菠蘿混合授粉雜交品系表型研究及其父本來源分析
2018-04-27 05:14:57劉傳和
農學學報 2018年4期
劉傳和,劉 巖
(1廣東省農業科學院果樹研究所,廣州510640;2農業部南亞熱帶果樹生物學與遺傳資源利用重點實驗室,廣州510640;3廣東省熱帶亞熱帶果樹研究重點實驗室,廣州510640)
0 引言
菠蘿[Ananas comosus(L.)Merr.]又稱鳳梨,屬鳳梨科(Bromeliaceae)鳳梨屬(Ananas)草本果樹,原產于巴西、阿根廷及巴拉圭一帶的熱帶地區,是著名的熱帶水果。中國菠蘿品種資源有限、栽培品種結構單一,目前主栽品種仍為‘巴厘’,其次是‘無刺卡因’,兩者約占中國栽培菠蘿的90%。‘神灣’菠蘿雖僅主栽于廣東省中山市,但因其香味濃郁、品質優越而深受消費者的喜愛。
雜交育種是種質創新、獲得植物新品種的有效途徑。菠蘿自花不育,在雜交授粉時不需去雄套袋,這為菠蘿雜交育種創造了方便。混合授粉作為一種有性雜交方式,具有操作簡便,花粉來源廣泛,后代變異豐富而有利于選擇等特點[1],能顯著提高雜交的可孕性及選種效率。菠蘿‘H’(暫定名)是由‘巴厘’與‘神灣’菠蘿的混合花粉授到‘無刺卡因’上而獲得的雜交品系,其果實大小適中、產量穩定,果肉爽脆、香甜。
但由于混合花粉授粉雜交后代的父本未知,在進一步安排品種比較試驗時有困難[2]。現代生物技術分子標記的應用為混合授粉雜交后代的父本鑒定提供了有效方法[2-3]。然而植物表型分析因其適應意義和進化意義,仍不失為研究物種變異和進化的有效傳統方法[4]。基于傳統分類學與計算技術結合的數量分類學,依據表型特征對植物進行分析,將定性描述提升為定量的綜合分析,可以再現研究對象之間的親疏關系,使基于表型特征的研究更加高效[5-6]。如基于歐氏距離(Euclidean distance)的表型性狀遺傳相似性分析已廣泛用于雜種后代選擇[7-8]、種質資源遺傳多樣性分析[5,9-10]。
而基于限制性內切酶酶切的SLAF-seq(specific length amplified fragment sequencing)技術通過生物信息學進行試驗方案系統設計,構建SLAF-seq文庫,篩選特異長度DNA片段,以高通量測序技術獲得海量序列,利用軟件進行數據分析比對,獲得大量特異DNA片段,進而依據其序列發展出特異分子標記。SLAF-seq技術可用于單體型圖譜、遺傳圖譜、關聯性圖譜、多態性圖譜的構建,為分子育種、系統進化、種質資源鑒定提供重要技術保障[11-13]。
本研究以混合授粉雜交菠蘿品系‘H’與其母本‘無刺卡因’,以及混合花粉供體‘巴厘’、‘神灣’為材料,通過田間試驗調查物候期,比較研究植株、果實大小及果實品質、色澤、香氣物質組成等;基于以上性狀分析‘H’與其母本以及混合花粉供體‘巴厘’、‘神灣’間的遺傳相似性,并通過SLAF-seq技術分析了‘H’的父本可能來源。本研究旨在從表型及分子水平為混合授粉雜交品系‘H’父本來源進行分析,為菠蘿雜交育種的親本選擇提供參考。
1 材料與方法
1.1 試驗地描述
本試驗在位于廣州市天河區(113.35°N,23.12°E)的廣東省農業科學院果樹研究所試驗果園內進行。試驗地土壤為沙壤土;所在區域為南亞熱帶季風性氣候,年平均降雨量1725 mm,年平均氣溫21.5~22.2℃;最高溫出現在7—8月,28~35℃;最低溫出現在12月—次年2月,平均氣溫10~15℃。
1.2 試驗材料
本試驗所用材料為菠蘿吸芽苗,苗長約35 cm;包括雜交品系‘H’及其母本‘無刺卡因’,以及混合花粉供體品種‘巴厘’、‘神灣’。
1.3 試驗設計
本試驗采用隨機區組試驗,試驗地由14個東西長6 m、南北寬1 m的小區組成,小區間隔0.7 m,平行排列。除兩端各留出1個小區種植非供試菠蘿品種作為保護行外,其余12個小區分別種植供試品種(系)‘H’及‘無刺卡因’、‘巴厘’、‘神灣’,每個品種(系)各種植3個小區,隨機排列。于2013年3月中旬種植,每個小區東西走向雙行種植,株行距30 cm×30 cm。果園的其他管理一致。
1.4 表型性狀觀察與測定
分別記錄各個品種(系)的在自然條件下的植株初始抽蕾、開花的時間,果實初始成熟時間(果實黃熟,底部1/3果眼轉黃)。植株自然抽蕾時用直尺測定植株的株高,最長葉片長度、寬度;統計自然抽蕾時的植株葉片數;每個小區測定5株。果實成熟時,從每個小區隨機采收6個果實用于品質等相關性狀分析。果實可溶性固形物含量用手持式折光儀測定;果實蔗糖、還原糖、全糖、可滴定酸、維生素C等參考仝月澳和周厚基[14]的方法測定;果實硬度用GY-1硬度計測定。果實色澤用SP60色差計(X-rite,美國產)測定。果實香氣物質通過固相頂空微萃取,用GC-MS聯用儀分析測定[15]。
1.5 遺傳相似性計算
分別基于物候期、植株大小、果實大小,果實理化品質、色澤、香氣物質等6類指標參數,采用SPSS Statistics17.0軟件對雜交品系‘H’與其母本‘無刺卡因’,以及混合花粉供體‘巴厘’、‘神灣’間的遺傳距離(不相似性系數)進行計算,比較‘H’與‘無刺卡因’、‘巴厘’、‘神灣’間的遺傳相似性[16]。遺傳距離(不相似性系數)越小,表示遺傳相似性越高。在基于物候期的遺傳距離計算中,將最早抽蕾、開花、成熟的品種(系)定為0,相比較遲n天的則其值記為n。
1.6 基于SLAF-seq的父本來源分析
基于SLAF測序分析‘H’的父本來源,通過母本‘無刺卡因’和子代‘H’的基因型,根據子代‘H’基因型來源于‘巴厘’、‘神灣’的比例來判斷哪個樣本作為父本的可能性更高。首先,過濾掉子代或待驗證父本是缺失基因型以及2個待驗證父本基因型相同的位點;再過濾掉2個待驗證父本都能提供的基因型位點,留下只能由其中1個父本提供基因型的位點,然后判斷子代基因型的來源。
1.7 統計分析
采用SPSS Statistics17.0軟件進行數據差異顯著性分析(鄧肯氏新復極差法測驗,n≥3)。
2 結果與分析
2.1 主要物候期
表1所列為菠蘿雜交品系‘H’與其母本‘無刺卡因’,以及混合花粉供體‘巴厘’、‘神灣’植株自然條件下的初始抽蕾、開花以及果實成熟時間。雜交品系‘H’植株的自然抽蕾時間與其母本‘無刺卡因’一樣,都在4月3日開始抽蕾,介于‘巴厘’與‘神灣’之間。‘H’的初始開花時間比其母本‘無刺卡因’略早,與混合授粉花粉供體之一‘神灣’一樣,而比‘巴厘’早。‘H’的果實成熟時間早于其母本‘無刺卡因’,而晚于混合花粉供體‘巴厘’及‘神灣’,但與‘神灣’更接近。
2.2 植株大小
表2所列為菠蘿自然抽蕾時的植株大小。雜交品系‘H’自然抽蕾時株高與其母本‘無刺卡因’間無顯著差異。在混合花粉供體‘巴厘’與‘神灣’中,‘H’的株高比‘神灣’略高(差異不顯著),而顯著低于‘巴厘’。‘H’的葉長低于其母本‘無刺卡因’及混合花粉供體‘巴厘’、‘神灣’,其中與‘巴厘’間的差異達到顯著水平。‘H’葉寬與‘無刺卡因’及‘巴厘’、‘神灣’間均無顯著差異,但與‘無刺卡因’及‘神灣’更接近。‘H’的葉片數顯著多于其母本‘無刺卡因’;在‘巴厘’與‘神灣’中,‘H’的葉片數顯著多于‘神灣’,而與‘巴厘’差異不顯著。
2.3 果實大小
表3所列為菠蘿果實成熟時的縱橫徑、單果重、果心寬及頂芽長度。雜交品系‘H’的縱徑顯著低于其母本‘無刺卡因’;在混合花粉供體‘巴厘’與‘神灣’中,‘H’的縱徑略高于‘神灣’,而顯著低于‘巴厘’。‘H’的橫徑顯著低于‘無刺卡因’,與‘巴厘’無顯著差異,但顯著高于‘神灣’。‘H’的單果重顯著低于其母本‘無刺卡因’及花粉供體之一‘巴厘’;而顯著高于‘神灣’。‘H’的果心寬與其母本‘無刺卡因’以及‘神灣’間無顯著差異,但顯著低于‘巴厘’。‘H’的頂芽長顯著低于其母本‘無刺卡因’;但顯著高于‘巴厘’及‘神灣’,與‘神灣’更接近。
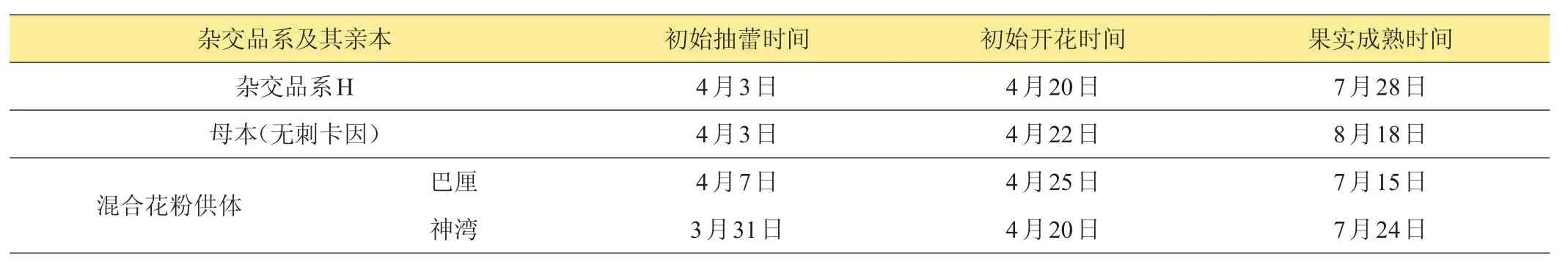
表1 菠蘿雜交品系‘H’及其親本的主要物候期比較
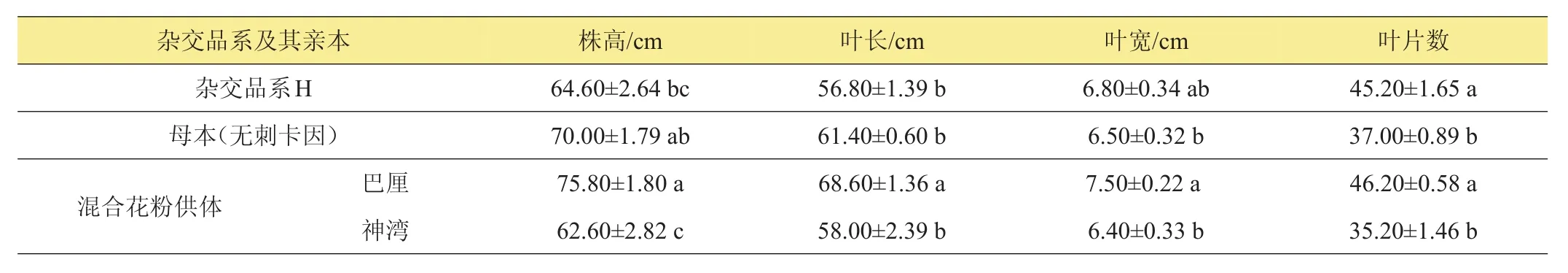
表2 菠蘿雜交品系‘H’及其親本自然抽蕾時植株大小
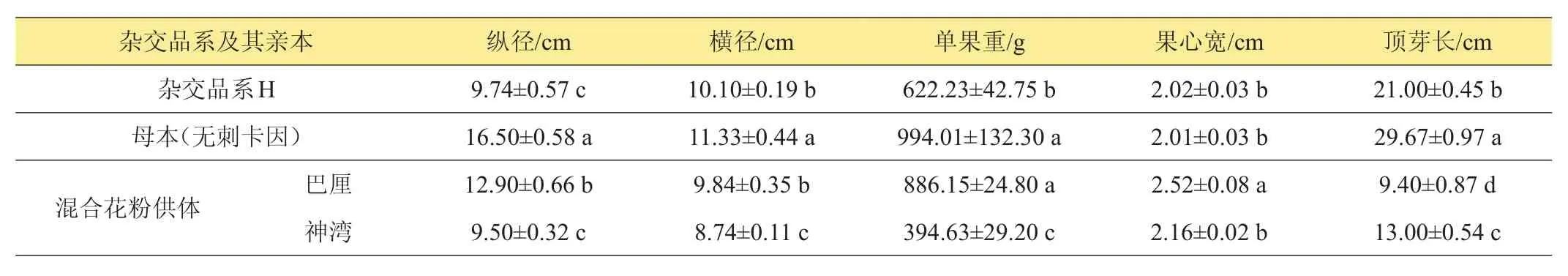
表3 菠蘿雜交品系‘H’及其親本果實大小
2.4 果實理化品質
表4所示為菠蘿果實成熟時的理化品質。‘H’的可溶性固形物、蔗糖、全糖、維生素C含量以及硬度等均顯著高于其母本‘無刺卡因’以及混合花粉供體‘巴厘’、‘神灣’,且在數值上均與‘神灣’更接近。‘H’的還原糖含量顯著低于其母本‘無刺卡因’,而與‘巴厘’、‘神灣’無顯著差異。‘H’的可滴定酸含量低于其母本‘無刺卡因’及‘巴厘’、‘神灣’,其中與‘神灣’間的差異達到顯著水平。‘H’的維生素C含量顯著高于‘無刺卡因’以及‘巴厘’、‘神灣’。‘H’的硬度也顯著高于‘無刺卡因’以及‘巴厘’、‘神灣’。
2.5 果實色澤
表5所列為菠蘿果實成熟時的果皮、果肉色澤指標。‘H’果皮的L*值與其母本‘無刺卡因’以及混合花粉供體‘巴厘’、‘神灣’間均無顯著差異。‘H’果皮的a*、b*值均顯著低于‘無刺卡因’,而與‘巴厘’、‘神灣’間無顯著差異。
‘H’果肉的L*、a*及b*值均顯著低于其母本‘無刺卡因’,而與混合花粉供體‘巴厘’、‘神灣’間無顯著差異。
2.6 果實香氣物質
表6列出了雜交品系‘H’與其母本‘無刺卡因’,以及混合花粉供體‘巴厘’、‘神灣’菠蘿果實中檢測到的香氣物質組分及其相對含量。在‘H’中檢測出4種酯類及7種烯類香氣物質,其總相對含量分別為47.60%及32.47%。在‘無刺卡因’中,檢測出2種酯類香氣物質及5種烯類香氣物質,酯類、烯類香氣物質總相對含量分別為44.57%及37.61%。在‘巴厘’果實中檢測出5種酯類香氣物質及3種烯類香氣物質;酯類香氣物質總相對含量為42.71%,烯類香氣物質總相對含量為4.68%。在‘神灣’的果實中檢測出酯類香氣物質5種,烯類香氣物質3種,酯類、烯類香氣物質總相對含量分別為75.62%及6.59%。
從檢測出的香氣物質分類統計可以看出,檢測出的香氣物質組分最多的是雜交品系‘H’,其次是混合花粉供體‘巴厘’及‘神灣’,而‘H’的母本‘無刺卡因’中檢測出的香氣物質組分數最少(表7)。從香氣物質的分類可以看出,雜交品系‘H’檢測到的酯類香氣物質總含量略高于其母本‘無刺卡因’以及‘巴厘’,而明顯低于‘神灣’。‘H’的烯類香氣物質總含量略低于‘無刺卡因’,而明顯高于‘巴厘’及‘神灣’。而‘H’的酯類、烯類香氣物質總含量略低于其母本‘無刺卡因’以及‘神灣’,而明顯高于‘巴厘’。
2.7 雜交品系H與其親本間的遺傳相似性
表8所示為基于主要表型性狀的‘H’與其親本間的遺傳距離。基于物候期,植株大小,果實大小、理化品質、色澤、香氣物質6類主要表型性狀分析,雜交品系‘H’與其母本‘無刺卡因’間的遺傳距離分別為12.166、10.847、371.945、9.120、74.219及8.003。‘H’與混合花粉供體之一‘巴厘’間的遺傳距離分別為14.491、16.315、264.194、8.497、4.758 及 43.476;而與‘神灣’間的遺傳距離分別為5.000、10.276、227.745、5.832、2.799及38.580。在‘巴厘’、‘神灣’2個混合花粉供體中,物候期、植株大小、果實大小、理化品質、色澤、香氣物質6類表型性狀分析均表明‘H’與‘神灣’間的遺傳距離相對更近,遺傳相似性更高。
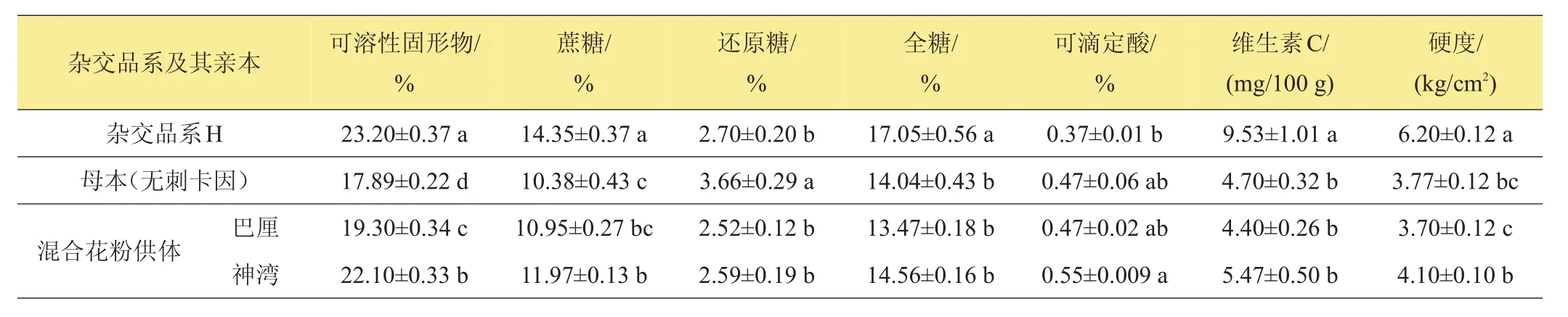
表4 菠蘿雜交品系及其親本果實理化品質
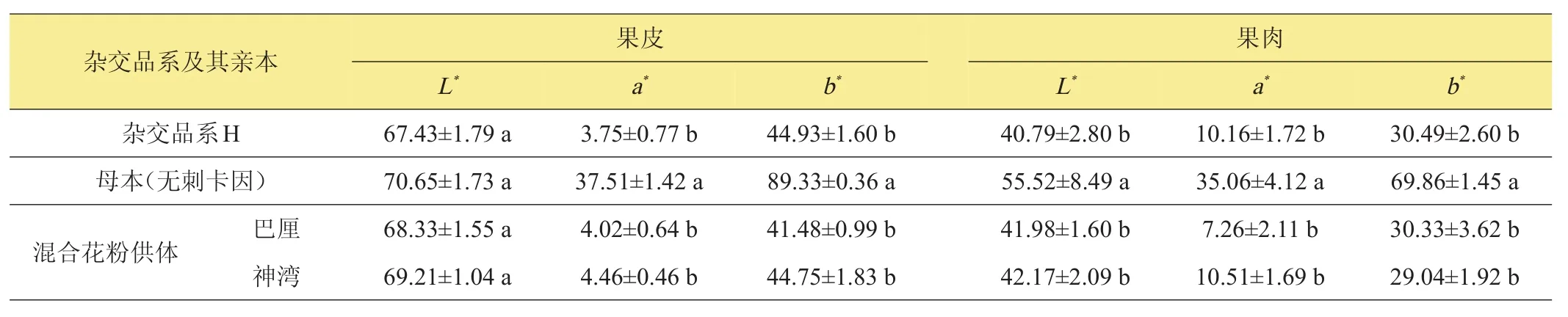
表5 菠蘿雜交品系及其親本果實色澤
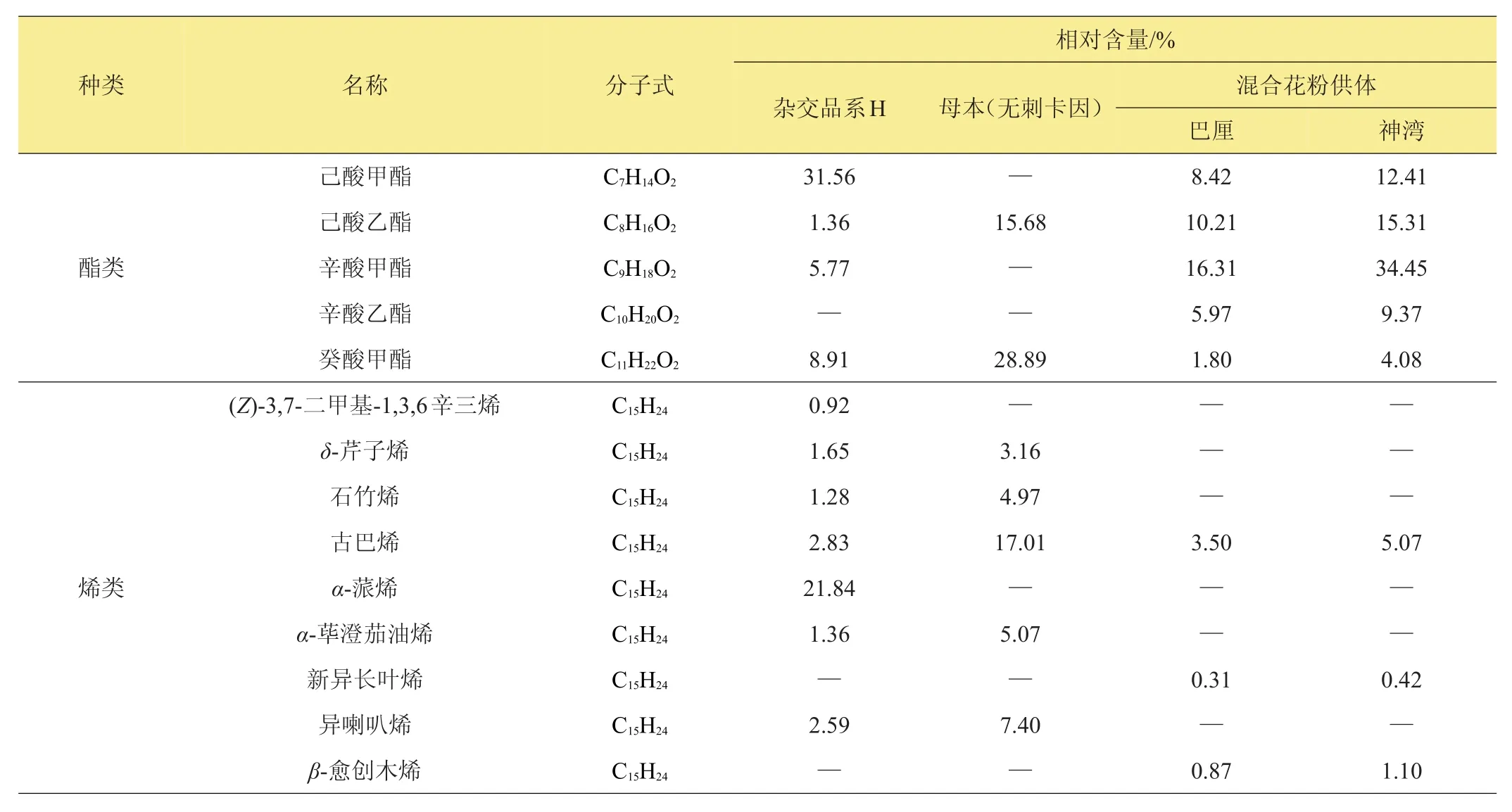
表6 菠蘿雜交品系及其親本果實香氣物質組分及含量
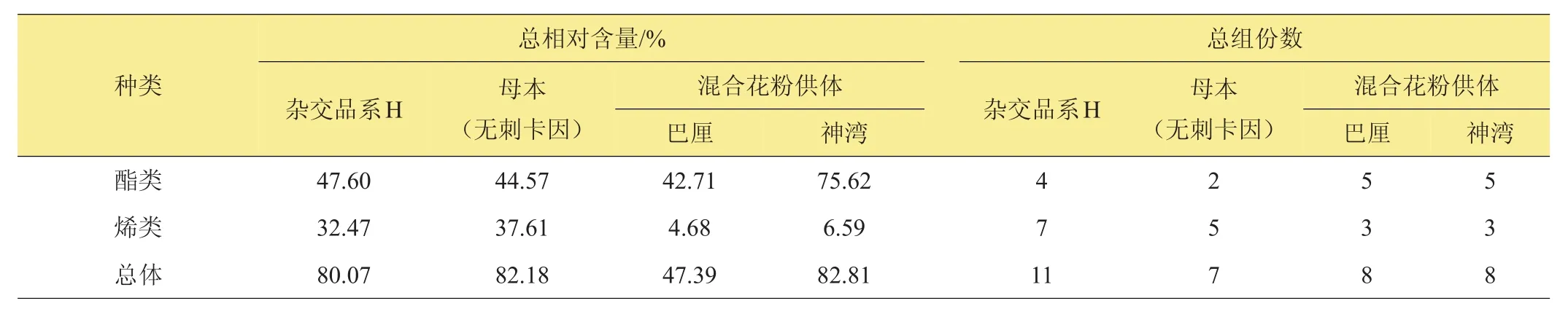
表7 菠蘿雜交品系及其親本果實香氣物質種類及總含量
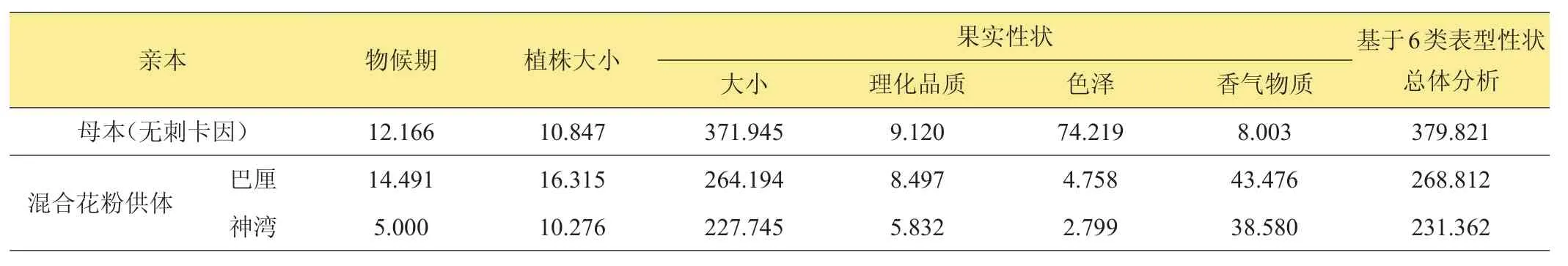
表8 基于表型性狀的雜交品系H與其親本間遺傳距離分析
基于物候期、植株大小、果實大小,果實品質、色澤、香氣物質前述6類指標參數共同綜合分析,雜交品系‘H’與其母本‘無刺卡因’間的遺傳距離最近為379.821;在‘巴厘’、‘神灣’2個混合花粉供體中,‘H’與‘神灣’的遺傳距離相對更近,為231.362,遺傳相似性更高。
2.8 基于SLAF-seq的雜交品系H父本來源分析
共獲得253506個SLAF標簽,標簽平均測序深度為76.59x,其中,多態性SLAF標簽有80353個,共獲得303779個SNP標記(表9)。根據SLAF-seq分析可知待驗證父本‘巴厘’、‘神灣’的親緣關系比較近。在2個混合花粉供體中,根據親本判斷子代基因型的異常率及子代SNP基因型的來源分析表明‘H’的父本來源于‘神灣’的可能性高于‘巴厘’(表10)。
3 討論與結論
據不完全統計,全世界約有60~70個栽培菠蘿品種[17]。近年來不少雜交菠蘿品種被選育出來,如美國Del Monte所選育的‘MD-2’,波多黎各大學農業試驗站選出的‘P.R.1-67’,中國臺灣省選育出的‘臺農’系列品種等都是雜交菠蘿品種。以‘無刺卡因’為母本,‘神灣’為父本雜交選育出的‘粵脆’菠蘿是中國大陸地區選育出的第一個雜交品種。竇美安等[18]也以‘無刺卡因’和‘巴厘’為親本雜交獲得了多個F1群體單株。
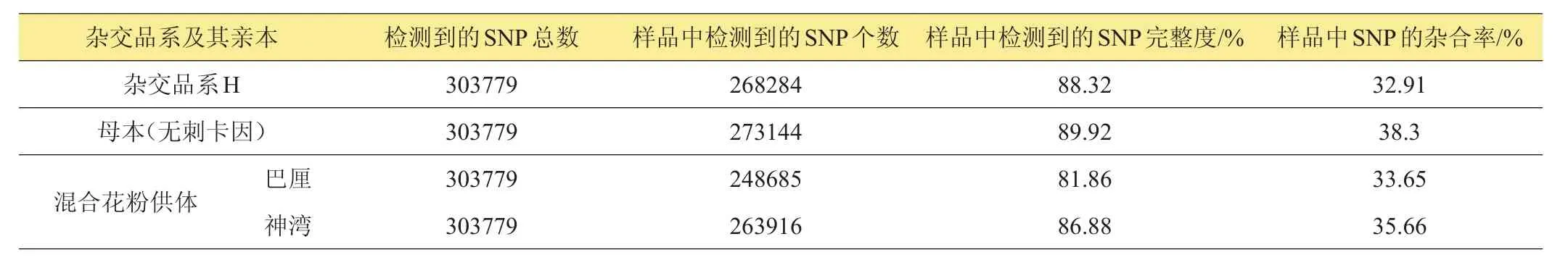
表9 雜交品系及其親本樣品SNP信息統計
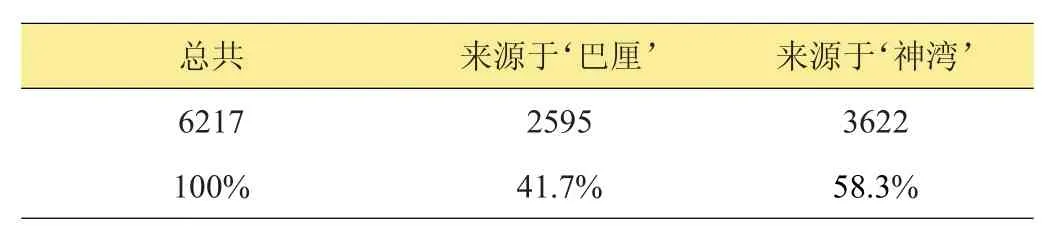
表10 雜交品系‘H’雜合SNP數量及其基因型來源分析
本研究中,‘H’是由‘巴厘’、‘神灣’的混合花粉授到‘無刺卡因’上獲得的雜交品系。‘H’的自然抽蕾、開花期與其母本‘無刺卡因’及混合花粉供體的‘巴厘’、‘神灣’等相差不大;成熟期介于母本‘無刺卡因’及‘巴厘’、‘神灣’之間,屬中早熟種。‘H’果實大小適中,可溶性固形物、蔗糖、全糖含量比品質相對較優的‘神灣’均高,可滴定酸含量低。體現菠蘿果實濃郁芳香的酯類[19]香氣物質含量雖比‘神灣’低,但總體高過‘巴厘’及‘無刺卡因’。
然而,由于混合花粉授粉雜交后代的父本來源未知[2],遺傳背景不明確,很大程度上限制了從遺傳育種到新品種推廣工作的開展[20]。近年來,AFLP、SSR、SRAP等分子標記技術已成功應用于茶樹、棗樹、百合等植物雜種后代父本鑒定[2,20-21],通過測算雜交后代與其親本間的遺傳距離為混合授粉雜交后代的父本確定提供了科學參考[20]。本研究中,根據SLAF測序分析表明雜交品系‘H’的父本來源于‘神灣’的可能性高于‘巴厘’。但測序結果表明‘H’雜合SNP基因型來源于‘神灣’(58.3%)與‘巴厘’(41.7%)的比例相差不大,這可能與2個待驗證父本‘巴厘’、‘神灣’的親緣關系本身就比較接近有關。
表型作為基因的表現型,能一定程度上反映種質間的本質差異,可以快速地分析其遺傳變異,常用于在同一生境下對不同種質材料進行表型性狀評價[22-23]。傳統分類學與統計學及計算技術相結合產生的數量分類學,通過計算雜種后代與雙親之間的遺傳距離,可以再現研究對象之間的親疏關系[5],常用來鑒別真假雜種、雜種優勢選擇以及種質資源聚類分析[24-26]。范凱等[27]基于形態學性狀(葉長、葉寬、一芽三葉長、一芽三葉百芽質量)的遺傳距離分析了茶樹自然雜交后代的遺傳多樣性。王小萍等[10]基于主要生化成分指標(水浸出物、茶多酚、氨基酸、咖啡堿、兒茶素總量、酯型兒茶素總量、簡單兒茶素總量等)對茶樹進行了資源鑒定及遺傳距離分析。本研究中,基于主要物候期、植株大小、果實大小、品質、色澤以及香氣物質組成等6類表型性狀對雜交品系‘H’與其母本‘無刺卡因’以及混合花粉供體‘巴厘’、‘神灣’進行了比較;在2個混合花粉供體‘巴厘’、‘神灣’中,雜交品系‘H’的指標參數整體上與‘神灣’更接近。基于以上6類參數指標的歐氏遺傳距離分析也均顯示雜交品系‘H’與‘神灣’間的遺傳距離小于‘H’與‘巴厘’間的遺傳距離,表明‘神灣’與雜交品系‘H’間的遺傳相似性更高。
通過表型的遺傳相似性分析及SLAF測序分析,可以初步認為混合授粉雜交品系‘H’的父本來源于‘神灣’的可能性更大。
[1]Varela M C,Bras R,Barros I R,et al.Opportunity for hybridization between two oak species in mixed stands as monitored by the timing and intensity of pollen production[J].Forest Ecology and Management,2008,256:1546-1551.
[2]曾貞,羅軍武,楊陽,等.混合花粉授粉茶樹雜交F1代AFLP分析[J].湖南農業大學學報:自然科學版,2008,34(6):719-723.
[3]梁月榮,田中淳一,武田善行.應用RAPD分子標記分析“晚綠”品種的雜交親本[J].茶葉科學,2000,20(1):22-26.
[4]王江民,陳素梅,滕年軍,等.基于形態性狀的菊屬與亞菊屬植物親緣關系的研究[J].植物遺傳資源學報,2013,14(6):1031-1037.
[5]孔志新,王亞,孫明,等.地被菊品種及其近緣種的數量分類[J].東北林業大學學報,2013,41(3):73-77,81.
[6]楊柳慧,于曉南.基于表型性狀的芍藥不同品種群親緣關系分析[J].植物遺傳資源學報,2016,17(2):209-216.
[7]武耀廷,張天真,朱協飛,等.陸地棉遺傳距離與雜種F1、F2產量及雜種優勢的相關分析[J].中國農業科學,2002,35(1):22-28.
[8]倪先林,張濤,蔣開鋒,等.雜交稻特殊配合力與雜種優勢、親本間遺傳距離的相關性[J].遺傳,2009,31(8):849-854.
[9]劉新龍,蔡青,吳才文,等.甘蔗品種資源的表型遺傳多樣性[J].生物多樣性,2010,18(1):37-43.
[10]王小萍,唐曉波,王迎春,等.52份茶樹資源生化組分的表型多樣性分析[J].茶葉科學,2012,32(2):129-134.
[11]蘇文瑾,趙寧,雷劍,等.基于SLAF-seq技術的甘薯SNP位點開發[J].中國農業科學,2016,49(1):27-34.
[12]陳士強,秦樹文,黃澤峰,等.基于SLAF-seq技術開發長穗偃麥草染色體特異分子標記[J].作物學報,2013,39(4):727-734.
[13]彭強,葉生鑫,黃龍,等.運用SLAF-seq技術構建水稻高密度遺傳圖譜[J].分子植物育種,2016,14(8):2127-2132.
[14]仝月澳,周厚基.果樹營養診斷法[M].北京:農業出版社,1982.
[15]Liu C H,Liu Y.Impacts of shading in field on micro-environmental factors around plants and quality of pineapple fruits[J].Journal of Food,Agriculture&Environment,2012,10(2):741-745.
[16]武耀廷,張天真,殷劍美.利用分子標記和形態學性狀檢測的陸地棉栽培品種遺傳多樣性[J].遺傳學報,2001,28(11):1040-1050.
[17]陳香玲,蘇偉強,劉業強,等.36份菠蘿種質的遺傳多樣性SCoT分析[J].西南農業學報,2012,25(2):625-629.
[18]竇美安,張惠云,孫偉生,等.菠蘿無刺卡因和巴厘雜交F1的苗期表型分析[J].熱帶作物學報,2011,32(8):1431-1433.
[19]Kaewtathip T,Charoenrein S.Changes in volatile aroma compounds of pineapple(Ananas comosus)during freezing and thawing[J].International Journal of Food Science and Technology,2012,47:985-990.
[20]馮麗媛,何祥鳳,王文和,等.利用SRAP分子標記技術對混合授粉后的百合F1代父本鑒定[J].沈陽農業大學學報,2015,46(2):230-233.
[21]王斯琪,唐詩哲,孔德倉,等.利用SSR標記進行棗樹子代苗父本鑒定[J].園藝學報,2012,39(11):2133-2141.
[22]劉子記,李靜婷,楊衍,等.辣椒雜交種SSR分子標記鑒定及表型比較分析[J].華北農學報,2014,29(1):69-72.
[23]趙欣欣,張新全,苗佳敏,等.多花黑麥草雜交種SSR分子標記鑒定及表型比較分析[J].農業生物技術學報,2013,21(7):811-819.
[24]Kaushik N,Kumar K,Kumar S,et al.Genetic variability and divergence studies in seed traits and oil content of Jatropha(Jatropha curcasL.)accessions[J].Biomass and Bioenergy,2007,31:497-502.
[25]Klimko M,Boratynska K,Montserrat J M,et al.Morphological variation ofJuniperus oxycedrussubsp.Oxycedrus(Cupressaceae)in the Mediterranean region[J].Flora,2007,202:133-147.
[26]Ganopoulos I,Moysiadis T,Xanthopoulou A,et al.Diversity of morpho-physiological traits in worldwide sweet cherrycultivars of GeneBank collection using multivariate analysis[J].Scientia Horticulturae,2015,197:381-391.
[27]范凱,洪永聰,丁兆堂,等.茶樹‘黃山種’自然雜交后代遺傳多樣性分析[J].園藝學報,2010,37(8):1357-1362.
