華貴櫛孔扇貝MEF2Cs基因克隆及表達特征分析
2018-03-21 01:07:32朱克誠劉寶鎖郭華陽張殿昌
淡水漁業 2018年2期
朱克誠,劉寶鎖,曹 明,郭華陽,張 楠,張殿昌,2
(1.中國水產科學研究院南海水產研究所,農業部南海漁業資源開發利用重點實驗室, 廣州 510300;2.廣東省海洋生物種業工程技術研究中心,廣州 510300;3.廣東省漁業種質保護中心,廣州 511453)
肌細胞增強因子(Myocyte enhancer factor-2,MEF2)屬于MADS(Agamous deficiens serum response factor,MADS)轉錄因子家族,是一種肌肉特異性轉錄因子。在果蠅及線蟲體內以單基因形式存在,但是在脊椎動物中存在四個家族成員,分別為MEF2A,MEF2B,MEF2C和MEF2D[1]。這四個家族成員能夠通過啟動子的選擇和對mRNA前體的選擇性拼接產生不同種類的增強因子蛋白[2]。四種MEF2主要參與肌肉發育過程中的調節作用,其中MEF2C能調控肌小節的完整性和胚胎后期骨骼肌的正常發育[3]。在小鼠MEF2C基因敲除實驗中,會導致肌小節溶解[3]或心血管異常發育到胚胎期9.5天時死亡[4]。在胚胎發育時期,為了調節組織特異性基因的表達,MEF2C能被很多類型的細胞激活進而調節肌肉組織的功能,諸如:心肌細胞、神經細胞、骨骼肌細胞、線粒體細胞、內皮細胞和免疫細胞[4]。
MEF2C基因最初在人胎兒大腦中發現[5],隨后在骨骼肌和心肌也陸續被報道[6]。盡管MEF2C基因mRNA在組織中廣泛表達,但激活的MEF2C主要分布于心肌、腦、骨骼肌和淋巴細胞組織等。以往該基因的研究主要集中在脊椎動物不同組織中的表達分析[7-10],很少涉及到無脊椎動物中的功能研究。在無脊椎動物中,僅僅克隆了長牡蠣(Crassostreagigas)、溫室希蛛(Parasteatodatepidariorum)和美洲鱟(Limuluspolyphemus)等的MEF2C基因,但是沒有組織表達譜的相關報道。目前還未見到華貴櫛孔扇貝MEF2C基因的相關研究報道。
華貴櫛孔扇貝(Chlamysnobilis)隸屬軟體動物門瓣鰓綱珍珠目扇貝科,廣泛分布于中國南海、日本、越南以及印度尼西亞等沿海地區。華貴櫛孔扇貝為暖水性附著型貝類,養殖周期較短、死亡率低、肉質鮮美,是一種重要水產經濟貝類[11]。目前對于華貴櫛孔扇貝的研究多集中在人工育苗、分子輔助育種、殼色和閉殼肌顏色、溫度和鹽度對其酶活和幼貝存活的影響等方面[12-14]。此外,在一些雙殼類物種中,其生長能受到養殖環境中鹽度的影響[15-16]。可見鹽度能影響肌肉的生長,間接影響MEF2C基因的表達水平,而華貴櫛孔扇貝MEF2C基因功能研究還未見報道。本實驗克隆了MEF2C和MEF2C-like基因序列,分析了其時空表達和親緣關系,初步了解以上基因在不同鹽度下的表達量特征,其結果可為探索最佳的鹽度生長提供基礎資料。
1 材料與方法
1.1 試驗材料
華貴櫛孔扇貝胚胎和幼貝取自海南三亞某養殖場,全部暫養于中國水產科學研究院南海水產研究所海南熱帶水產研究開發中心實驗基地,隨機挑選一定數量的健康且閉殼肌活力旺盛的個體[體質量(25.11±2.85)g;殼長(5.51±0.15)cm;殼寬(5.12±0.12 cm)],清洗貝殼表面的污物,早晚各投喂1次亞心型扁藻(Platymonassubcordiformis),日換水50%。放養在300 L的水泥池內,循環水養殖,溫度為(22±0.5)℃,暫養2周后用于試驗。隨機抽取5個健康的華貴櫛孔扇貝,冰浴麻醉后取外套膜、腹足、消化腺、鰓、心臟、閉殼肌、性腺,立即放入液氮中速凍,然后放置于-80 ℃冰箱中備用。
針對華貴櫛孔扇貝不同發育階段(受精卵、囊胚期、原腸胚期、擔輪幼蟲期和D-型幼蟲期)取樣,每個時期至少取20個個體;針對華貴櫛孔扇貝不同生長階段(30 phd、40 phd、60 phd、120 phd和180 phd)的閉殼肌取樣,每個時期至少取5個個體,立即放入液氮中速凍,然后放置于-80 ℃冰箱中備用。
1.2 試驗方法
選擇上述暫養后健康的華貴櫛孔扇貝,隨機挑選270個分別放在9個水泥池中,每池30個。三個組設定的鹽度分別為22 、28 和 34 。試驗為期一周,試驗結束后將貝冰浴麻醉后取閉殼肌,立即放入液氮中速凍,然后放置于-80 ℃冰箱中備用。
1.3 總RNA的提取和逆轉錄
根據TRIzol說明書提取華貴櫛孔扇貝外套膜、腹足、消化腺、鰓、心臟、閉殼肌、性腺等組織RNA。RNA的質量和總量用NanoDrop 2000 分光光度計(ThermoScientific,美國)檢測,并測定RNA的OD值。取2 μg RNA合成 cDNA(用Takara試劑盒PrimeScript RT reagent Kit with gDNA Eraser)用于實時熒光定量PCR(qRT-PCR)。
1.4 MEF2C和MEF2C-like基因克隆
在該實驗室前期研究基礎中,得到了華貴櫛孔扇貝轉錄組序列的注釋結果(未發表),從中獲得華貴櫛孔扇貝MEF2C和MEF2C-like基因預測的序列。針對該序列,運用Primer Premier 5.0軟件,設計引物來分別驗證上述基因的ORF序列(表1)。PCR擴增總體系為25 μL,其中去離子水18.2 μL,10×Ex buffer 2.5 μL,dNTP 2 μL,引物各0.5 μL,cDNA模板為1 μL,ExTaq酶為0.3 μL。PCR擴增條件:94 ℃預變性 3 min,94 ℃變性 1 min,56 ℃退火30 s,72 ℃延伸1 min,30個循環;72 ℃延伸10 min,4 ℃保存。取擴增產物5 μL用1%的瓊脂糖凝膠電泳檢測,切膠回收PCR產物,然后將膠回收產物與pMD18-T載體(TaKaRa公司)16 ℃連接過夜,接著轉化到感受態細胞(DH5α)中,進行陽性克隆篩選并測序。
1.5 生物信息學分析
用BLAST program(http://blast.ncbi.nlm.nih. gov/Blast.cgi)來搜索氨基酸和核苷酸序列的同源序列;利用ORF Finder(http://www.ncbi.nlm.nih. gov/projects/gorf/)來預測MEF2C和MEF2C-like基因的ORF;使用ExPASy程序(http://prosite.expasy.org/)和SignaIP 4.1 Server對MEF2C和MEF2C-like氨基酸等電點、分子質量和信號肽等功能基序進行預測;通過使用Clustalw2程序(http://www.ebi.ac.uk/Tools/msa/clustalw2/)進行多重序列比對,使用MEGA 6.0軟件構建系統進化樹,對MEF2C和MEF2C-like與其他物種的同源性和親緣關系進行比對分析[17]。
1.6 MEF2C和MEF2C-like 基因表達分析
實時熒光定量PCR(qRT-PCR)用于分析MEF2C和MEF2C-like基因在不同組織和不同時期的mRNA表達量。針對MEF2C和MEF2C-like基因的ORF序列,跨內含子區域來設計熒光定量引物(表1)。內參引物β-actin-F/R參照Lu等[18],qRT-PCR的程序參照Zhu等[19]。此外,該研究中使用2-ΔΔCT法來分析基因的相對表達量[20]。MEF2C和MEF2C-like基因的表達量用平均值±方差來表示(mean ± SE)。用SPSS statistics 18.0 進行單因素方差分析,差異顯著性用P<0.05來表示。
2 結果
2.1 華貴櫛孔扇貝MEF2C和MEF2C-like基因克隆和序列分析
MEF2C和MEF2C-like基因序列已驗證。華貴櫛孔扇貝MEF2C基因的cDNA全長為1 506 bp(GenBank no.MF614124),開放閱讀框為1 302 bp,編碼433個氨基酸。預測的MEF2C蛋白的相對分子質量和等電點分別為47×103和8.83,其疏水性平均值(Ggrand average of hydropathicity,GRAVY)為 -0.908。MEF2C信號肽(SignalP)預測結果表明沒有信號肽存在于推測的氨基酸序列中。此外,MEF2C-like基因ORF全長為1 158 bp(GenBank no.MF614125),編碼385個氨基酸。預測的MEF2C-like蛋白的相對分子質量和等電點分別為41×103和8.58,其疏水性平均值(Ggrand average of hydropathicity,GRAVY)為 -0.541。同樣地,MEF2C-like信號肽(SignalP)預測結果表明也沒有信號肽存在于推測的氨基酸序列中。如MEF2家族中的其他成員,蛋白結構預測結果表明兩種MEF2Cs都含有保守的MADS-Box(氨基酸 1-58)和鄰近保守的MEF2結構域(氨基酸59-86)(圖1)。此外,在MEF2C和MEF2C-like中,N-末端較C-末端高度保守。

表1 實驗中所需引物序列Tab.1 Primers used for cloning and expression

圖1 華貴櫛孔扇貝MEF2C(MF614124)和MEF2C-like(MF614124)的氨基酸序列與其它物種MEF2C 和 MEF2C-like的氨基酸多重序列比對Fig.1 Comparison of deduced amino acid sequences of C.nobilis of MEF2C (MF614124)and MEF2C-like (MF614124)with published MEF2C-like MADS 和 MEF2結構域分別標記的是紅色和藍色。表2列出了所有物種的Genebank序列號。
2.2 進化分析
為了探究MEF2C基因的同源性和進化特征,搜索的MEF2C直系同源基因數據后用MEGA6.0進行比對。基于鄰位相連法(Neighbor-Joining,NJ)分析結果表明華貴櫛孔扇貝和長牡蠣親緣關系最近,其次是蝦夷扇貝等軟體動物(圖2)。此外,結果也顯示了軟體動物MEF2C與節肢動物(溫室希蛛和美洲鱟)MEF2C聚為一支,然后無脊椎動物(軟體動物和節肢動物)再與脊椎動物聚在一起。這些結果符合傳統的分類學。
經過Blast對比分析發現,華貴櫛孔扇貝MEF2C氨基酸序列與長牡蠣、蝦夷扇貝MEF2C氨基酸序列同源性最高,分別為67%和58%;與安水金線鲃MEF2C-like-1和MEF2C-like-2的同源性最低,分別為34%和33%(表2)。
2.3 MEF2C和MEF2C-like基因時空表達分析
MEF2C和MEF2C-like基因在胚胎和幼體不同發育時期中的表達量類似,隨著發育時期的推移,表達量顯著增加,在D型幼蟲期達到最高值(圖3A),隨后表達量都下降。另外,在不同組織中的表達量分析結果表明MEF2C和MEF2C-like基因也具有相似的表達趨勢,在所有組織中都有表達,其中在閉殼肌中的表達量顯著高于其他組織(圖 3B),外套膜和消化腺次之,腹足和雄性腺中表達量最低。
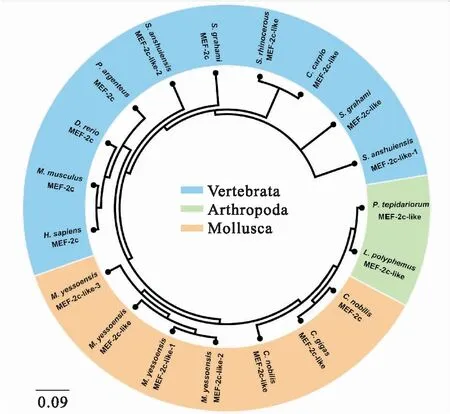
圖2 用MEGA6.0 鄰位相連法構建的MEF2C氨基酸序列系統進化樹Fig.2 Phylogenetic trees of C.nobilis MEF2C relative to the homologues of other species from the amino acid datasets using the MEGA 6.0 software with Neighbor-Joining (NJ)method

表2 預測的華貴櫛孔扇貝MEF2C氨基酸序列與其他物種MEF2C氨基酸序列的比較Tab.2 Comparison of deduced amino acid region of C.nobilis MEF2C with MEF2C of other species
在華貴櫛孔扇貝閉殼肌不同生長時期中,MEF2C和MEF2C-like基因的表達量顯著上升至孵化后60和120 d達到最大,隨后孵化后180 d顯著下降(圖4)。在不同鹽度養殖的試驗中,MEF2C和MEF2C-like基因的表達量在鹽度為28下顯著高于其他兩個鹽度組,其中在28 下,MEF2C的mRNA水平分別是22 和34的4.24倍和1.80倍,MEF2C-like的mRNA水平分別是22和34的6.14倍和1.60倍(圖5)。
3 討論
MEF2C是MEF2基因家族成員之一,參與調控細胞増殖和肌肉分化的重要調節因子,在脊椎動物的功能較為清晰,但是在貝類等有殼的水生動物中的功能還沒有完全被闡明。華貴櫛孔扇貝MEF2C含有1 302 bp核苷酸,編碼433個氨基酸;MEF2C-like含有1 074 bp ORF,編碼358個氨基酸;這兩個基因的氨基酸同源相似性高達68%。序列比對分析表明華貴櫛孔扇貝MEF2C和MEF2C-like含有高度保守的MADS和MEF2結構域,這與鴨和哺乳動物在N-末端的結構類似[21]。這兩個高度保守的結構域結合在DNA共有序列CTA(A/T)4TAG上,才能有效地進行聚合作用[22]。華貴櫛孔扇貝MEF2C和MEF2C-like包含有脊椎動物典型的HJURP-C結構域,可能是由于MEF2C在進化過程中脊椎動物為了適應新環境而演變出不同的功能。華貴櫛孔扇貝MEF2C氨基酸與其他物種MEF2C氨基酸的同源性范圍為33%~68%,其中與軟體動物親緣關系最近,范圍為58%~68%,這與其他基因在無脊椎動物中序列差異性較大觀點一致。與其他后生動物的氨基酸序列比對分析結果說明華貴櫛孔扇貝MEF2Cs與長牡蠣的親緣關系最近,這可能是因為華貴櫛孔扇貝和長牡蠣都屬于扇貝科。然而脊椎動物和軟體動物的氨基酸序列差異較大(圖2, 表 2),這也基本符合傳統生物學分類的結果。
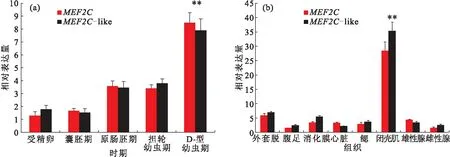
圖3 MEF2C 和 MEF2C-like在華貴櫛孔扇貝不同發育時期(A)和不同組織(B)中的表達量Fig.3 Expression patterns of MEF2C and MEF2C-like at different development stages (A)and tissues (B)of C.nobilis P<0.01表示具有顯著差異性。圖4,5同。
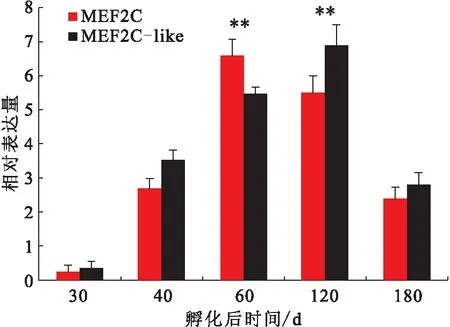
圖4 MEF2C 和 MEF2C-like在華貴櫛孔扇貝不同閉殼肌生長時期的表達量Fig.4 Expression level of MEF2C and MEF2C-like at different muscle development stages of C.nobilis
MEF2C廣泛表達于體內的各種細胞中,尤其是調節心肌細胞、骨骼肌細胞、平滑肌細胞、線粒體細胞、免疫細胞、神經細胞和內皮細胞發育的核心元件[23]。MEF2C基因的研究主要集中在脊椎動物中,只有少量的研究在軟體動物中,尤其是扇貝科。該研究結果表明隨著發育時期的推移,MEF2C表達量逐漸升高,在D型幼蟲期達到最高,隨后表達量顯著下降。這可能是在D型幼蟲期閉殼肌前體開始萌芽形成。此外,組織特異性表達譜揭示了兩種MEF2Cs在各個組織中都廣泛表達,其中顯著表達于閉殼肌組織中,這與山羊的研究結果類似[24]。在胚胎形成時期,MEF2C基因涉及到建立肌源性的細胞系[3]。Chen等[24]人的研究表明山羊MEF2C的mRNA表達量在出生后3 d時達到最高水平,隨后下降,到30~120 d又是一個高峰值。在該研究中,MEF2C和MEF2C-like基因的表達量在60~120 d時達到最高峰,可能是因為這個時期是閉殼肌的一個快速生長時期,而肌肉發育調節重要的因子MEF2Cs表達量處于一個高峰期。

圖5 在不同鹽度下,MEF2C 和 MEF2C-like在華貴櫛孔扇貝閉殼肌中的表達量Fig.5 Expression patterns of MEF2C and MEF2C-like at different salinity groups of C.nobilis
在養殖過程中影響雙殼類閉殼肌質量的因素很多,尤其是鹽度。先前的研究表明最佳的組合是溫度為25.7 ℃/29 ℃,可以使孵化率高達72.42%[25]。在對華貴櫛孔扇貝幼蟲存活影響實驗中,鹽度為28.03 ,20日齡存活率為48.25%[26]。劉志剛等[27]在針對華貴櫛孔扇貝幼貝生長的研究中表明最適生長鹽度為21.9~34.0 之間。翡翠貽貝稚貝階段,鹽度為20~25時生長速度最快[16]。然而,MEF2C涉及到肌肉發育的正調控。在該研究中,MEF2C和MEF2C-like基因的mRNA表達量在鹽度為28中顯著高于其他鹽度組,說明在此鹽度是華貴櫛孔扇貝生長的最適鹽度,這與之前的研究結果類似[16,25,27]。可見,為了滿足華貴櫛孔扇貝快速生長的需求,28鹽度是最佳的生長條件。
[1]Morisaki T,Sermsuvitayawong K,Byun S H,et al.MouseMEF2Bgene:unique member ofMEF2 gene family[J].J Biochem,1997,122(5):939-946.
[2]Lilly B,Galewsky S,Firulli A B,et al.D-MEF2:a MADS box transcription factor expressed in differentiating mesoderm and muscle cell lineages during Drosophila embryogenesis[J].Proc Natl Acad Sci USA,1994,91(12):5662-5666.
[3]Potthoff M J,Arnold M A,Mcanally J,et al.Regulation of skeletal muscle sarcomere integrity and postnatal muscle function byMEF2C[J].Mol Cell Biol,2007,27(23):8143-8151.
[4]Naya F J,Black B L,WU H,et al.Mitochondrial deficiency and cardiac sudden death in mice lacking theMEF2Atranscription factor[J].Nat Med,2002,8(11):1303-1309.
[5]Leifer D,Krainc D,Yu Y T,et al.MEF2C,a MADS/MEF2-family transcription factor expressed in a laminar distribution in cerebral cortex[J].Proc Natl Acad Sci USA,1993,90(4):1546-1550.
[6]Lyons G E,Micales B K,Schwara J,et al.Expression ofMEF2Cgenes in the mouse central nervous system suggests a role in neuronal maturation[J].J Neurosci,1995,15(8):5727-5238.
[7]Tessier S N,Storey K B.Myocyte enhancer factor-2 and cardiac muscle gene expression during hibernation in thirteen-lined ground squirrels[J].Gene,2012,501(1):8-16.
[8]Aguilar O A,Hadj-moussa H,Storey K B.Freeze-responsive regulation of mef2 proteins and downstream gene networks in muscles of the wood forg,rana sylvatica[J].J Therm Biol,2017,67:1-8.
[9]Ganassi M,Badodi S,Polacchini A,et al.Distinct functions of alternatively spliced isoforms encoded by zebrafishMEF2Ca andMEF2Cb[J].Biochim et Biophys Acta,2014,1839(7):559-570.
[10]Leifer D,Golden J,Kowall N W.Myocyte-specific enhancer binding factor 2C expression in human brain development[J].Neuroscience,1994,63(4):1067-1079.
[11]Lou Y.Fisheries and aquaculture:China//Scallops,Ecology and Aquaculture (Shumway S E)[M].B.V.Amsterdam:Elsevier Science Publishers,1991:809-824.
[12]錢佳慧,栗志民,申玉春,等.溫度和鹽度對華貴櫛孔扇貝抗氧化酶活性的聯合效應研究[J].南方水產科學,2015,11(6):49-57.
[13]Liu H,Zheng H,Wang S,et al.Cloning and functional characterization of a polyunsaturated fatty acid elongase in a marine bivalve noble scallopChlamysnobilisReeve[J].Aquaculture,2013,416(5):146-151.
[14]Wang Y,Fu D,Xia J.The genetic diversity of the noble scallop (Chlamysnobilis,Reeve 1852)in China assessed using five microsatellite markers[J].Mar Genom,2013,9(3):63-67.
[15]O’connor W A,Lawler N F.Salinity and temperature tolerance of embryos and juveniles of the pearl oyster,Pinctada imbricate Roding[J].Aquaculture,2004,229(1):493-506.
[16]楊 鵬,閆喜武,韓 華,等.鹽度對翡翠貽貝受精卵孵化及幼蟲和稚貝生長和存活的影響[J].大連海洋大學學報,2013,28(6):549-552.
[17]Tamura K,Stecher G,Peterson D,et al.MEGA6:Molecular Evolutionary Genetics Analysis version 6.0[J].Mol Biol Evol,2013,30(12):2725-2729.
[18]Lu Y Q,Zheng H P,Zhang H K,et al.Cloning and differential expression of a novel toll-like receptor gene in scallopChlamysnobiliswith different total carotenoid content[J].Fish Shellfish Immunol,2016,56:229-238.
[19]Zhu K C,Zhang N,Guo H Y,et al.Molecular characterization of MyD88 inPinctadafucataand its response to lipopolysaccharides and polyinosinic-cytidylic acid[J].Israeli J Aquacult-bamidgeh,2016,68:1-8.
[20]Livak K J,Schmittgen T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod[J].Methods,2001,25:402-408.
[21]Wu Y,Dey R,Han A,et al.Structure of the MADS-box/MEF2 domain ofMEF2Abound to DNA and its implication for myocardin recruitment[J].J Mol Biol,2010,397(2):520-533.
[22]Yu Y,Breitbart R,Smoot L,et al.Human myocyte-specific enhancer factor 2 comprises a group of tissue-restricted MADS box transcription factors[J].Genes Dev,1992,6(9):1783-1798.
[23]Chang S,Young B D,Li S,et al.Histone deacetylase 7 maintains vascular integrity by repressing matrix metalloproteinase 10[J].Cell,2006,126:321-334.
[24]Chen L,Cheng B,Li L,et al.The molecular characterization and temporal-spatial expression of myocyte enhancer factor 2 genes in the goat and their association with myofiber traits[J].Gene,2015,555(2):223-230.
[25]Lu W G,Li W D,Ke C H,et al.Reproductive success under the joint influences of temperature and salinity in noble scallop,Chlamysnobilis(Reeve)[J].Aquacult Res,2017,48(2):686-696.
[26]李衛東,呂文剛,王 輝,等.溫度和鹽度對華貴櫛孔扇貝幼蟲存活聯合效應研究[J].水產養殖,2016,37(1):1-8.
[27]劉志剛,劉建勇,楊 博.溫度和鹽度對華貴櫛孔扇貝幼蟲存活與生長的互作效應研究[J].海洋科學,2011,35(10):75-80.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
