磷素對小麥籽粒支鏈淀粉合成及其鏈長分布的影響
2018-05-04 02:35:45張潤琪付凱勇徐芳芳朱永琪覃安祥李春艷
麥類作物學報 2018年3期
張潤琪,李 誠,付凱勇,李 超,徐芳芳,朱永琪,覃安祥,李春艷
(石河子大學農學院/新疆兵團綠洲生態農業重點實驗室,新疆石河子 832000)
淀粉是小麥籽粒的主要組成成分,根據結構差異可分為直鏈淀粉和支鏈淀粉[1],其中支鏈淀粉占淀粉含量的70%~80%,其構型直接影響淀粉的理化特性和加工、食用品質[2]。支鏈淀粉的主鏈通過α-1,4-糖苷鍵連接而成,支鏈通過α-1,6-糖苷鍵與主鏈相連,可分為A、B和C三種鏈[3],其中,C鏈為主鏈,B鏈為C鏈的支鏈,而A鏈為B鏈的側鏈。前人將支鏈淀粉的色譜圖分為3 部分[4]:A鏈:DP 6~18,B1鏈:DP 19~34,B2鏈:DP>35。
淀粉的合成與結構不但與其遺傳特性有關,還受栽培過程中營養水平的影響[5]。磷素是作物必需的三大營養元素之一,施加磷肥是緩解土壤低磷脅迫、改善作物產量和品質的有效方法。腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)是淀粉合成的關鍵酶之一,在植物葉片中受變構調節,被 3-磷酸甘油酸激活而被無機磷酸所抑制,3-磷酸甘油酸與無機磷酸的比值影響其活性,從而調節淀粉的合成[6]。可溶性淀粉合成酶(SSS)、淀粉分支酶(SBE)和淀粉去分支酶(DBE)是負責支鏈淀粉合成的關鍵酶[7],分別由其對應的酶基因編碼[8]。其中,SBE的兩種同工酶均可被磷酸鹽激活[9]。磷素對淀粉的合成有重要影響。姜宗慶等[10]研究表明,在施磷(P2O5)量0 ~ 108 kg·hm-2范圍內,小麥籽粒直鏈淀粉、支鏈淀粉、總淀粉積累量、積累速率均隨施磷量的增加而上升。此外,磷對淀粉的品質特性也有顯著影響,Li等[11]研究表明,施磷顯著提高了小麥新冬20號籽粒淀粉中B型淀粉粒的比例,對其持水力、膨脹勢、溶解性等也有顯著影響。前人對于磷素影響小麥支鏈淀粉含量及支鏈淀粉合成關鍵酶活性已有研究,如Ni等[12]研究發現,施磷顯著提高了成熟期小麥籽粒中的支鏈淀粉含量,王旭東[13]的研究表明,施磷提高了花后28 d之前籽粒中蔗糖合成酶(SS)、顆粒淀粉合成酶(GBSS)和可溶性淀粉合成酶(SSS)的活性,提高了花后14~28 d 籽粒中AGPase的活性,但關于磷素影響小麥支鏈淀粉合成關鍵酶基因表達及鏈長分布的研究則鮮見報道。本研究選用不同冬小麥品種,測定不同磷素水平下淀粉合成關鍵酶基因表達量以及對應酶的活性、籽粒支鏈淀粉含量、支鏈淀粉鏈長分布和籽粒全磷含量,探究不同磷素水平對小麥籽粒支鏈淀粉合成及其鏈長分布的調控效應,以期揭示小麥支鏈淀粉的積累特征,為預測小麥淀粉品質提供理論依據。
1 材料與方法
1.1 試驗材料與設計
供試材料選用新冬20號(南疆主栽品種)和新冬23號(北疆強筋面包小麥),由石河子大學麥類作物研究所提供。
試驗于2014年10月至2015年6月在石河子大學農學院試驗站(44°17′N, 86°03′E)進行。前茬作物為向日葵,土質為灰漠土,0~20 cm 土層內含堿解氮63 mg·kg-1、速效磷15 mg·kg-1、速效鉀208 mg·kg-1。播種時按75 kg·hm-2施加尿素,灌溉方式為滴灌,冬前澆水3次,返青至成熟每隔10~12 d澆水一次,共澆水6次,每次滴水量1 125 m3·hm-2,總滴水量10 125 m3·hm-2。在拔節期、抽穗期和揚花期分別隨水施尿素45、75和120 kg·hm-2。
隨機區組設計,3個重復,小區面積2.4 m×3 m,小區之間隔離帶寬度為50 cm。設置3個磷素(P2O5)水平分別為0、105和210 kg·hm-2,分別用CK(對照)、CP(常規施磷)和HP(過量施磷)表示,所用肥料為重過磷酸鈣,在播種后160 d(大約5%的植株已返青)開溝條施。
1.2 試驗方法
1.2.1 樣品采集
從每個小區選取同日開花的麥穗做好標記,分別在花后7、14、21、28和35 d在每個小區剝取穗中部的籽粒,同處理籽粒集中到一起,一部分經液氮速凍5 min后保存于-80 ℃冰箱,用于測定酶活性和提取RNA;另一部分于105 ℃殺青30 min,80 ℃烘干至恒重備用。
1.2.2 測定項目與方法
(1)籽粒支鏈淀粉含量的測定 取烘干的小麥籽粒用實驗室粉碎磨(JFSD-70,上海嘉定)打成粉末,參照趙永亮[14]的方法測定。
(2)胚乳淀粉粒的提取 參照Peng等[15]的方法提取,常溫干燥后于-20 ℃保存。
(3)支鏈淀粉鏈長分布的測定 參照賀 偉等[16]的方法。
(4)籽粒全磷含量的測定 取烘干的小麥籽粒用實驗室粉碎磨(JFSD-70,上海嘉定)打成粉末,參照鮑士旦[17]的方法測定。
(5)支鏈淀粉合成相關酶活性的測定 胚乳SS酶活性的測定參照Douglas等[18]的方法;SBE酶活性的測定參照趙法茂等[19]的方法;DBE酶活性的測定參照劉 霞等[20]的方法。
1.2.3 支鏈淀粉合成相關酶基因相對表達量的測定
(1)引物設計 使用Primer Premier 5.0.軟件,根據NCBI上公布的小麥籽粒淀粉合成相關酶基因的序列,設計 sbe1, sbe2a, sbe2b和 iso1基因的引物(表1),由生工生物工程(上海)股份有限公司合成。使用小麥actin基因(NCBI編號DN551593)作為內參。通過梯度PCR驗證引物特異性同時優化PCR條件。
(2) RNA的提取和cDNA的合成 用Fruit-mate(Takara)和RNAiso plus(Takara)試劑盒提取RNA,按說明書操作,通過瓊脂糖凝膠電泳檢驗提取的總RNA質量。使用TIANScript cDNA第一鏈合成試劑盒(Tiangen, Cat#KR104-02, China)合成cDNA,用其作為模板擴增actin基因以檢驗其質量。
(3)實時熒光定量PCR 使用SYBR Premix ExTaq試劑盒(Takara),在實時熒光定量PCR儀(Roche LightCycler 480 Ⅱ)上檢測各個基因的擴增情況,具體操作按說明書進行。
1.3 數據處理
應用Excel 2003對數據進行統計分析和繪圖,用SPSS 11.5進行單因素方差分析,采用鄧肯法(Duncan)進行多重比較。
2 結果與分析
2.1 不同磷素處理對支鏈淀粉合成相關酶基因表達的影響
施磷顯著提高了兩個小麥品種花后14~28 d胚乳中 sbe1基因的相對表達量(圖1),且CP處理顯著高于HP處理(花后28 d除外)。新冬20號胚乳中 sbe1基因的相對表達量總體低于新冬23號。對照條件下,新冬20號胚乳中 sbe1基因相對表達量在花后28 d達到峰值,而新冬23號則在花后21 d達到峰值。CP處理下兩個參試品種 sbe1基因的相對表達量基本呈先上升后下降的趨勢,均在花后21 d達到峰值。HP處理下新冬20號胚乳中 sbe1基因的相對表達量呈雙峰分布,而新冬23號則呈單峰分布。花后35 d,隨著種子的成熟,兩品種各處理下胚乳中 sbe1基因的相對表達量降至最低值。
施磷顯著提高了兩參試品種花后14~28d胚乳中 sbe2a基因的相對表達量(圖2),且CP處理顯著高于HP處理。新冬20號胚乳中 sbe2a基因的相對表達量整體低于新冬23號。對照條件下兩品種胚乳中 sbe2a基因的相對表達量均較低;CP處理下新冬20號胚乳中 sbe2a基因的相對表達量在花后7 d最高,而新冬23號則在花后14 d達到峰值;HP處理下兩參試品種胚乳中 sbe2a基因的相對表達量均在花后14 d達到峰值。

A:新冬20;B:新冬23。圖柱上不同字母表示處理間差異在0.05水平顯著。下同。
A:Xindong 20;B:Xindong 23.Different letters above columns mean significant difference among treatments at 0.05 level. The same below.
圖1不同磷處理下小麥胚乳sbe1基因的相對表達量
Fig.1Relativeexpressionofsbe1inwheatendospermunderdifferentphosphoruslevels
如圖3所示,施磷可不同程度提高兩個小麥品種胚乳中 sbe2b基因的相對表達量。對照和CP處理下新冬20號胚乳中 sbe2b基因的相對表達量均在花后7 d最高,隨后不斷降低,而HP處理下則呈先上升后下降的趨勢,在花后14 d達到峰值。各處理下新冬23號胚乳中 sbe2b基因的相對表達量均呈先升后降之勢,都在花后14 d達到峰值。

圖2 不同磷處理下小麥胚乳 sbe2a基因的相對表達量

圖3 不同磷處理下小麥胚乳 sbe2b基因的相對表達量
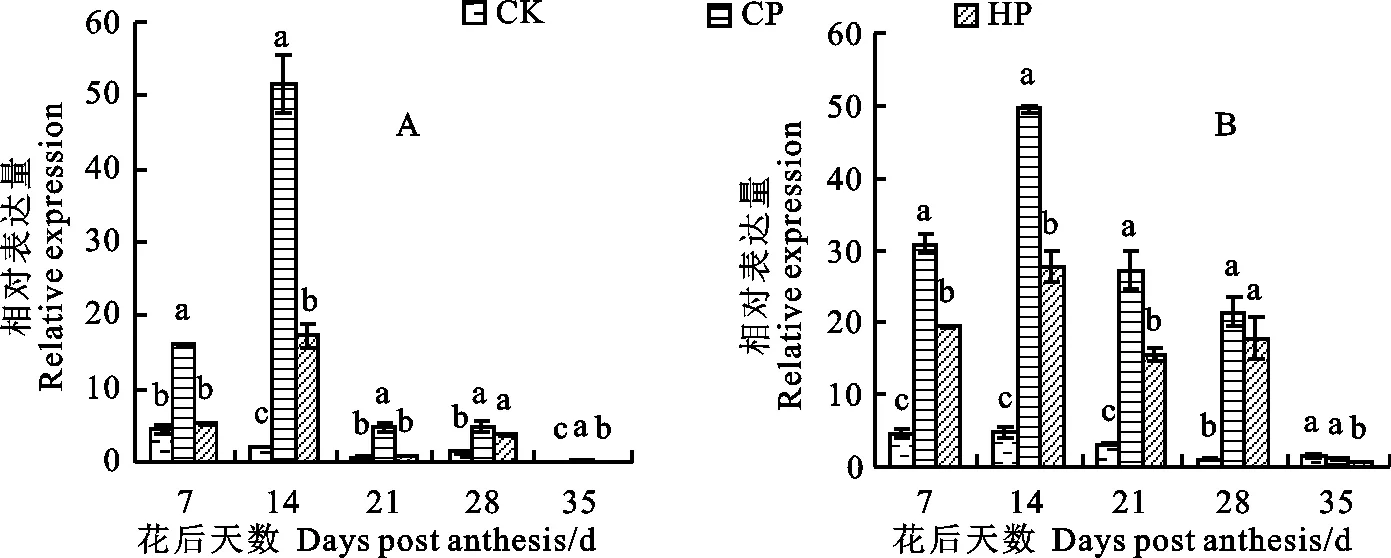
圖4 不同磷處理下小麥胚乳 iso1基因的相對表達量
如圖4所示,施磷顯著提高了兩個參試品種花后7~28 d胚乳中 iso1基因的相對表達量(新冬20花后7、28 d除外),且CP處理下顯著高于HP處理(花后28 d除外)。對照條件下兩參試品種胚乳中 iso1基因的相對表達量均較低,且各時期變化不大。CP和HP處理下兩品種胚乳中 iso1基因的相對表達量均呈先升后降之勢,都在花后14 d達到表達峰值;花后35 d,隨著灌漿的完成,兩品種胚乳中 iso1基因的相對表達量下降至最低。
2.2 不同磷處理對支鏈淀粉合成相關酶活性的影響
磷處理對小麥胚乳中SS酶活性的影響存在品種間差異(圖5),CP處理顯著提高了新冬20號胚乳中的SS酶活性(花后14 d除外),促進了淀粉合成原料蔗糖向籽粒的供應;除花后14 d外,HP處理下新冬20號籽粒發育各時期胚乳SS酶活性均顯著高于同期對照,花后35 d達到峰值;對照條件下胚乳中的SS酶活性呈升-降-升之勢,在花后14 d達到最大值。CP處理下新冬23號的胚乳SS酶活性呈升-降-升的趨勢,在花后35 d達到峰值且顯著高于同期HP和對照處理;HP處理下新冬23號的胚乳SS酶活性整體呈先升后降之勢,在花后21 d達到最大值且顯著高于同期對照和CP處理;對照條件下的胚乳SS酶活性呈降-升-降的趨勢,在花后28 d達到峰值。
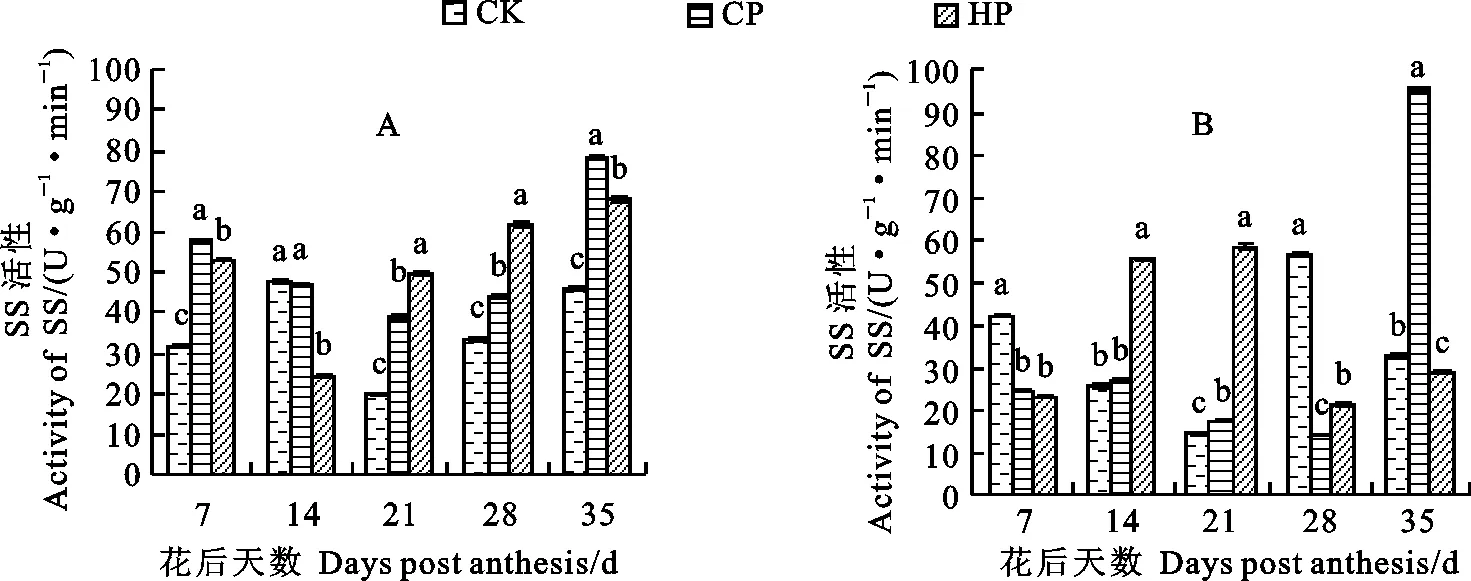
圖5 不同磷處理下的小麥胚乳SS酶活性
如圖6所示,新冬20號胚乳中的SBE酶活性CP處理下呈先升后降的趨勢,在花后14 d達到最大值且顯著高于同期HP和對照處理;在HP處理下呈升-降-升之勢,分別在花后14 d和35 d有最高峰和次高峰,且顯著高于同期對照處理;對照條件下胚乳SBE酶活性在籽粒灌漿期間逐漸下降。各處理下新冬23號胚乳中的SBE酶活性均在花后7 d酶活最高。

圖6 不同磷處理下的小麥胚乳SBE活性
如圖7所示,對照條件下新冬20號胚乳中的DBE酶活性呈降-升-降之勢,花后28 d達到最大值;CP和HP處理下新冬20號胚乳中的DBE酶活性均在花后7 d最高,且CP處理下顯著高于對照。相比于對照,施磷處理提前了新冬20號胚乳中DBE酶活性峰值的出現時間。對照和HP處理下新冬23號胚乳中的DBE酶活性均在花后7 d最高,之后則呈降-升-降的趨勢;CP處理下胚乳中的DBE酶活性呈升-降-升之勢,花后14 d達到最大值且顯著高于同期對照和HP處理。
2.3 不同磷處理對籽粒支鏈淀粉含量的影響
如圖8所示,兩品種籽粒中的支鏈淀粉含量在灌漿早期都很低,隨著籽粒的發育,其支鏈淀粉含量逐步提高。花后7 d時不同磷處理下的支鏈淀粉含量無顯著差異,在此之后施磷顯著或不顯著提高了籽粒中支鏈淀粉的含量,成熟期(花后35 d)籽粒中CP處理下的支鏈淀粉含量最高,且顯著高于對照和HP處理。
2.4 不同磷處理對籽粒支鏈淀粉鏈長分布的影響
由表2可以看出,CP處理提高了兩品種支鏈淀粉中B鏈所占的比例。新冬20號各處理下支鏈淀粉中B鏈所占比例隨籽粒的發育不斷上升,而新冬23號對照的支鏈淀粉中B鏈所占比例呈先下降后上升的趨勢,HP處理下呈先上升后下降的趨勢,CP處理下則在花后各時期變化不大。

圖7 不同磷處理下的小麥胚乳DBE活性

圖8 不同磷處理下的小麥籽粒支鏈淀粉含量
2.5 不同磷處理對籽粒中全磷含量的影響
如圖9所示,CP處理下,新冬20號花后7 d、21 d和28 d以及新冬23號花后21 d籽粒中全磷含量顯著高于對照,其他時期及HP處理與對照相比并無顯著差異,說明CP處理有提高籽粒全磷含量的趨勢,但并不明顯;花后35 d,隨著種子的成熟,兩參試品種各處理下籽粒中的全磷含量無顯著差異。
3 討 論
研究表明,低磷土壤(速效磷含量9.37~10.73 mg·kg-1)上隨著施磷量的增加,小麥胚乳 sss3、 sbe1基因相對表達量升高,中磷(速效磷含量19.54~20.71 mg·kg-1)、高磷土壤(速效磷含量37.46~38.77 mg·kg-1)施磷對 sss3、 sbe1基因相對表達量調節不顯著[21]。本研究表明,施磷可顯著提高小麥胚乳 sbe1, sbe2a, sbe2b和 iso1基因的相對表達量,且CP處理下更加顯著,這說明一定范圍內施加磷肥有利于支鏈淀粉合成相關酶基因的表達。Wang等[22]認為,SSS、SBE和DBE可能在轉錄水平上調節淀粉的合成,與本研究結果基本一致。王樹亮等[23]研究表明,隨著土壤磷水平的提高,小麥各器官含磷量均有提高(其中籽粒磷含量提高幅度最小),但磷素利用效率降低。姜宗慶[24]的研究表明,成熟期籽粒磷含量隨施磷量提高而增加,但施磷量超過一定水平則造成籽粒磷含量下降,與本研究的結果一致。由于AGPase與SBE酶活性均與磷素有關[8-9],因此,不同磷處理下籽粒磷含量的不同可能暗示了這兩種酶活性的變化。
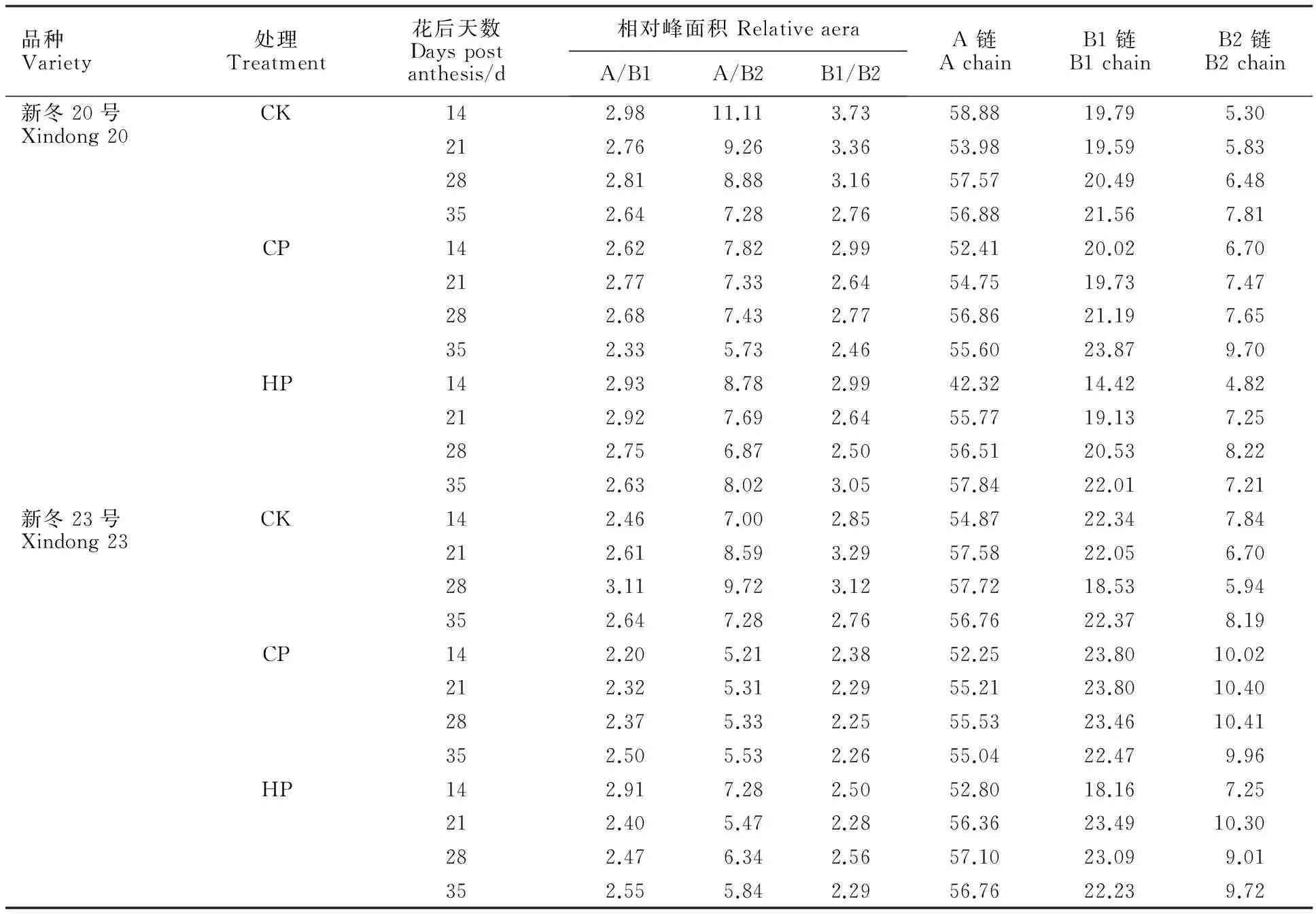
表2 不同磷水平下的小麥支鏈淀粉鏈長分布Table 2 Chain-length distribution of wheat amylopectin under different phosphorus levels %

圖9 不同磷處理下的小麥籽粒全磷含量
支鏈淀粉的合成是多種酶協同作用的結果。姜宗慶等[10,24]研究發現,在缺磷土壤(速效磷含量4.1 mg·kg-1)上增加施磷量可提高小麥籽粒SS酶的活性、籽粒蔗糖含量和SBE酶的活性。本研究發現,施磷提高了新冬20號胚乳SS酶活性和SBE酶活性峰值,且提前了新冬20號胚乳中DBE酶活性峰值的出現時間。說明磷處理促進了淀粉合成原料蔗糖在籽粒中的積累,為淀粉合成提供了充足的原料,同時通過提高酶活高峰期的SBE酶活性和DBE酶活性峰值的出現時間,提高了支鏈淀粉的合成速率,促進了支鏈淀粉的積累。這與本研究結果中支鏈淀粉合成相關酶基因表達量、支鏈淀粉含量的提高相符。此外,本研究使用的試驗材料新冬20號為南疆主栽弱冬性品種,早熟高產[25],全生育期較短(約238 d);而新冬23號則為北疆強冬性品種,中晚熟,中高產,全生育期較長(280~290 d)[26],品種特性差異可能是造成兩品種支鏈淀粉合成關鍵酶活性對磷素敏感性不同的原因。石河子大學冬小麥課題組的水培試驗亦表明,新冬20號根系比新冬23號更加發達(未發表),而根干重對植株磷素積累量有極顯著的直接作用[27],此外,新冬20號生育期雖然較新冬23號短,但其產量卻比新冬23號高,說明新冬20號對于磷素的吸收利用更加高效,因而其支鏈淀粉合成關鍵酶活性對磷素的敏感性相對較高。
本研究表明,適量施磷提高了支鏈淀粉中B鏈的比例,降低了A鏈的比例。這說明磷處理下支鏈淀粉的外側鏈較短,對長鏈的空間影響作用力較小,長鏈的運動性較強,有利于相互靠攏形成雙螺旋結構。前人研究表明,支鏈淀粉的鏈長分布對淀粉的品質特性有顯著的影響。DP 6~11支鏈與相對結晶度呈極顯著負相關,而DP 28~34支鏈與糊化溫度呈極顯著負相關[2];除崩解值外,長B鏈( DP≥65. 8)與黏度各特征值(峰值黏度、低谷黏度、最終黏度、反彈值、峰值時間)均呈極顯著正相關[28]。此外,Li等[11]的研究表明,施磷顯著提高了黏度特征值中的峰值黏度、低谷黏度、最終黏度和反彈值,顯著降低了糊化溫度,這與本研究中施磷提高B1鏈和B2鏈比例的結果相符。除長、短鏈外,內鏈、外鏈、平均鏈長、分支化度以及內外鏈比例對淀粉的結構也有影響,磷對淀粉品質特性的影響與這些因素的關系尚需深入研究。
本研究發現,磷素處理下兩個小麥品種支鏈淀粉合成相關酶基因( sbe1、 sbe2a、 sbe2b、 iso1)的表達量顯著上調,相關酶的活性也有所提高,從而增加了籽粒支鏈淀粉含量和支鏈淀粉B鏈段比例。不同磷素處理下支鏈淀粉含量與分子精細結構的變化可能還與其他淀粉合成酶基因表達量的變化、酶活性的改變等有關,其具體原因尚需進一步探究。
參考文獻:
[1]WEI L,WANG L,YANG Y,etal.Abscisic acid increases leaf starch content of polyethylene glycol-treated wheat seedlings by temporally increasing transcripts of genes encoding starch synthesis enzymes [J].ActaPhysiologiaePlantarum,2015,37(10):2.
[2] 賀曉鵬,朱昌蘭,劉玲瓏,等.不同水稻品種支鏈淀粉結構的差異及其與淀粉理化特性的關系[J].作物學報,2010,36(2):277.
HE X P,ZHU C L,LIU L L,etal.Difference of amylopectin structure among various rice genotypes differing in grain qualities and its relation to starch physicochemical properties [J].ActaAgronomicaSinica,2010,36(2):277.
[3] 李海普,李 彬,歐陽明,等.直鏈淀粉和支鏈淀粉的表征[J].食品科學,2010,31(11):275.
LI H P,LI B,OUYANG M,etal.Advances in characterization of amylose and amylopectin starch [J].FoodScience,2010,31(11):275.
[4] NAGAMINE T,KOMAE K.Improvement of a method for chain-length distribution analysis of wheat amylopectin [J].JournalofChromatographyA,1996,732(2):257.
[5] 梁曉芳,于振文.施鉀時期對冬小麥旗葉光合特性和籽粒淀粉積累的影響[J].應用生態學報,2004,15(8):1349.
LIANG X F,YU Z W.Effect of potassium application stage on photosynthetic characteristics of winter wheat flag leaves and on starch accumulation in wheat grains [J].ChineseJournalofAppliedEcology,2004,15(8)∶1349.
[6] MACDONALD P W,STROBEL G A.Adenosine diphosphate-glucose pyrophosphorylase control of starch accumulation in rust-infected wheat leaves [J].PlantPhysiology,1970,46(1):132.
[7] KALINGA D N,BERTOFT E,TETLOW I,etal.Evolution of amylopectin structure in developing wheat endosperm starch [J].CarbohydratePolymers,2014,112(2):316.
[8] RAHMAN S,LI Z,BATEY I,etal.Genetic alteration of starch functionality in wheat [J].JournalofCerealScience,2000,31:97,100.
[9] 高振宇,黃大年,錢 前.植物支鏈淀粉生物合成研究進展[J].植物生理與分子生物學學報,2004,30(5):490.
GAO Z Y,HUANG D N,QIAN Q.Advances in research on biosynthesis of plant amylopectin [J].JournalofPlantPhysiologyandMolecularBiology,2004,30(5):490.
[10] 姜宗慶,封超年,黃聯聯,等.施磷量對弱筋小麥揚麥9號籽粒淀粉合成和積累特性的調控效應[J].麥類作物學報,2006,26(6):81.
JIANG Z Q,FENG C N,HUANG L L,etal.Grain starch formation characteristics as affected by phosphorus application in weak-gluten wheat Yangmai 9 [J].JournalofTriticeaeCrops,2006,26(6):81.
[11] LI C Y,LI C,ZHANG R Q,etal. Effect of phosphorus on the characteristics of starch in winter wheat [J].Starch,2013,65:806.
[12] NI Y,WANG Z,YIN Y,etal.Starch granule size distribution in wheat grain in relation to phosphorus fertilization [J].JournalofAgriculturalScience,2012,150(1):45.
[13] 王旭東.磷對小麥產量和品質的影響及其生理基礎研究[D].泰安:山東農業大學,2003:51-53.
WANG X D.Effect of phosphorus on kernel yield and quality and physiological basis in winter wheat [D].Tai’an:Shandong Agricultural University,2003:51-53.
[14] 趙永亮.一種同時測定小麥種子中直鏈淀粉、總淀粉含量的新方法-微量分光光度法[J].食品與發酵工業,2005,31(8):24.
ZHAO Y L.A novel micro-amount method with spectrophotometer to determine amylose and total starch content of wheat seeds [J].FoodandFermentationIndustries,2005,31(8):24.
[15] PENG M,GAO M,ABDEL-AAL E S M,etal.Separation and characterization of A- and B-type starch granules in wheat endosperm [J].CerealChemistry,1999,76(3):375.
[16] 賀 偉,丁 卉,王婕琛,等.高效陰離子交換色譜-脈沖安培法測定支鏈淀粉糖鏈長分布[J].分析測試學報,2012,31(10):1243.
HE W,DING H,WANG J C,etal.Determination of chain-length distribution of amylopectin by high performance ion exchange chromatography with pulsed amperometric detection [J].JournalofInstrumentalAnalysis,2012,31(10):1243.
[17] 鮑士旦.土壤農化分析[M].北京:中國農業出版社,2000:268.
BAO S D.Soil and agro-chemistry analysis [M].Beijing:China Agriculture Press,2000:268.
[18] DOUGLAS C D,KUO T M,FELKER F C.Enzymes of sucrose and hexose metabolism in developing kernels of two inbreds of maize [J].PlantPhysiol,1988,86:1014.
[19] 趙法茂,齊 霞,肖 軍,等.測定淀粉分支酶活性方法的改進[J].植物生理學通訊,2007,43(6):1168.
ZHAO F M,QI X,XIAO J,etal.Improved method for determining starch branching enzyme activity [J].PlantPhysiologyJournal,2007,43(6):1168.
[20] 劉 霞,譚勝兵,田紀春,等.不同淀粉組分含量小麥品種灌漿過程中淀粉去分支酶的活性及類型[J].中國農業科學,2010,43(4):851.
LIU X,TAN S B,TIAN J C,etal.Activity and types of starch debranching enzyme in wheat cultivars containing different starch contents during the grain filling period [J].ScientiaAgriculturaSinica,2010,43(4):851.
[21] 陳 靜.小麥籽粒淀粉合成關鍵酶表達對磷素的響應[D].揚州:揚州大學,2010:33.
CHEN J.Expressions starch synthase response in wheat grain to phosphorus [D].Yangzhou:Yangzhou University,2010:33.
[22] WANG Z B,LI W H,QI J C,etal.Starch accumulation,activities of key enzyme and gene expression in starch synthesis of wheat endosperm with different starch contents [J].JournalofFoodScienceandTechnology,2011,51(3):419.
[23] 王樹亮,田奇卓,李娜娜,等.不同小麥品種對磷素吸收利用的差異[J].麥類作物學報,2008,28(3):476.
WANG S L,TIAN Q Z,LI N N,etal.Differences of phosphorus utilization efficiency among different wheat varieties [J].JournalofTriticeaeCrops,2008,28(3):476.
[24] 姜宗慶.磷素對小麥產量和品質的調控效應及其生理機制[D].揚州:揚州大學,2006:5.
JIANG Z Q.Effects of phosphorus on yield and quality formation and physiological mechanism in wheat [D].Yangzhou:Yangzhou University,2006:5.
[25] 陳淑琴,路香彩,張耀泉,等.適宜兩熟制種植的早熟冬小麥新品種冀麥37號的選育及應用[J].河北農業科學,1998,2(2):1.
CHEN S Q,LU X C,ZHANG Y Q,etal.Breeding and application of new early-variety Jimai 37 which fit for double-crop system area [J].JournalofHebeiAgriculturalScience,1998,2(2):1.
[26] 姚翠琴,蘇敬國,李新杰,等.優質面包小麥新冬 23號品質分析及應用前景[J].種子,2010,29(5):92.
YAO C Q,SU J G,LI X J,etal.Quality analysis and application prospect of superior bread wheat Xindong 23 [J].Seed,2010,29(5):92.
[27] 李春艷,馬 龍,張 宏,等.新疆冬小麥苗期耐低磷指標的篩選[J].麥類作物學報.2013,33(1):137.
LI C Y,MA L,ZHANG H,etal.Screening on indexes for tolerance to phosphorus stress at seedling stage of winter wheat [J].JournalofTriticeaeCrops,2013,33(1):137.
[28] 李春燕,封超年,王亞雷,等.不同小麥品種支鏈淀粉鏈長分配及其與淀粉理化特性的關系[J].作物學報,2007,33(8):1240.
LI C Y,FENG C N,WANG Y L,etal.Chain length distribution of debranched amylopectin and its relationship with physicochemical properties of starch in different wheat cultivars [J].ActaAgronomicaSinica,2007,33(8):1240.
