基于DNA條形碼技術的永暑礁瀉湖魚卵鑒定研究
2018-03-21 01:07:34郭華陽朱克誠楊靜文江世貴張殿昌
淡水漁業(yè) 2018年2期
吳 娜,張 楠,曹 明,郭華陽,朱克誠,楊靜文,江世貴,張殿昌
(1.中國水產科學研究院南海水產研究所,農業(yè)部南海漁業(yè)資源開發(fā)利用重點實驗室,廣東省漁業(yè)生態(tài)環(huán)境重點開放實驗室,廣州 510300;2.上海海洋大學水產與生命學院,上海 201306;3.廣東省漁業(yè)種質保護中心,廣州 510300)
魚卵和仔稚魚是魚類生活史的早期階段,魚卵和仔稚魚調查是研究魚類繁殖習性、生殖洄游路線、產卵場、種類分布和種群動態(tài)的基礎,也可為魚類人工繁育技術和生理生化機制研究提供基礎數據[1]。魚卵和仔稚魚種類的準確鑒別,是開展魚卵和仔稚魚研究的重要前提。通常,魚卵鑒別主要依據卵的形狀、大小、卵周隙、油球、內膜、絨毛膜、卵黃形態(tài)和胚胎形狀等形態(tài)特征[2],然而近緣物種的魚卵由于形態(tài)相似很難區(qū)分,大多種類只能鑒定到科或屬的水平,目前只有極少種類魚卵鑒定到種的水平,尋求穩(wěn)定、準確的分類依據和鑒定方法,是魚類浮游生物研究亟待解決的關鍵問題。DNA條形碼(DNA barcoding)技術是一種利用標準基因區(qū)域序列多態(tài)性進行物種鑒定的分子方法,條形碼序列分析不依賴于形態(tài)特征,對形態(tài)特征近似,形態(tài)學分類比較困難的疑難種類的分類以及形態(tài)學分類上存有疑問種類的驗證提供了新的方法和手段[3-4]。魚類線粒體細胞色素C氧化酶亞基I(mtCOI)基因具有序列變異水平適中,變異區(qū)域兩端序列高度保守和序列長度適宜的特點,是被廣泛接受的魚類DNA條形碼標準基因[5-7],目前魚類DNA條形碼數據庫(Barcode of Life Data System [BOLD])中已有超過17.5萬件憑證標本,代表了1.5萬種以上的魚類種類[8]。隨著魚類DNA條形碼數據庫的不斷完善,研究證實DNA條形碼技術在魚卵和仔稚魚的種類鑒定中也具有重要的應用價值[9-11]。
南沙群島位于南海南部,屬于印度洋-太平洋熱帶動物區(qū)系,是世界上海洋生物最豐富的區(qū)域之一。根據已有的調查資料顯示南海魚類有3 000多種,分別隸屬于3個綱、35個目、236個科和822屬[12]。目前南海魚類中只對極少數種類的魚卵和仔稚魚進行過研究[4,13-14],絕大多數種類的魚卵和仔稚魚的形態(tài)特征并不清楚,使得南海重要經濟魚類產卵行為、產卵場判別和種群補充機制研究無法開展。永暑礁是一座珊瑚環(huán)礁,為珊瑚礁魚類和熱帶大洋性魚類的重要產卵場,然而目前未見有關永暑礁海域魚卵鑒定和分類的相關研究。因此,本研究擬以永暑礁瀉湖魚卵為研究對象,采用形態(tài)分類特征和DNA條形碼技術對采集的魚類進行鑒定分析,以期了解珊瑚礁魚類和熱帶大洋性魚類的產卵季節(jié)和種類分布特征,為相關種類的種群補充機制研究提供基礎材料。
1 材料和方法
1.1 樣品采集和形態(tài)特征分析
依托中國水產科學研究院南海水產研究所“南鋒號”科學調查船,于2016年4月13號在中國南海永暑礁瀉湖海域按照《海洋調查規(guī)范》(GB12763.1-7-91)進行魚卵樣品采集。采用孔徑為0.505 mm篩絹制成的大型浮游生物網(網口內徑80 cm、長270 cm、網口面積為0.5 m2)按照設定站位進行表層水平和垂直拖網,采集魚卵樣品,-20 ℃冰箱中保存?zhèn)溆谩2杉臉悠愤\至實驗室后,于解剖鏡下進行形態(tài)特征觀察和顯微拍照,觀察記錄魚卵的卵膜(Chorion)、圍囊腔(Perivitelline space)、卵黃(Yolk)、油球(Oil globule)、色素分布(Pigmentation)和胚體發(fā)育(Embryo)等形態(tài)特征和魚卵卵徑參數,并對魚卵形態(tài)特征進行了描述分析(表1)。
1.2 DNA提取和COI基因測序分析
魚卵樣品用雙蒸水沖洗,將樣品置于1.5 mL離心管中,采用Magen公司的試劑盒(HiPure Mollusc DNA Kit)提取基因組DNA,提取的DNA置于-20 ℃保存?zhèn)溆谩R镌O計參照Ward等[15],由廣州睿博生物公司合成,序列擴增引物為Fish-F:5′-TCRACYAAYCAYAAAGAYATYGGCAC-3′和Fish-R:5′-ACTTCWGGGTGRCCRAAGAATCA-3′。PCR擴增總體積為25 μL,反應體系包括RNase-Free Water (16.7 μL),10×loading buffer (Mg2+free)2.5 μL,DNTP(2.5 mmol/L)2 μL,MgCl2(2.5 μmol/L)1.5 μL,F(xiàn)ish-F(10 μmol/L)0.5 μL,F(xiàn)ish-R(10 μmol/L)0.5 μL,rTaq (5 U/μL)0.3 μL及DNA模板(20 ng/μg)1.0 μL。PCR反應程序為:94 ℃預變性3 min,94 ℃變性30 s,55 ℃退火30 s,72 ℃延伸90 s,共35個循環(huán),最后72 ℃延伸10 min。PCR產物進行1.2%的瓊脂糖膠電泳檢測,選取擴增結果較好的產物送廣州睿博興科股份有限公司測序分析。
由于魚卵樣品在提取DNA、PCR擴增及測序的實驗過程中,都有可能會出現(xiàn)結果不佳的情況,因此本研究以最終獲得的有效的序列個體來進行分析。
1.3 分子鑒定分析
測序獲得的魚卵COI基因序列與BOLD系統(tǒng)數據庫(http://www.barcodinglife.org)比對分析進行物種鑒定,在比對分析中,BOLD系統(tǒng)為每個樣品分配一個條形碼索引號(Barcode Index Number,BIN),當樣品COI基因序列與BIN中參考序列的相似性≥98.4%時,鑒定為種,當相似性在95%~ 98.3%之間,鑒定到屬,當相似性<95%時,為未知種[2,16]。利用MEGA6.0軟件根據配對遺傳距離采用Kimura two-parameter model (K2P)構建鄰接進化樹(Neighbor-joining tree)[17],進化樹末端節(jié)點代表了種類分組。
2 結果和分析
2.1 魚卵種類鑒定分析
采集的71個魚卵樣品根據外部形態(tài)特征初步劃分為圓形和橢圓形2種類型,圓形魚卵44個,橢圓形魚卵27個。經DNA提取和線粒體COI基因片段測序分析,71個魚卵樣品中的30個個體獲得了有效的線粒體COI序列信息,測序成功率為42.3%。44個圓形魚卵中有27個測序成功,測序成功率達61.4%;27個橢圓形魚卵中有3個測序成果,測序成功率僅為11.1%。30個魚卵線粒體COI序列分別于BOLD數據庫比對分析,結果顯示與BOLD數據庫中參考序列相似性在99.6~100%之間,30個魚卵均能鑒定到種的水平,分屬于1個目,7個科,13個種(表1,表2)。刺尾魚科(Acanthuridae)有5個種,分別為白面刺尾魚(Acanthurusnigricans)、扁體櫛齒刺尾魚(Ctenochaetusstrigosus)、雙斑櫛齒刺尾魚(C.binotatus)、白尾櫛齒刺尾魚(C.flavicauda)和黑背鼻魚(Nasolituratus),共有19個魚卵,占鑒定魚卵總數的63.3%。鸚哥魚科(Scaridae)有2個種,分別為史氏鸚哥魚(Scarusschlegeli)和斑點鯨鸚哥魚(Cetoscarusocellatus)。隆頭魚科(Labridae)有2個種,分別為單帶尖唇魚(Oxycheilinusunifasciatus)和雙線尖唇魚(O.digramma)。鲹科(Carangidae)有1個種為藍圓鲹(Decapterusmaruadsi);籃子魚科(Siganidae)有1個種為銀籃子魚(Siganusargenteus);笛鯛科(Lutjanidae)有1個種為欖色細齒笛鯛(Aphareusfurca);鰈魚科(Zanclidae)有1種為鐮魚(Zancluscornutus)。雙斑櫛齒刺尾魚有10個魚卵,白尾櫛齒刺尾魚有4個魚卵,這兩個種為該海域的優(yōu)勢種類,其余各種類均為1~2個魚卵。
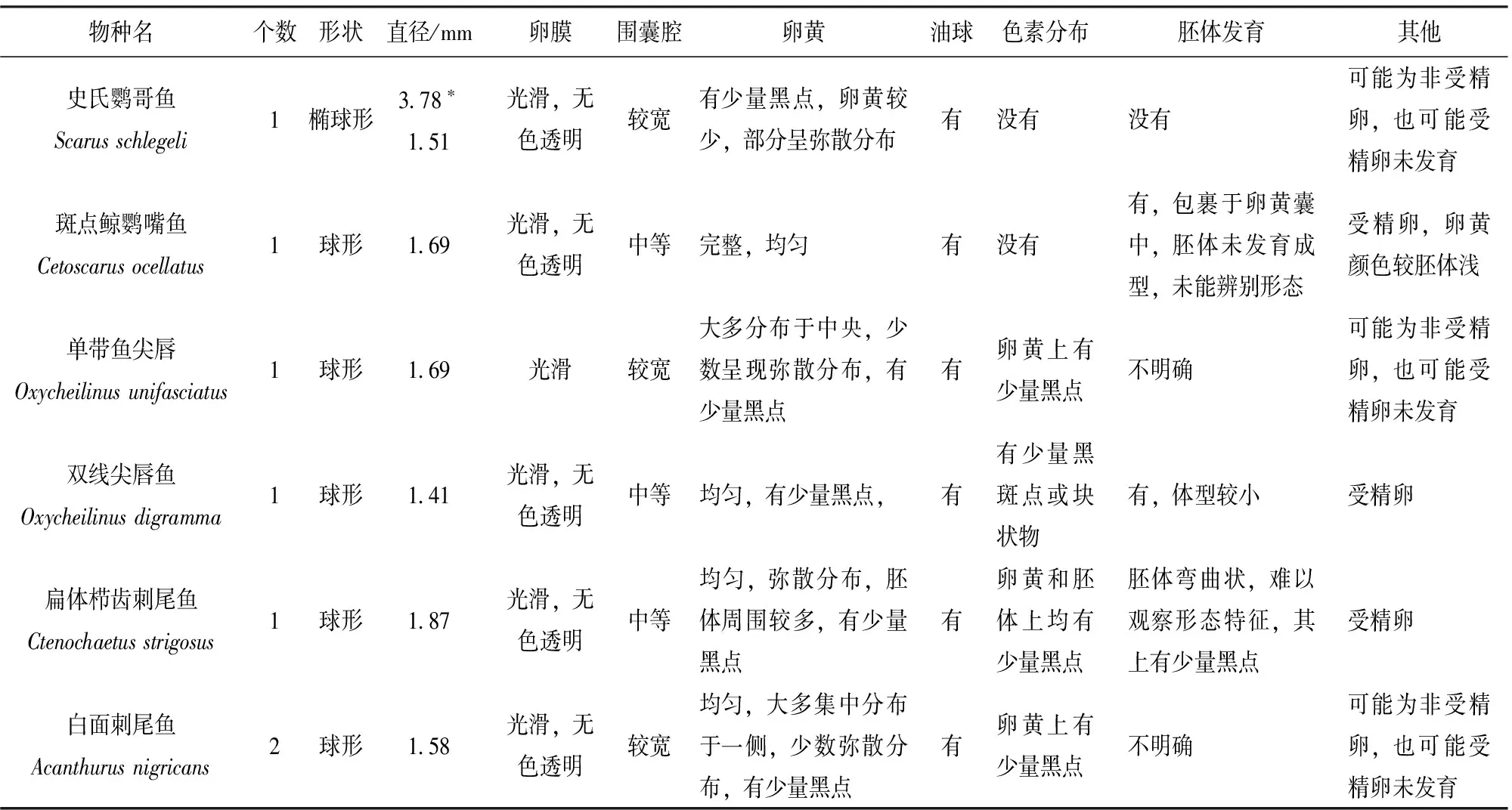
表1 13種魚卵的形態(tài)特征Tab.1 Morphological features of 13 species of fish eggs
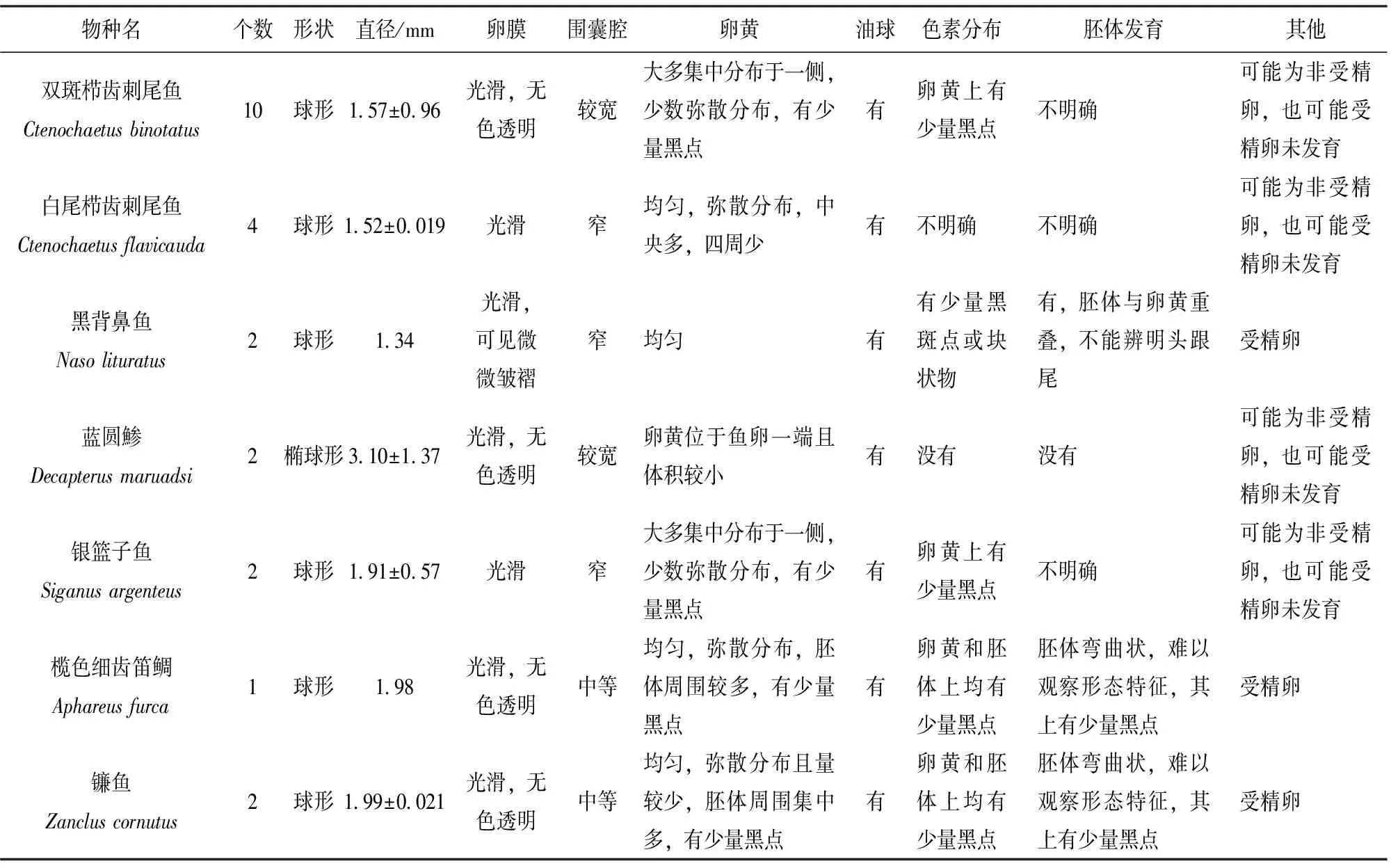
續(xù)表1

表2 南海永暑礁海域魚卵mtCOI序列比對結果Tab.2 Sequences comparison results of mtCOI in fish eggs from Yongshu reef lagoon of South China Sea
2.2 系統(tǒng)進化分析
獲得的30個魚卵的線粒體COI序列長度在463~662 bp之間,序列中沒有插入/缺失位點和終止密碼子。根據線粒體COI序列計算種內的K2P遺傳距離為0~0.002,同一個屬種間的遺傳距離為0.004~0.132,而同一個科不同屬間的遺傳距離為0.117~0.185。根據K2P模型利用遺傳距離構建的鄰接系統(tǒng)進化樹顯示,相同的種類聚為一個分支;刺尾魚科中的刺尾魚屬和櫛齒刺尾魚屬聚為一支,而與同一科的鼻魚屬未形成單系支(圖1)。
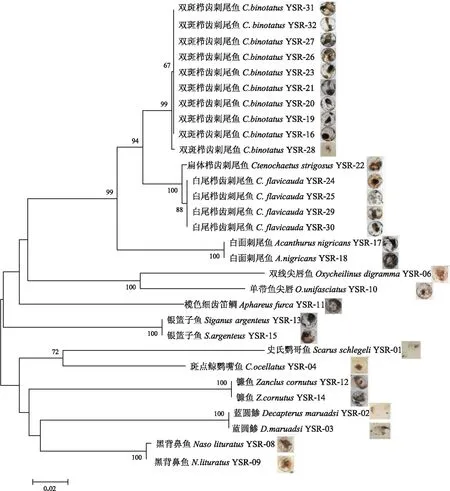
圖1 根據K2P模型利用遺傳距離構建的鄰接系統(tǒng)進化樹Fig.1 Neighbor-joining tree constructed based on K2P model
2.3 魚卵形態(tài)特征分析
根據魚卵的形狀、卵膜、圍囊腔、卵黃、油球、色素分布和胚胎發(fā)育情況等形態(tài)性狀,詳細描述了該研究鑒定的13種魚類的魚卵形態(tài)特征(表1)。史氏鸚哥魚和藍圓鲹的魚卵均為橢圓形,卵膜光滑,無色透明,圍囊腔較寬,但是史氏鸚哥魚卵徑較大,卵黃均勻分布,而藍圓鲹的卵黃位于魚卵的一端,并且體積較小。其余11種魚類的魚卵均為圓形,魚卵直徑在1.34~1.99 mm之間,黑背鼻魚的魚卵直徑最小,鐮魚的魚卵直徑最大。已有的研究表明海水魚卵直徑在0.5~5.5 mm之間[18],該研究所獲得的永暑礁海域魚卵直徑也處于該范圍,但是比尤卡坦半島(Yucatan Penisula)海域魚卵直徑大[2],可能是由于其將魚卵保存于酒精中導致魚卵縮水造成的,而該研究是將魚卵樣品直接冷凍保存。
3 討論
近年來,由于過度捕撈和填海造島等人類活動干擾,漁業(yè)生態(tài)環(huán)境發(fā)生改變,導致漁業(yè)資源顯著減少[19]。對魚卵進行準確鑒別是了解魚類繁殖習性、生活史和種群動態(tài)的基礎,可用于鑒別魚卵的形態(tài)特征較少,傳統(tǒng)的分類手段很難準確鑒別到種的水平。永暑礁為典型的珊瑚環(huán)礁,是熱帶珊瑚礁魚類的重要棲息地和產卵場,目前未見有關永暑礁魚卵分類鑒別的相關文獻報道。DNA條形碼技術的迅速發(fā)展,為從分子水平準確鑒定魚卵提供了有效的手段[10,20]。從魚卵中提取高質量的DNA用于PCR擴增,是利用DNA條形碼鑒別魚卵的前提條件,該研究分析的71個冷凍保存的魚卵中30個成功擴增測序獲得了有效的線粒體COI序列信息,成功率為42.3%。部分魚卵未能成功獲得COI序列,可能由于魚卵樣品為冰凍保存,樣品在凍融過程中降解,導致無法提取其基因組DNA,或者由于魚卵樣品體積較小,在DNA提取過程中操作不當導致樣品損失,因此,魚卵樣品保存和基因組DNA的提取需要進一步的優(yōu)化,提高魚卵鑒定的檢出率。有趣的是圓形魚卵的擴增測序成功率(61.4%)顯著高于橢圓形魚卵的擴增測序成功率(11.1%),其中原因有待于進一步研究。酒精保存魚卵容易導致魚卵脫水,致使魚卵縮小變形,但是酒精保存不需要特殊的設備,容易操作,也是目前魚卵采集過程中的主要保存方式之一,Leyva等[2]研究表明酒精保存的魚卵DNA條形碼測序成功率為46.3%,Burghart等[9]的研究表明50%的異丙醇保存魚卵也不影響樣品用于DNA條形碼分析。由此可見目前魚卵的擴增成功率并不高,如何開發(fā)高效的魚卵保存和擴增方法是有待進一步研究的問題。
近年來有關魚類DNA條形碼參考序列數據庫構建得到了快速發(fā)展,Díaz等[21]采集分析了79種魚類樣品,測序分析了其DNA條形碼序列,構建了美洲Lower Paraná河魚類DNA條形碼參考序列數據庫;Chang等[22]采集分析了1 245種魚類憑證標本及其DNA條形碼序列,構建了臺灣海峽魚類DNA條形碼參考序列數據庫;Weight等[23]采集分析了521種魚類樣品及其DNA條形碼序列,構建了加勒比海魚類DNA條形碼參考序列數據庫;中國水產科學研究院依托國家科技基礎工作專項采集分析了763種南海魚類憑證標本及其DNA條形碼序列,構建了較為完備的南海珊瑚魚類DNA條形碼數據庫,這些序列均已錄入到BOLD條形碼數據庫中,這些條形碼序列數據庫的建立為基于DNA條形碼技術開展魚卵和仔稚魚鑒定奠定了基礎。北大西洋大陸架海域93.26%的魚卵DNA條形碼序列能夠與BOLD數據庫中的參考序列相匹配[8],在加勒比海珊瑚礁海域僅有85.7%的魚卵DNA條形碼序列能夠與BOLD數據庫中的參考序列相匹配[2],該研究中100%的魚卵DNA條形碼序列能夠與參考數據庫中的序列相匹配。這些研究結果表明DNA條形碼參考數據庫的完整性直接影響到魚卵鑒定的效率。
由于線粒體COI基因存在同義突變偏好性,不同種屬間的突變率存在顯著差異[24],致使種內或種、屬間的線粒體COI序列多態(tài)性存在顯著差異,這種差異稱為條形碼間隙(Barcode gap)[25],DNA條形碼間隙的存在是利用條形碼進行物種鑒定的前提,已有的關于魚類DNA條形碼研究證實魚類種內或種、屬間存在顯著的條形碼間隙[22,26]。該研究表明永暑礁同種魚類間的K2P遺傳距離為0~0.002,同一個屬不同種間的遺傳距離為0.004~0.132,屬間的遺傳距離為0.117~0.185,存在著明顯的DNA條形碼間隙,分析的所有種類在NJ進化樹中均能形成單系支,證實了DNA條形碼技術適用于南海珊瑚礁魚類魚卵和仔稚魚鑒定。該研究中南沙珊瑚礁魚類種內遺傳距離最高僅為0.002,顯著低于墨西哥尤卡坦半島魚類種內遺傳距離(0.006)[2]和臺灣海峽魚類種內遺傳距離(0.151)[21],可能與該研究中每個種類使用魚卵的數量較少有關,下一步應加大魚卵采集和分析的數量,以便更準確反映種內和種屬間的遺傳關系。
[1]周美玉,陳 驍,楊圣云.采用DNA條形碼技術對廈門海域魚卵、仔稚魚種類的鑒定[J].海洋環(huán)境科學,2015,34(1):120-135.
[2]Leyva C E,Vasquez Y L,Carrillo L,et al.Identifying pelagic fish eggs in the southeast Yucatan Peninsula using DNA barcodes[J].Genome,2016,59:1117-1129.
[3]Steinke D ,Connell A D,Hebert P D N.Linking adults and immatures of South African marine fishes[J].Genome,2016,59(11):959-967.
[4]萬瑞景,張仁齋.中國近海及其鄰近海域魚卵與仔稚魚[M].上海:上海科學技術出版社,2016:1-435.
[5]Zeng Y,Wu Z,Zhang C,et al.DNA barcoding of Mobulid Ray Gill Rakers for Implementing CITES on Elasmobranch in China[J].Sci Rep,2016,6:37567.
[6]Sakinan S,Karahan A,Ok M.Integration of DNA barcoding for the initial recordings of Lessepsian fishes:a case study of the Indo-Pacific slender ponyfish Equulites elongates[J].Fish Biol,2017,90(3):1054-1061.
[7]Bhatta C M,Sharma A R,Patra B C,et al.DNA barcoding to fishes:current status and future directions[J].Mitochondrial DNA,2016,27(4):2744-2752.
[8]Leah A,Lewis D E,Richardson E V,et al.Intergrating DNA barcoding of fish egg into ichthyoplankton monitoring programs[J].Fish Bull,2016,114:153-165.
[9]Burghart S E,Woudenberg L V,Daniels C A,et al.Disparity between planktonic fish egg and larval communities as indicated by DNA barcoding[J].Mar Ecol Prog Ser,2014,503(1):195-204.
[10]Harada A E,Lindgren E A,Hermsmeier M C,et al.Monitoring spawning activity in a southern california marine protected area using molecular identification of fish eggs[J].PLoS ONE,2015,10(8):e0134647.
[11]Carole C B,Johnson G D.Connectivity across the Caribbean Sea:DNA barcoding and morphology unite an enigmatic fish larva from the Florida straits with a new species of sea bass from deep reefs off Curacao[J].PLoS ONE,2014,9(5):e97661.
[12]中國科學院動物研究所等.南海魚類志[M].北京:科學出版社,1962:1-1184.
[13]張仁齋,李昌華.洋浦灣的魚卵及其仔魚調查(一)[J].水產學報,1965,2(4):51-62.
[14]張仁齋,陸穗芬.短尾大眼鯛卵及仔、稚魚的發(fā)育[J].水產學報,1982,6(3):243-251.
[15]Ward R D,Zemlak T S,Innes B H,et al.DNA barcoding Australia’s fish species[J].Phil Trans R Soc B,2005,360:1847-1857
[16]Ratnasingham S,Hebert P D N.A DNA-based registry for all animal species:The barcode index number (BIN)System[J].PLoS ONE,2013,8(8):e66213.
[17]Kumar S,Stecher G,Tamura K.MEGA7:Molecular evolutionary genetics analysis version 7.0 for bigger datasets[J].Mol Biol Evol,2016,33(7):1870-1874.
[18]Matarese A C,Sandknop E M.Identification of fish eggs//Ahlstrom E H,ed.Ontogeny and systematics of fishes[M].California:Spec Publ Am Soc Ichthyol Herpetol,1984:27-31.
[19]湯嬌雯,張 富,陳兆波.我國魚類生物多樣性保護策略[J].淡水漁業(yè),2009,39(4):76-77
[20]Mabuchi K.A method for PCR-based identication of fish eggs attached to vegetation in lake Biwa,Japan[J].Conserv Genet Resour,2016,(8):215-217.
[21]Diaz J,Villanova G V,Brancolini F,et al.First DNA Barcode Reference Library for the Identification of South American Freshwater Fish from the Lower Paraná River[J].PLoS ONE,2016,11(7):e0157419.
[22]Chang C H,Shao K T,Lin H Y,et al.DNA barcodes of the native ray-finned fishes in Taiwan[J].Mol Ecol Resour,2016,DOI:10.1111/1755-0998.12601
[23]Weight L A,Baldwin C C,Driskell A,et al.Using DNA Barcoding to Assess Caribbean Reef Fish Biodiversity:Expanding Taxonomic and Geographic Coverage[J].PLoS ONE,2012,7(7):e41059.
[24]Lane N.Biodiversity:on the origin of bar codes[J].Nature,2009,462:272-274.
[25]Ward R D,Holmes B H.An analysis of nucleotide and amino acid variability in the barcode region of cytochrome c oxidase I (cox1)in fishes[J].Mol Ecol Notes,2007,7:899-907.
[26]Pereira L H,Hanner R,F(xiàn)oresti F,et al.Can DNA barcoding accurately discriminate megadiverse Neotropical freshwater fish fauna[J].BMC Genetics,2013,14:20.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
財經(2017年15期)2017-07-03 22:40:49
財經(2017年2期)2017-03-10 14:35:35
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
