植物天冬氨酸蛋白酶的結構與功能
2018-04-10 08:39:40高杉藍興國
生物技術通訊 2018年6期
高杉,藍興國
東北林業(yè)大學 生命科學學院,黑龍江 哈爾濱150040
天冬氨酸蛋白酶(aspartic proteinases,APs)(EC3.4.23)是四大類蛋白水解酶之一,廣泛存在于多種生物中[1]。APs 在MEROPS 數(shù)據(jù)庫(http://merops.sanger.ac.uk/)中被劃分為16 個家族,植物中的APs 主要分布于AA 和AD 族中[2]。在模式植物擬南芥(Arabidopsis thaliana)和水稻(Oryza sati?vaL.)基因組中,目前分別發(fā)現(xiàn)69 和96 個APs 基因[3-4]。近年來,對植物APs 結構與功能的研究表明這些APs 在植物生長發(fā)育、脅迫等方面有重要的生物學功能。
1 植物天冬氨酸蛋白酶的結構與分類
植物APs 主要分為3 種類型:A 類為典型的APs,B 類為類珠心(nucellin-like)特異的APs,C類為非典型的APs。典型的植物APs 主要含有N端的信號肽(signal peptide)、前導片段(proseg?ment)、ASP 結構域(ASP domain)、植物特有插入片段(plant specific insert,PSI)和C 端結構域(car?boxy-terminal)。ASP 結構域包含2 個保守的植物ASP 活性位點,即天冬氨酸-蘇氨酸-甘氨酸(DTG)和天冬氨酸-絲氨酸-甘氨酸(DSG)[3]。PSI是植物APs 特有結構域,包括50~100 個氨基酸殘基[5],在APs 成熟過程中被加工并切除[6]。依據(jù)是否含有PSI,可將植物典型的APs 分為A1 和A2 兩類(表1)。類珠心特異的APs 在蛋白結構上含有N 端的信號肽、ASP 結構域和C 端結構域。與典型植物天冬氨酸蛋白酶相比,類珠心特異的APs不含前導片段和PSI。在擬南芥中,類珠心特異的APs 主要有A41(At1g77480)、A47(At1g44130)、A53(At4g33490)、A46(At1g49050)。非典型的APs被細分為6個亞類(C1、C2、C3、C4、C5 和CX)。C1 亞類含有一個信號肽和一段25~30 個氨基酸殘基的前導片段;C2 亞類具有轉(zhuǎn)運肽特征的富絲氨酸蛋白,但其一個非常罕見的特征是缺乏前導片段,蛋白水解活性必須受pH 或通過內(nèi)源性抑制劑調(diào)節(jié);C3 亞類包含長前導片段;C4 亞類特異性具有C 端延伸區(qū)段;C5 亞類含有DTS 活性位點而不是DTG 活性位點;非典型的APs 序列中不符合上述描述的組被列入CX 亞類(表1)[7]。
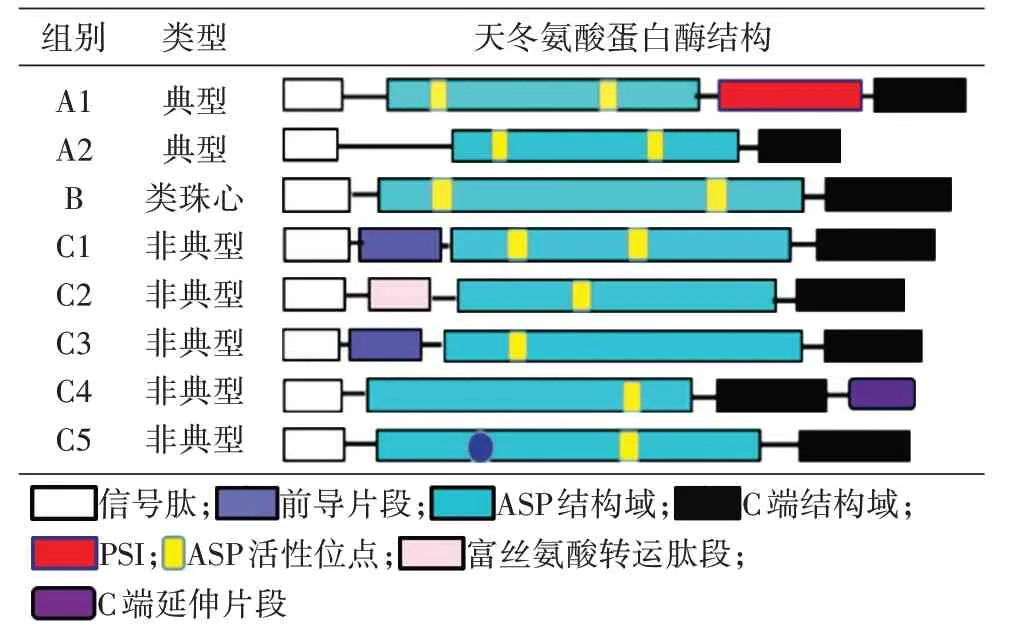
表1 擬南芥天冬氨酸家族基因基本信息
2 植物天冬氨酸蛋白酶的加工機制
目前鑒定的大多數(shù)植物APs 都是用前體結構域合成的,隨后轉(zhuǎn)化為成熟的雙鏈酶[6]。蛋白水解加工在轉(zhuǎn)位至內(nèi)質(zhì)網(wǎng)時切除信號肽、前導片段以及全部或部分去除C 端部分的PSI。前導片段的去除可以通過自動催化過程實現(xiàn),僅須降低pH值,或者可能需要其他分子的催化作用[8]。植物APs 前體的加工涉及去除前導片段和PSI 結構域,然而每個肽段從前體中除去的機理和順序不盡相同。第一類是PSI 在前導片段之前被完全移除。刺苞菜薊(Cynara cardunculusL.)中cardosin A 的前體procardosin A 是相對分子質(zhì)量為64 000的含有PSI 的前體,在低pH 值條件下PSI 被切割并且可被胃蛋白酶抑制劑A 抑制[9]。第二類是先去除前導片段,然后再切割PSI。大麥天冬氨酸蛋白酶最先進行N 端前導片段的加工,然后再切割PSI[10]。由于體外獲得的中間形式和最終產(chǎn)物與體內(nèi)檢測的產(chǎn)物略有不同,可能蛋白質(zhì)的完全成熟還需要其他蛋白酶的參與[11]。第三類是切除了前導片段和部分PSI。在畢赤酵母中表達重組成熟的cyprosin,首先切除前導片段和大部分PSI,然后通過二硫鍵把輕鏈和重鏈結合在一起[12]。此外,畢赤酵母中重組progaline B 的活化須首先部分去除存在于植物APs 中的PSI,將前體蛋白切割成2 條多肽鏈,隨后去除前導片段[13]。
3 植物天冬氨酸蛋白酶的功能
通過對植物APs 遺傳突變體的表型分析,結合對APs 的生理生化方面的研究,已發(fā)現(xiàn)植物APs 在植物生物脅迫、非生物脅迫、種子萌發(fā)、有性生殖及植物衰老等方面具有重要作用。
3.1 生物脅迫
植物在生長發(fā)育過程中會受到病原菌的生物脅迫,APs 在這過程中發(fā)揮著重要作用。編碼擬南芥天冬氨酸蛋白酶A62(AT1g92290)的組成型抗病性基因CDR1,其過表達導致矮化病和對毒性丁香假單胞菌的抗性;反義CDR1植物對病原體的抵抗力減弱,比野生型更易受病原菌株的影響,由此證明該基因可能與病原菌種的抵抗相關[14]。編碼擬南芥A46(At1g49050)的APCB1參與BAG6介導的基礎抗性,靶向失活天冬氨酸蛋白酶APCB1基因?qū)е驴共⌒詥适15]。編碼水稻天冬氨酸蛋白酶的基因OsCDR1(OsAP5)在過表達轉(zhuǎn)基因植物中表現(xiàn)為相關防御基因增加與細菌真菌病原體抗性的增強,證明其參與水楊酸介導的抗病性[16]。用稻瘟病真菌(Magnaporthe oryzae)、葉枯病菌(Xanthomonas or orzazae pv.oryzae)或黃瓜花葉病毒(CMV)感染,水稻維管組織中的OsAP77表達明顯增強,認為水稻OsAP77基因參與了生物脅迫機制[17]。此外,在馬鈴薯中也鑒定出響應生物脅迫的APs。馬鈴薯天冬氨酸蛋白酶StAsp 在致病疫霉感染后的葉片中累積,StAsp 表達水平與馬鈴薯栽培種對致病疫霉的抗性程度正相關[18]。把馬鈴薯天冬氨酸蛋白酶StAP-PSI 轉(zhuǎn)化擬南芥植株,顯著增強擬南芥對灰葡萄孢菌感染的抗性。StAP-PSI 被分泌到葉質(zhì)外體中并直接作用于病原體,從而補充植物先天性免疫反應[19]。
3.2 非生物脅迫
編碼擬南芥天冬氨酸蛋白酶A5(At3g18490)的ASPG1(Aspartic Protease In Guard Cell 1)參與非生物脅迫反應。過度表達ASPG1導致保衛(wèi)細胞中脫落酸(ABA)敏感性增加,減少植物中水的損失,可以賦予擬南芥抗旱性[20]。用水楊酸(SA)、異煙酸(INA)、過氧化氫(H2O2)和ABA 處理水稻,OsAP77在維管組織中的表達增加[17]。普通大豆(Phaseolus vulgaris)葉片中天冬氨酸蛋白酶前體PvAP1 的表達受水分脅迫的嚴格調(diào)控[21]。耐寒菠蘿(Ananas comosus)中的天冬氨酸蛋白酶AcAP1,在采摘后冷卻處理條件下表現(xiàn)出前體基因表達的上調(diào),而易感性表現(xiàn)出采摘后冷卻處理的下調(diào),表明AcAP1 與收割后低溫脅迫下抗黑心病的發(fā)展呈正相關[22]。這些研究結果說明植物APs 參與了植物的非生物脅迫反應。
3.3 種子萌發(fā)
擬南芥2S 白蛋白(albumin)是一種液泡蛋白,經(jīng)翻譯后加工形成成熟的2S 白蛋白,植物APs 在此過程中發(fā)揮重要作用[23]。蓖麻種子中的APs 在體外能夠?qū)?S 白蛋白的前肽(pro2S)進行部分加工。亞細胞定位表明,天冬氨酸內(nèi)肽酶定位于蓖麻籽成熟胚乳液泡的基質(zhì)區(qū)域,因此認為蓖麻種子的APs 可能參與了pro2S 白蛋白前體的成熟過程[24]。此外,擬南芥的ASPG1參與種子的萌發(fā),突變ASPG1導致種子休眠且種子活力降低,當在無營養(yǎng)培養(yǎng)基上生長時,突變體aspg1-1的幼苗停止發(fā)育。因此,ASPG1對種子休眠、種子壽命和種子萌發(fā)很重要[25]。水稻OsAp41編碼的Oryzasin1在萌發(fā)的種子、幼嫩的根和葉中表達量較高,水稻開花后2~4 周,Oryzasin1的表達呈上升趨勢,種子形成后其表達量下降,說明Oryzasin1在種子形成過程中發(fā)揮重要作用,推測其通過降解儲備蛋白以供萌發(fā)需要[26]。另外,小麥中的天冬氨酸蛋白酶WAP1和WAP2 可以在體外消化貯藏蛋白谷蛋白,推測其可能參與種子萌發(fā)過程[27]。
3.4 有性生殖
天冬氨酸蛋白酶在絨氈層和配子體發(fā)育中起重要作用。沉默編碼擬南芥A51(At4g12920)的AtUNDEAD的表達,導致絨氈層PCD 出現(xiàn)較早,花粉敗育,說明植物APs 對于絨氈層和花粉發(fā)育是必需的[28]。水稻OsAP25和OsAP37也參與絨氈層的降解,過表達OsAP25和OsAP37誘導植物和酵母細胞死亡,同時參與轉(zhuǎn)錄因子正調(diào)節(jié)水稻花藥中絨氈層細胞的程序性細胞死亡,揭示了APs在水稻生殖發(fā)育動態(tài)調(diào)節(jié)中的重要作用[29]。編碼擬南芥天冬氨酸蛋白酶A35(At5g02190)的PCS1與胚胎發(fā)育和有性生殖有關,PCS1功能喪失突變導致雄性和雌性配子體的變性及發(fā)育中胚胎細胞的過度死亡;相反,PCS1過表達通常導致花藥開裂處和隔膜處細胞存活,表明PCS1對胚胎發(fā)育是必需的[30]。Gao 等[31]描述了新的擬南芥APs A36(At5g36260)和A39(Atlg65240),二者是膜錨定蛋白,與GPI 錨定蛋白COBRA-Like10 共定位于花粉管細胞壁的頂端區(qū)域。a36a39突變體顯示明顯的敗育,花粉粒發(fā)生類似凋亡的程序性細胞死亡,表明A36 和A39 是參與配子體發(fā)生和擬南芥花粉導向的重要因素。水稻OsAP65 蛋白定位于細胞內(nèi)的囊泡中,其編碼基因的缺失導致花粉萌發(fā)和花粉管的生長受到抑制,最終導致雄配子體的敗育[32]。大麥天冬氨酸蛋白酶基因nucellin則是在發(fā)生雙受精后進行表達。授粉前nucellin基因僅在靠近合胞體端的胚囊附近的珠心細胞簇中以非常低的水平表達,但授粉后其在整個胚胎周圍的大部分珠心細胞中高度表達。nucellin基因表達的時間和空間模式與珠心細胞同步,推測nucellin與珠心細胞死亡有關[33]。
3.5 植物衰老
隨著植物莖、葉、種子等器官的衰老,表現(xiàn)為生長速率的下降,如光合速率下降、核酸和蛋白質(zhì)含量下降、線粒體體積減小導致呼吸速率下降、激素水平變化等。在植物衰老過程中,APs 主要參與光合作用關鍵酶的降解、激素調(diào)控及細胞器降解等過程。煙草CND41 是一種在葉綠體中表達的與DNA 結合的天冬氨酸蛋白酶,它通過降解二磷酸核酮糖羧化酶(Rubisco),在葉片老化中發(fā)揮重要作用[34]。馬鈴薯SPAP1 是一種功能性典型的天冬氨酸蛋白酶,參與乙烯利介導葉片的衰老。SPAP1 促進的葉片衰老及其活性很可能與PSI 結構域無關[35]。天冬氨酸蛋白酶在白楊(Pop?ulus tremula)的幼葉中表達很少,但在衰老葉片中的表達量明顯升高,可能與細胞中葉綠體的降解緊密相關[36]。在豇豆(Vigna unguiculataL.Walp.)中通過基因組文庫篩選分離天冬氨酸蛋白酶基因,發(fā)現(xiàn)豇豆APs 在葉和莖中積累但在根中不積累,在衰老的葉片中轉(zhuǎn)錄物累積增加,表明該酶可能參與葉片衰老過程[37]。
3.6 其他功能
豬籠草(Nepenthes macferlaneiL.)產(chǎn)生各種水解酶,捕食昆蟲以獲得蛋白質(zhì),這些水解酶中包括天冬氨酸蛋白酶[38]。另外,稻根霉菌(Rhizopus oryzae)產(chǎn)生的天冬氨酸蛋白酶Rhio1 能觸發(fā)各種過敏性疾病,說明Rhio1 具有免疫學特征,將有助于診斷霉菌過敏[39]。
4 結語
目前雖然對植物APs 的結構和功能有了一定的認識,但大部分APs 的作用機制尚不清楚。在擬南芥和水稻中,大部分APs 的生物學功能尚未得到研究,而且對其下游底物及信號傳遞因子知之甚少。未來,需要通過大規(guī)模系統(tǒng)生物學方法結合遺傳學方法(如CRISPR/Cas9),進一步解析APs 的生物學功能。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
