NMHC-ⅡA在PRRSV感染M arc-145細胞過程中的作用
2018-05-05 07:18:27劉成倩孫鳳萍易建中
上海農業學報 2018年2期
關鍵詞:檢測
李 紅,劉成倩,孫鳳萍,高 駿,易建中*
(1上海市農業科學院畜牧獸醫研究所,上海 201106;2上海佳牧生物制品有限公司,上海 201106)
豬繁殖與呼吸綜合征(Porcine reproductive and respiratory syndrome,PRRS)是由PRRS病毒(PRRSV)感染所致的一種接觸性傳染病,主要表現為母豬發熱、厭食、流產、早產、木乃伊胎、弱仔等繁殖障礙及仔豬和豬呼吸系統疾病[1-3]。已公布的 PRRSV受體中,存在于豬肺泡巨噬細胞(Porcine alveolar macrophages,PAM)的 3個受體分別為硫酸乙酰肝素 (Heparin sulfate,HS)[4-5]、唾液酸黏附素(Sialoadhesin,Sn)[5-6]和清道夫受體 CD163[7],存在于 PRRSV易感染細胞系 Marc-145上的受體為 CD151和波形蛋白(Vimentin)[8-9]。目前,PRRSV與受體之間相互作用的分子機制還不完全清楚,普遍認為HS能夠調節低水平病毒的吸附,但并不能將病毒內吞。Sn能夠調節病毒的吸附和內吞,但并不能將病毒內吞,且不需要硫酸乙酰肝素的協助。CD163受體的作用可能是協助唾液酸黏附素內吞、PRRS病毒脫衣殼以及基因組RNA的釋放[10]。
存在于非肌肉細胞中的肌肉蛋白II稱為非肌肉肌球蛋白Ⅱ(Non-musclemyosinⅡ,NMⅡ),其與肌動蛋白結合構成細胞中的分子馬達[11-12]。研究發現,NMⅡ在囊泡的轉移和釋放、細胞吞飲、病毒入侵等方面也有一定的作用[13-14],但NMⅡ其他功能研究較少。非肌肉肌球蛋白重鏈Ⅱ型(Non-muscle myosin heavy chain-Ⅱ,NMHC-Ⅱ)廣泛分布在生物體內,參與細胞遷移、黏附、胞質分裂等各種生理活動[15]。NMHC-Ⅱ有NMHC-ⅡA、NMHC-ⅡB、NMHC-ⅡC 3種不同的亞型,它們具有相似的結構,均為由一對大小為171—244 ku的重鏈和兩對大小為16—23 ku的輕鏈組成的六聚體,其中NMHC-ⅡA的重鏈大小為230 ku[16]。2008年周恩民等[17]首次提出NMHC-ⅡA可能是PRRSV的受體或輔助因子,為了進一步研究該蛋白在PRRSV感染宿主細胞中的作用,本試驗以NMHC-IIA為研究對象,探討其與CD163受體的關系以及在PRRS病毒感染過程中的定位。
1 材料與方法
1.1 細胞、毒株、抗體以及試劑
Marc-145細胞系為本實驗室保存;PRRSV 12#毒株(GenBank:HQ416720)、抗PRRSV N蛋白的單克隆抗體(6D10)以及PRRSV陽性豬血清(345#)為山東農業大學免疫生物學實驗室提供;抗PRRSV GP5抗獨特型抗體Mab2-5G2由西北農林科技大學周恩民教授提供;二抗HRP-羊抗豬IgG和FITC-標記羊抗鼠IgG購自Jackson公司;G-cy5標記的羊抗鼠二抗購自SoutherBiotech公司;Vectashield包埋試劑購自Vector labs公司;其他常規試劑均為國產分析純。
1.2 免疫共沉淀提取NMHC-ⅡA
將Marc-145細胞從細胞培養瓶上輕輕刮下,3 000 r/min離心10min,收集細胞,PBS緩沖液(0.01mol/L,pH 7.2)重懸細胞,洗滌3次,使細胞達4×107個左右。加入RIPA細胞裂解液[18]1 mL重懸細胞,放入-70℃冰箱反復凍融裂解細胞3次,轉入4℃搖床上輕搖30 min,離心,收集上清液,并加入50μL用RIPA洗滌3次的Resin,放入4℃搖床上輕搖1 h,離心,收集上清,加入5μLMab2-5G2(11 mg/mL),4℃輕搖孵育14—16 h,加入Resin 50μL,4℃輕搖孵育4 h,離心,收集沉淀物,加入1 mL RIPA洗滌沉淀,離心,棄上清,洗滌3次。取Resin 10μL于試管中,進行SDS-PAGE檢測。
1.3 SDS-PAGE蛋白電泳檢測NMHC-ⅡA
參照分子克隆實驗指南操作制備8%分離膠和5%積層膠,進行凝膠染色和脫色[19]。按照電泳裝置的使用說明,裝好潔凈干燥的玻璃板,灌膠、電泳。
1.4 W estern Blot檢測NMHC-IIA
按照1.3 SDS-PAGE的步驟制備凝膠及電泳。將分離膠放入電轉緩沖液中,按照實驗室常規半干轉膜法轉PVDF膜[19]。取出的膜放入含2.5%脫脂奶粉的PBST中,封閉1 h。將膜放入含有Mab2-5G2的抗獨特型抗體中,孵育1 h。PBST洗滌3次,每次5 min。PVDF膜放入HRP標記羊抗鼠的二抗中孵育1 h。PBST洗滌3次,每次5 min。將膜放入底物中顯色,用去離子水終止反應。
1.5 NMHC-IIA與PRRSV的檢測
按照2×105個/mL含量培養Marc-145細胞于含有蓋玻片的細胞培養板內,24 h后,100μL細胞液接入100 TCID50的PRRSV,細胞板轉入4℃孵育1 h,然后放入37℃、5%CO2培養箱內培養不同時間,分別于 0 min、5 min、10 min、15 min、30 min、60 min、90 min、120 min用預冷的 75%乙醇 4℃固定 30 min,棄去細胞板內乙醇,超凈工作臺內吹干。固定細胞于0.5%TritonX-100,室溫處理5 min,PBS洗滌3次,每次5 min。加入阻斷緩沖液[18],室溫放置1 h,PBS洗滌3次,每次5min。細胞按照一定稀釋度加入Mab2-5G2(1 mg/mL)和PRRSV陽性豬血清345#(1∶160),室溫孵育1 h,PBS洗滌3次。將稀釋好的G-cy5標記的羊抗鼠和FITC標記的羊抗豬二抗(G-cy5-羊抗鼠IgG:1μg/106個細胞,FITC-羊抗豬IgG:1μg/106個細胞)加入到細胞上,室溫孵育1 h,PBS緩沖液洗滌3次。將細胞板內蓋玻片取出,將蓋玻片上細胞包埋在Vectashield包埋試劑中,用指甲油包封閉蓋玻片,于激光共聚焦顯微鏡下觀察、拍照。
1.6 NMHC-IIA與CD163片段的相互作用
將按照1.2方法提取的NMHC-IIA蛋白進行SDS-PAGE,然后轉入PVDF膜,將轉好的膜放于2.5%脫脂奶粉中4℃孵育過夜。膜用0.5%PBST洗滌3次,每次10 min,加入純化的M1、M2、M3蛋白[20],室溫孵育2 h,用0.5%PBST洗滌3次,每次10 min。加入Mab2-5G2或抗HIS標簽的單克隆抗體,室溫孵育2 h后,洗滌3次,每次10 min。加入羊抗鼠-HRP標記的二抗,室溫孵育1 h后,加入增強型HRP-DBA底物顯色液進行檢測。
2 結果與分析
2.1 NMHC-ⅡA蛋白的提取與檢測
通過免疫共沉淀技術獲得beads-5G2-NMHC-ⅡA蛋白復合物,將此蛋白復合物洗脫、中和,獲得純化的NMHC-ⅡA蛋白,取適量NMHC-ⅡA蛋白進行SDS-PAGE電泳。如圖1所示,NMHC-ⅡA蛋白大小為230 ku。
2.2 NMHC-IIA與PRRSV感染關系
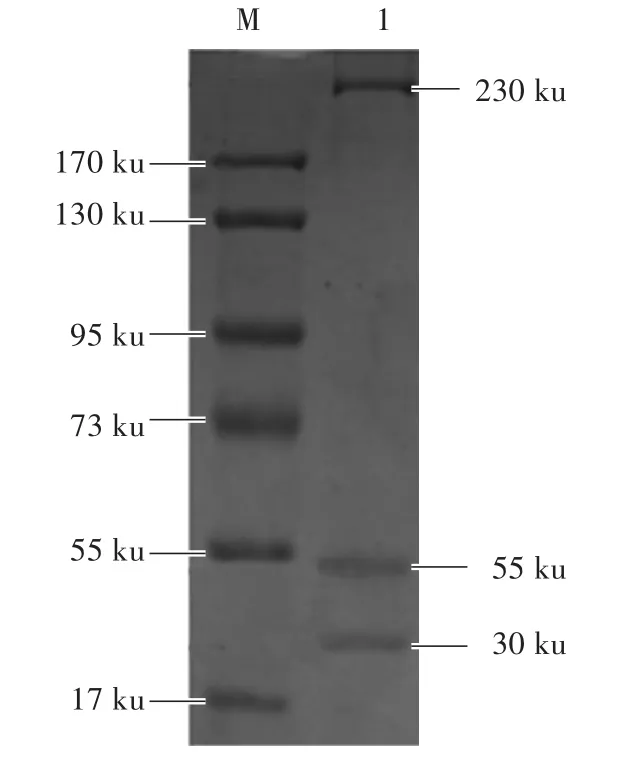
圖1 SDS-PAGE檢測M arc-145細胞中NMHC-IIAFig.1 Detection of NMHC-IIA in M arc-145 cells by SDS-PAGE
分別通過綠色和紅色熒光二抗(FITC-標記的羊抗豬和CYTM5-標記的羊抗鼠)檢測在PRRSV感染Marc-145細胞時,NMHC-ⅡA蛋白在細胞中的位置和表達量。通過激光共聚焦顯微鏡發現:將細胞轉入4℃時,PRRS病毒主要分布在被感染細胞表面,當細胞從4℃轉入37℃時,PRRS病毒在5 min左右進入細胞。隨著病毒感染細胞時間的延長,NMHC-ⅡA蛋白在Marc-145細胞內的表達量增加;當PRRS病毒完全進入被感染細胞后,NMHC-ⅡA的表達量減少(圖2)。
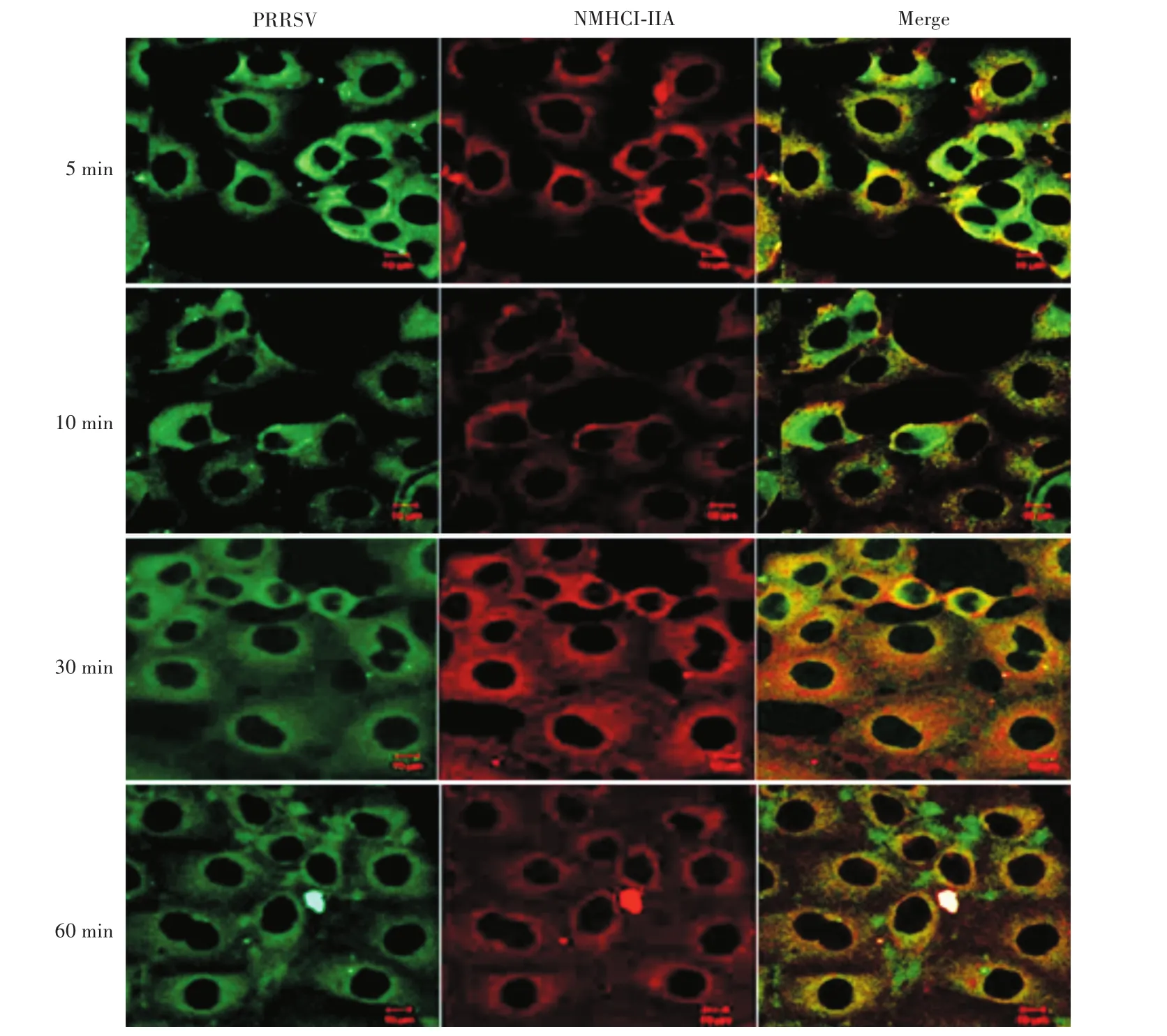
圖2 NMHC-ⅡA蛋白在PRRSV感染M arc-145細胞中的分布和表達量Fig.2 Distribution and expression of NMHC-ⅡA protein in M arc-145 cells infected with PRRSV
2.3 NMHC-ⅡA蛋白與CD163蛋白的相互作用
將純化的NMHC-ⅡA蛋白與CD163分段蛋白(M1、M2、M3)孵育1 h后,經免疫共沉淀試驗發現:NMHC-ⅡA蛋白與M1和M3蛋白能夠相互作用,與M2蛋白沒有相互作用(圖3)。
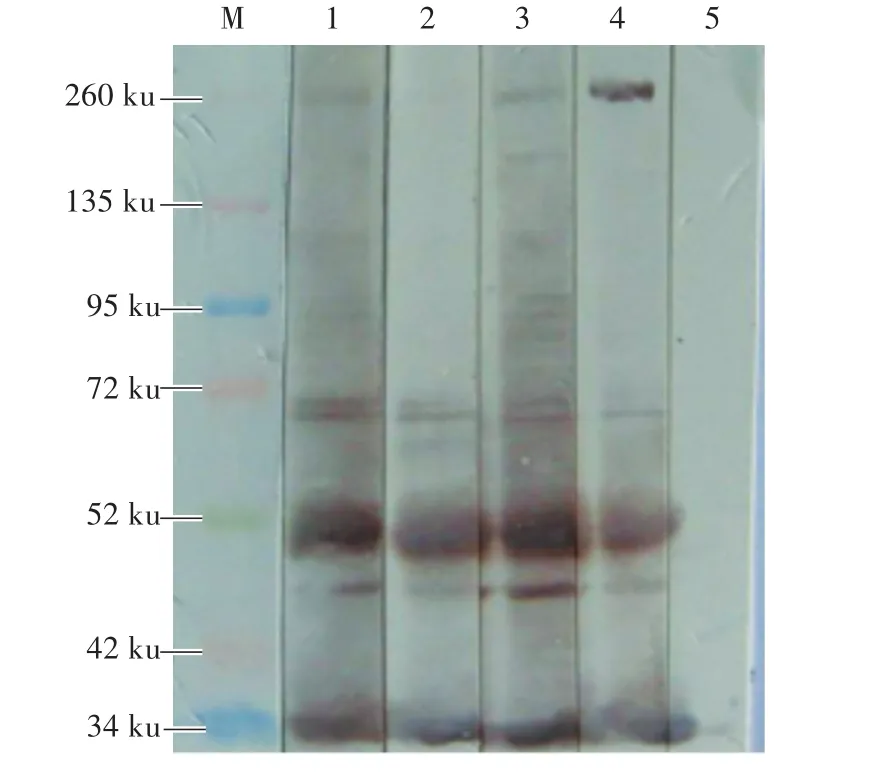
圖3 Western Blot檢測NMHC-ⅡA和CD163片段的相互作用Fig.3 Detection of interaction between NMHC-ⅡA and CD163 fragments by Western Blot
3 討論與結論
目前,PRRSV感染致病機制尚不清楚。病毒感染宿主細胞,主要是通過與細胞表面的特異性受體結合,利用細胞的內吞作用感染易感細胞。PRRSV感染易感細胞系非洲綠猴腎細胞Marc-145的過程也是通過細胞表面的受體進行的,但已有報道稱Marc-145細胞系上沒有Sn受體[21]。PRRSV能夠感染Marc-145細胞,說明PRRSV可能與Marc-145細胞上的其他受體或者輔助因子相互作用繼而引起PRRSV感染和進入細胞。Kim等[22]研究發現,波形蛋白存在于Marc-145細胞表面,并能夠與PRRSV的N蛋白相互作用,說明波形蛋白參與PRRSV感染Marc-145細胞的過程。周恩民等[17]通過免疫共沉淀技術,利用PRRSV結構蛋白GP5的單克隆抗獨特型抗體Mab2-5G2,從PAM和Marc-145細胞中提取獲得非肌肉肌球蛋白重鏈ⅡA(NMHC-ⅡA)。本研究發現,Marc-145細胞上的PRRSV受體CD163與NMHC-ⅡA蛋白能夠相互作用,作用位點主要集中在CD163蛋白的M1(20—259 aa)和M3(714—1 040 aa)蛋白上。同時,通過激光共聚焦顯微鏡定位發現,NMHC-ⅡA蛋白在PRRSV感染細胞過程中隨著感染時間的延長和病毒感染量的增加,其表達量也有所增加。本試驗認為,NMHC-ⅡA蛋白可能在PRRSV感染Marc-145細胞過程中發揮協同作用。本研究進一步揭示了PRRSV受體或協同因子之間的相互作用,對PRRSV感染致病機制有一定的促進作用。后期可以進一步縮短結合蛋白的長度,找到相互作用位點,利用RNAi技術敲除結合位點,從而減少病毒侵染宿主細胞的機會。
[1]WENSVOORT G.Lelystad virus and the porcine epidemic abortion and respiratory syndrome[J].Vet Res,993,24(2):117-124.
[2]MEULENBERG J J.PRRSV,the virus[J].Vet Res,2000,31(1):11-21.
[3]GREBENNIKOVA T V,CLOUSER D F,VORWALD A C,et al.Genomic characterization of virulent attenuated,and revertant passages of a North American porcine reproductive and respiratory syndrome virus strain[J].Virology,2004,321(2):383-390.
[4]DELPUTTE P L,VANDERHEIJDEN N,NAUWYNCK H J,et al.Involvement of thematrix protein in attachment of porcine reproductive and respiratory syndrome virus to a heparinlike receptor on porcine alveolarmacrophages[J].JVirol,2002,76(9):4312-4320.
[5]DELPUTTE P L,COSTERS S,NAUWYNCK H J.Analysis of porcine reproductive and respiratory syndrome virus attachment and internalization:distinctive roles for heparan sulphate and sialoadhesin[J].JGen Virol,2005,86(Pt5):1441-1445.
[6]VANDERHEIJDEN N,DELPUTTE P L,FAVOREEL H W,et al.Involvement of sialoadhesin in entry of porcine reproductive and respiratory syndrome virus into porcine alveolarmacrophages[J].JVirol,2003,77(15):8207-8215.
[7]CALVERT JG,SLADE D E,SHIELDS S L,et al.CD163 expression confers susceptibility to porcine reproductive and respiratory syndrome viruses[J].JVirol,2007,81(14):7371-7379.
[8]KIM JK,FAHAD A M,SHANMUKHAPPA K,et al.Defining the cellular target(s)of porcine reproductive and respiratory syndrome virus blockingmonoclonal antibody 7G10[J].JVirol,2006,80(2):689-696.
[9]SHANMUKHAPPA K,KIM JK,KAPIL S.Role of CD151,A tetraspanin,in porcine reproductive and respiratory syndrome virus infection[J].Virol J,2007,4:62.
[10]DELPUTTE P L,NAUWYNCK H J.Porcine artervirus entry in macrophage:heparin sulfate-mediated attachment,sialoadhesin-mediated internalization,and a cell-specific factormediating virus disassembly and genome release[J].Adv Exp Med Bio,2006,581:247-252.
[11]SIMONSM,WANG M,MCBRIDE O W,et al.Human nonmuscle myosin heavy chains are encoded by two genes located on different chromosomes[J].Circ Res,1991,69(2):530-539.
[12]BERG JS,POWELL B C,CHENEY R E.A millennialmyosin census[J].Mol Biol Cell,2001,12(4):780-794.
[13]SEABROOKE S,QIU X,STEWART B A.Nonmuscle myosinⅡ helps regulate synaptic vesicle mobility at the Drosophila neuromuscular junction[J].BMC Neurosci,2010,11:37.
[14]DOREIAN BW,FULOP T G,SMITH C B.MyosinⅡ activation and actin reorganization regulate the mode of quantal exocytosis in mouse adrenal chromaffin cells[J].JNeurosci,2008,28(17):4470-4478.
[15]VICENTE-MANZANARESM,MA X,ADELSTEIN R S,et al.Non-musclemyosinⅡ takes centre stage in cell adhesion and migration[J].Nat Rev Mol Cell Biol,2009,10(11):778-90.
[16]GOLOMB E,MA X,JANA SS,et al.Identification and characterization of nonmusclemyosin II-C,a new member of themyosin II family[J].J Biol Chem,2004,279(4):2800-2808.
[17]ZHOU E M,XIAO Y,SHIY,etal.Generation of internal imagemonoclonal anti-idiotypic antibodies against idiotypic antibodies to GP5 antigen of porcine reproductive and respiratory syndrome virus[J].JVirol Methods,2008,49(2):300-308.
[18]李紅.CD163及其他蛋白在PRRSV感染細胞過程中的相互作用[D].泰安:山東農業大學,2011.
[19]J薩姆布魯克,DW拉塞爾.分子克隆實驗指南[M].3版.北京:科學出版社,2002.
[20]李紅,劉成倩,易建中.PRRSV受體CD163與Marc-145細胞的相互作用[J].中國獸醫學報,2016,36(8)1273-1277.
[21]DUAN X,NAUWYNCK H J,FAVOREEL HW,etal.Identification of a putative receptor for porcine reproductive and respiratory syndrome virus on porcine alveolarmacrophages[J].JVirol,1998,72(5):4520-4523.
[22]KIM H S,KWANG J,YOON I J,et al.Enhance replication of porcine reproductive and respiratory syndrome(PRRSV)virus in a homogeneous subpopulation of MA-104 cell line[J].Arch Virol,1993,133(3/4):477-483.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
