.
[16] 李葉云,龐磊,陳啟文,等.低溫脅迫對茶樹葉片生理特性的影響[J].西北農林科技大學學報:自然科學版,2012,40(4):134-138,145.
[17] 張靜,朱為民.低溫脅迫下番茄細胞膜保護酶活性的變化[J].江蘇農業科學,2012,40(6):121-124.
[18] 許桂芳,張朝陽,向佐湘,等.利用隸屬函數法對4種珍珠菜屬植物的抗寒性綜合評價[J].西北林學院學報,2009,24(3):24-26.
[19] 田丹青,葛亞英,潘剛敏,等.低溫脅迫對3個紅掌品種葉片形態和生理特性的影響[J].園藝學報,2011,38(6):1173-1179.
[20] 吳海寧,羅興錄,樊吳靜.低溫脅迫對不同木薯品種幼苗生理特性的影響[J].南方農業學報,2013,44(11):1791-1799.
[21] 余麗玲,何天友,陳凌艷,等.人工低溫脅迫下西洋杜鵑生理生化指標的變化[J].農學學報,2014,4(1):48-53.
[22] 馬蘭濤,陳雙林,李迎春.低溫脅迫對Guaduaamplexfolia抗寒性生理指標的影響[J].林業科學研究,2008(2):235-238.
[23] 熊燕,曲彥婷,唐煥偉,等.低溫脅迫及不同基質對高山杜鵑生理和根系活力的影響[J].東北農業大學學報,2013,44(10):117-121.
[24] Siddig K, Ebert G, Ladders P. Tamarind (TamarindusindicaL.): A review on a multipurpose tree with promising future in the Sudan[J]. Angewandte Botanik, 2012, 73(5/6): 202-205.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
