銅綠假單胞菌重組Bb-OprI疫苗免疫及PA01株感染后小鼠脾細胞因子IL-17和Treg的變化
2018-04-09 03:37:20劉瀟李文桂
生物技術通訊 2018年2期
關鍵詞:小鼠
劉瀟,李文桂
重慶醫科大學 附屬第一醫院 傳染病寄生蟲病研究所,重慶 400016
銅綠假單胞菌(Pseudomonas aeruginosa,Pa)是院內感染的主要病原體之一。Pa易定植,且耐藥機制多樣復雜,可導致免疫力受損或囊性纖維化(cystic fibrosis,CF)等患者發生難以清除的感染[1]。疫苗具有良好的特異性,且受耐藥機制的影響小,為防治Pa感染提供了新的思路。
Pa的外膜蛋白I(outer membrane protein I,OprI)是一種具有較高保守性的外膜蛋白,既往研究[2-3]證實OprI具有良好的免疫原性,是Pa疫苗研制有希望的候選分子之一。兩歧雙歧桿菌(Bifi?dobacterium bifidum,Bb)不僅是人與動物腸道內重要的益生菌之一,其穩定表達外源抗原的特征使其成為新型疫苗載體的研究熱點[4]。
本課題組前期研究發現重組Bb-OprI疫苗可誘導免疫小鼠CD4+T細胞比例升高[5]。Th17細胞是重要的CD4+T細胞之一,既往研究[6-7]表明Th17細胞及其分泌的IL-17在宿主對Pa感染的免疫防御中有重要作用。CD4+CD25+Foxp3+T細胞是機體重要的調節性T細胞(regulatory T cell,Treg),與Th17細胞相互拮抗[8],在維持機體自身的免疫穩定中發揮不可或缺的作用[9-10]。我們擬將重組Bb-OprI疫苗接種BALB/c鼠,用PA01株攻擊后研究脾細胞IL-17和Treg的變化,探索其免疫機制及對Pa感染的保護作用。
1 材料與方法
1.1 材料
21只SPF級雌性BALB/c小鼠,購自重慶醫科大學實驗動物中心;重組Bb-OprI疫苗由本所構建并保存;銅綠假單胞菌PA01株由重慶醫科大學附屬兒童醫院余加林教授惠贈。
DNA抽提試劑盒、PCR試劑盒、RPMI1640培養基、胎牛血清(FBS)、青霉素、鏈霉素購自上海生工公司;LB液體培養基、LB固體培養基、MRS液體培養基和銅綠假單胞菌抗原(Pa antigen,PaAg)由本所保存;PCR儀、電泳儀、凝膠成像分析儀購自Bio-Rad公司;離心機購自Eppendorf公司;CO2培養箱購自Thermo Forma公司。
1.2 動物免疫及攻擊方案
BALB/c鼠隨機分為Bb-OprI疫苗組、空載體(Bb-pGEX-1λT)對照組和Bb對照組,每組7只,用100 μL MRS培養基重懸5×109CFU的疫苗進行灌胃接種,每周連續接種3 d,連續接種3周。初次接種后4周用10 μL LB液體培養基重懸5× 106CFU的PA01株進行滴鼻攻擊。
1.3 肺組織菌落計數
PA01株攻擊后2周處死小鼠,分離肺組織,勻漿稀釋后涂布于LB固體培養基,37℃培養48 h,計數菌落數。
1.4 脾細胞制備
分離小鼠脾臟,常規制備脾細胞懸液,用含10%FBS、1×105U/L青霉素和1×105μg/L鏈霉素的RPMI1640培養基使脾細胞重懸,調整細胞終濃度為5×106/mL。
1.5 脾細胞誘導和培養
細胞培養板中加入上述制備的1 mL脾細胞懸液和10 μg PaAg,37℃、5%CO2條件下培養48 h,4000 r/min離心5 min收集脾細胞。
1.6 PCR擴增脾細胞IL-17和Foxp3基因
用DNA提取試劑盒提取脾細胞DNA,以此為模板擴增IL-17和Foxp3基因,引物由上海生工公司合成(表1)。反應體系均為50 μL,包括25 μL 2×PCR Master,2 μL上、下游引物,2 μL DNA模板,19 μL滅菌雙蒸水。反應條件:①IL-17:95℃ 1 min;95℃ 5 s,60℃ 5 s,72℃ 30 s,46個循環;72℃ 10 min;②Foxp3:95℃ 5 min;95℃ 10 s,55℃ 30 s,72℃ 30 s,40個循環;72℃ 10 min。擴增產物進行1.2%瓊脂糖凝膠電泳鑒定。
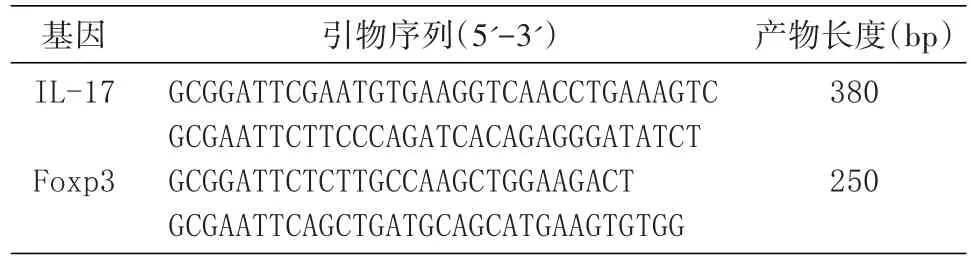
表1 PCR引物
1.7 統計學分析
用SPSS20.0軟件對實驗數據進行分析。實驗數據以x±s表示,多組間比較用Kruskal-Wallis檢驗,P<0.05為差異有統計學意義。
2 結果
2.1 肺組織的菌落計數
Bb-OprI疫苗組、空載體對照組和Bb對照組小鼠肺組織的菌落數分別為(0.46±0.09)×108、(5.42±0.79)×108和(6.20±0.95)×108CFU,Bb-OprI疫苗組明顯低于空載體對照組(P<0.01)和Bb對照組(P<0.01),2個對照組之間差異無統計學意義(P>0.05)(圖1)。
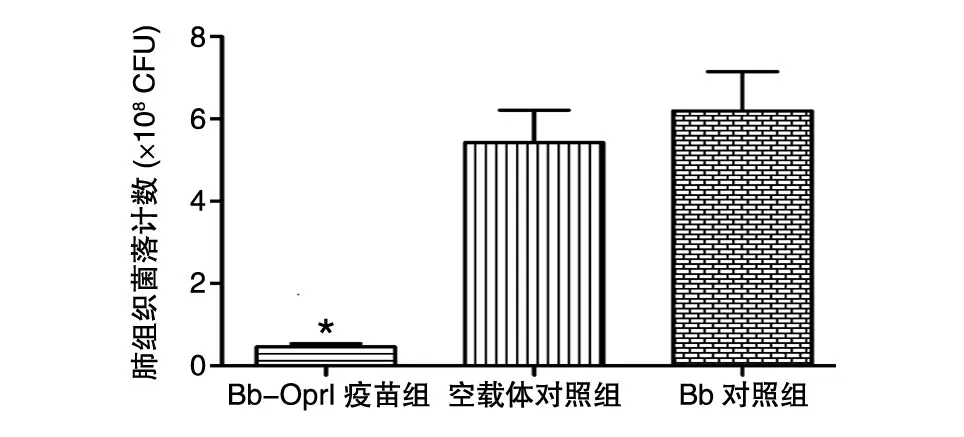
圖1 疫苗免疫及PA01株攻擊后小鼠肺組織的菌落計數*:與空載體和Bb對照組相比,P<0.01
2.2 IL-17和Treg檢測
PCR檢測表明,疫苗組可擴增出380 bp的IL-17基因及250 bp的Foxp3基因(圖2、3)。空載體對照組和Bb對照組未能擴增出IL-17和Foxp3編碼基因。
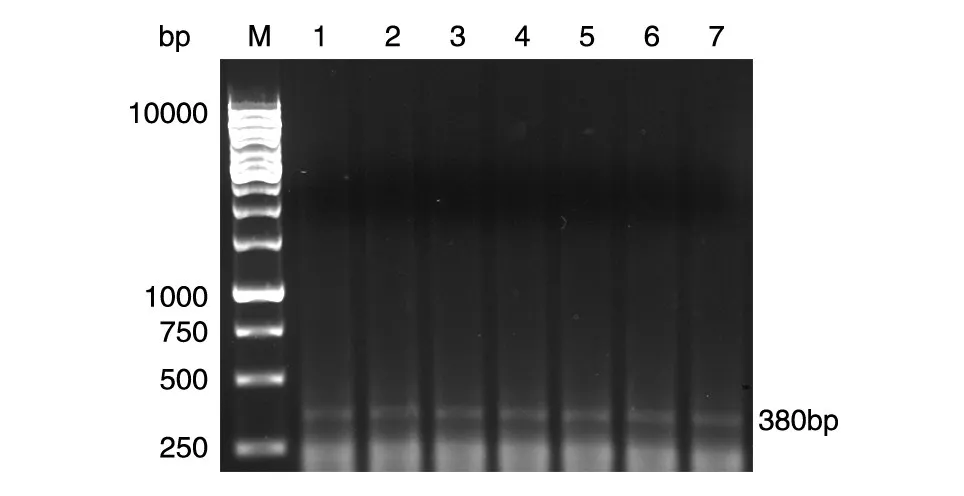
圖2 疫苗免疫及PA01株攻擊后小鼠脾細胞IL-17基因的PCR擴增結果M:DNA marker;1~7:IL-17基因的PCR產物
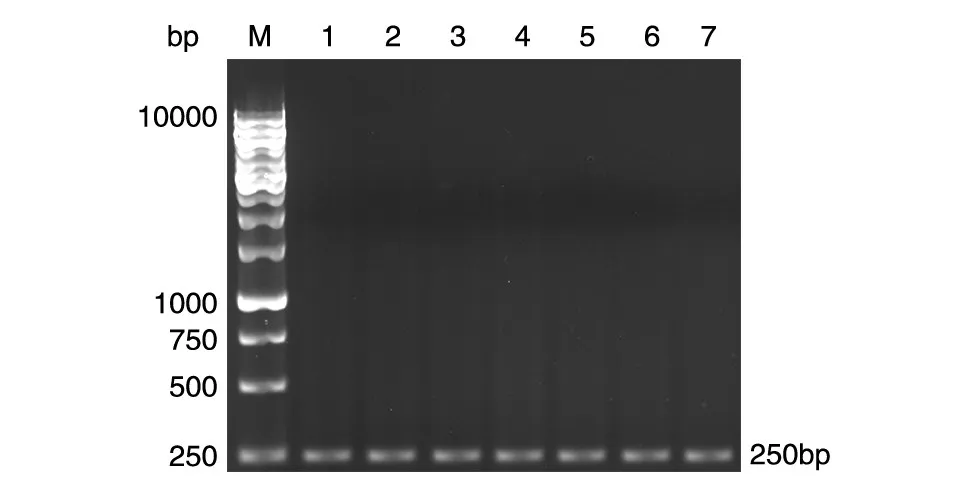
圖3 疫苗免疫及PA01株攻擊后小鼠脾細胞Foxp3基因的PCR擴增結果M:DNA marker;1~7:Foxp3基因的PCR產物
3 討論
Yang等[2]將OprI蛋白加Al(OH)3佐劑肌肉注射免疫BALB/c鼠,發現PA01株攻擊后免疫鼠的肺組織Pa負荷量明顯減少。本研究將重組Bb-OprI疫苗灌胃接種BALB/c鼠,發現PA01株攻擊后免疫鼠肺組織的細菌菌落數顯著減少,與上述研究結果相似,表明該疫苗可誘導小鼠產生一定的保護力。
Liu等[7]發現急性Pa感染后,Th17細胞產生IL-17迅速增多,用IL-17抗體中和IL-17后,小鼠支氣管肺泡灌洗液(bronchoalveolar lavage fluid,BALF)中的細菌數明顯增多,肺組織的病理損傷明顯加重,粒細胞集落刺激因子(granulocyte colo?ny-stimulating factor,G-CSF)和角質形成細胞趨化物(keratinocyte chemoattractant,KC)水平下降,使肺組織募集的中性粒細胞顯著減少。Xu等[11]發現感染Pa后,小鼠BALF中IL-17水平和中性粒細胞計數顯著升高;在正常小鼠氣管內滴入表達IL-17的pcDNA3.1-IL-17質粒,可使BALF中促炎癥因子IL-1β、巨噬細胞炎癥蛋白2(macro?phage inflammatory protein-2,MIP-2)和G-CSF及中性粒細胞升高,PA01株攻擊后小鼠肺組織細菌計數明顯減少,生存率顯著升高。這些研究結果提示IL-17可能通過下游因子的作用,募集中性粒細胞等免疫細胞發揮抵御Pa感染的保護作用。Chen等[12]在肺炎克雷伯菌肺部感染小鼠的模型中證實,在多個血清型間保守的外膜蛋白免疫小鼠可使縱膈淋巴結的記憶性Th17細胞活化增殖,從而提供長期的保護力。Aguado-Martínez等[13]將OprI與犬新孢子蟲的嵌合蛋白Mic3-1-R融合并免疫小鼠,發現融合蛋白誘導脾細胞產生的IL-17水平明顯高于單個Mic3-1-R蛋白。Yang等[2]將OprI外膜蛋白加Al(OH)3佐劑肌肉注射免疫BALB/c鼠,末次免疫后2周脾細胞IL-17升高,保護力實驗顯示該蛋白疫苗可以提高2倍LD50的PA01株攻擊后小鼠的生存率。這些研究結果提示OprI蛋白作為疫苗分子或佐劑,均具有較好的誘導Th17免疫應答的能力,從而抵抗Pa的感染。本研究中我們用重組Bb-OprI疫苗灌胃免疫BALB/c鼠,PA01株攻擊后免疫鼠脾細胞IL-17升高,與上述研究[2]相似,提示Bb-OprI疫苗可誘導小鼠產生Th17免疫應答。推測抗原提呈細胞將OprI抗原呈遞給脾CD4+T細胞,在轉化生長因子β(transforming growth factor-β,TGF-β)和IL-6的誘導下,CD4+T細胞向Th17細胞分化,分泌IL-17并形成記憶性Th17細胞,在PA01株攻擊后,菌體的OprI蛋白刺激記憶性Th17細胞迅速大量增殖,分泌更高水平的IL-17,刺激肺泡上皮細胞分泌單核細胞趨化蛋白-1(monocyte chemotac?tic protein-1,MCP-1)、MIP-1α/β和趨化因子CX?CL13等,募集并活化巨噬細胞[14-15],通過氧依賴途徑等殺傷Pa;分泌CXCL8和G-CSF等,募集并激活中性粒細胞吞噬和清除Pa[16]。
疫苗免疫產生適當的免疫應答可為機體提供良好的保護力,但過度的免疫應答則有可能造成機體損傷。Mulcahy等[17]發現CF患者外周血中Th17細胞的比例越高,肺功能越差。丁鳳鳴等[18]證實Pa慢性感染后小鼠Th17細胞功能亢進而導致的IL-17水平持續升高,可使小鼠肺組織持續被大量的以中性粒細胞為主的白細胞浸潤,中性粒細胞釋放氧化劑和蛋白酶等成分造成肺組織嚴重的免疫性病理損傷[19]。因此,由Treg介導的免疫調節作用至關重要。Sanchez等[9-10]證實在病原體初次感染時,Treg通過TCR被特異性抗原激活,并且不顯著影響CD4+T細胞的增殖和分化,初次免疫應答后激活的Treg可轉化為記憶性Treg,再次免疫應答時,記憶性Treg以比初次免疫更快的速度和更多的數量增殖并募集至感染部位,通過分泌大量IL-10抑制CD4+T細胞的大量增殖,從而在不影響病原體清除的情況下抑制過強的再次免疫應答產生的組織損傷。Hector等[20]發現CF患者的外周血Treg的數量與肺功能正相關,慢性Pa感染的CF患者Treg數量明顯降低,記憶性Treg的數量也顯著下降,Treg細胞對非Treg細胞的抑制功能下降,提示Treg細胞在保護Pa感染患者的肺功能及改善預后方面有重要作用。本研究發現重組Bb-OprI疫苗免疫及PA01株攻擊后,免疫鼠脾細胞中Treg增多,推測Bb-OprI疫苗免疫后,能夠誘導記憶性Treg生成,PA01株攻擊后Treg增殖活躍,活化的Treg分泌TGF-β和IL-10等細胞因子[21],作用于Th17細胞,抑制其過度增殖與活化,最終表現為在保證有足夠的Th17細胞參與免疫應答并清除Pa感染的前提下,防止過多的IL-17所誘發的炎癥反應和免疫損傷,保持內環境穩定的免疫狀態。
本研究選用的疫苗分子OprI蛋白是一種無毒性、無致熱原性,并且在Pa的多個血清型間具有保守性的疫苗分子[22];本研究所用疫苗載體為Bb,使該疫苗具有安全無毒和接種方便等優點,重組Bb-OprI疫苗的研制有望為抗Pa感染治療提供新的思路。后續將進一步摸索疫苗免疫的劑量和途徑,研究疫苗的保護性免疫機制等。
[1]Ullah W,Qasim M,Rahman H,et al.Beta-lactamaseproducing Pseudomonas aeruginosa:Phenotypic charac?teristics and molecular identification of virulence genes [J].J Chin Med Assoc,2017,80(3):173-177.
[2] Yang F,Gu J,Yang L,et al.Protective efficacy of the trivalentPseudomonasaeruginosa vaccine candi?date PcrV-OprI-Hcp1 in murine pneumonia and burn models[J/OL].Sci Rep,2017,7(1):3957.
[3] Hassan R,El-Naggar W,Abd El-Aziz A M,et al. Immunization with outer membrane proteins(OprF and OprI)and flagellin B protects mice from pulmonary in?fection with mucoid and nonmucoid Pseudomonas aeru?ginosa[J].J Microbiol Immunol Infect, 2017. doi: 10.1016/j.jmii.2016.08.014.
[4] 譚建蓉,李文桂,覃婷.日本血吸蟲重組Bb(pGEXSj32)疫苗免疫BALB/c小鼠誘導免疫應答的動態研究[J].中國人獸共患病學報,2015,31(9):805-811.
[5] 劉瀟,李文桂.兩歧雙歧桿菌介導的銅綠假單胞菌外膜蛋白I(Bb-OprI)疫苗免疫增強小鼠對銅綠假單胞菌的抑制作用[J].細胞與分子免疫學雜志,2017,33(8):1040-1044.
[6] Bayes H K,Ritchie N D,Evans T J.Interleukin-17 is required for control of chronic lung infection caused by Pseudomonas aeruginosa[J].Infect Immun, 2016,84(12):3507-3516.
[7] Liu J,Feng Y,Yang K,et al.Early production of IL-17 protects against acute pulmonary Pseudomonas aeru?ginosa infection in mice[J].FEMS Immunol Med Mi?crobiol,2011,61(2):179-188.
[8] Noack M,Miossec P.Th17 and regulatory T cell bal?ance in autoimmune and inflammatory diseases[J].Au?toimmun Rev,2014,13(6):668-677.
[9] Sanchez A M,Zhu J,Huang X,et al.The develop?ment and function of memory regulatory T cells after acute viral infections[J].J Immunol,2012,189(6):2805-2814.
[10]Brincks E L,Roberts A D,Cookenham T,et al.Anti?gen-specific memory regulatory CD4+Foxp3+T cells control memory responses to influenza virus infection [J].J Immunol,2013,190(7):3438-3446.
[11]Xu X,Shao B,Wang R,et al.Role of Interleukin-17 in defense againstpseudomonas aeruginosa infection in lungs[J].Int J Clin Exp Med,2014,7(4):809-816.
[12]Chen K,McAleer J P,Lin Y,et al.Th17 cells medi?ate clade-specific,serotype-independent mucosal im?munity[J].Immunity,2011,35(6):997-1009.
[13]Aguado-Martínez A,Basto A P,Müller J,et al.N-ter?minal fusion of a toll-like receptor2-ligand to a Neosporacaninum chimeric antigen efficiently modi?fies the properties of the specific immune response[J]. Parasitology,2016,143(5):606-616.
[14]Gopal R,Rangel-Moreno J,Slight S,et al.Interleu?kin-17-dependent CXCL13 mediates mucosal vaccineinduced immunity against tuberculosis[J].Mucosal Im?munol,2013,6(5):972-984.
[15]Liu J,Qu H,Li Q,et al.The responses of γδ T-cells against acute Pseudomonas aeruginosa pulmonary infection in miceviainterleukin-17[J].PathogDis, 2013,68(2):44-51.
[16]Pelletier M,Maggi L,Micheletti A,et al.Evidence for a cross-talk between human neutrophils and Th17 cells[J].Blood,2010,115(2):335-343.
[17]Mulcahy E M,Hudson J B,Beggs S A,et al.High peripheralblood Th17 percentassociated with poor lung function in cystic fibrosis[J/OL].PLoS One,2015, 10(3):e0120912.
[18]丁鳳鳴,朱松雷,沈策.Th1和Th17細胞功能失調在銅綠假單胞菌慢性肺部感染中的致病作用[J].免疫學雜志,2012,28(6):510-514.
[19]Kruger P,Saffarzadeh M,Weber A N,et al.Neutro?phils:Between host defence,immunemodulation,and tissue injury[J/OL].PLoS Pathog,2015,11(3):e1004651.
[20]Hector A,Sch?fer H,P?schel S,et al.Regulatory T-cell impairment in cystic fibrosis patients with chron?ic Pseudomonas infection[J].Am J Respir Crit Care Med,2015,191(8):914-923.
[21]Stewart C A,Metheny H,Iida N,et al.Interferon-de?pendent IL-10 production by Tregs limits tumor Th17 inflammation[J].J Clin Invest,2013,123(11):4859-4874.
[22]Basto A P,Morais J,Marcelino E,et al.An efficient depyrogenation method for recombinant bacterial outer membrane lipoproteins[J].Protein Expr Purif,2014,98: 10-17.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
