干旱年份沙質草地生態系統凈CO2通量年變化特征
2018-01-19 07:30:16牛亞毅李玉強王旭洋龔相文羅永清田德宇
草業學報 2018年1期
牛亞毅,李玉強,王旭洋,龔相文,羅永清,田德宇
(1.中國科學院西北生態環境資源研究院,甘肅 蘭州 730000;2.中國科學院大學,北京 100049)
草地是全球主要自然群系中面積最大的一類,其中溫帶草原生態系統占全球自然植被的32%[1]。近年來有關草地生態系統CO2通量的長期監測得到了快速的發展。很多研究發現草地是全球陸地生態系統中非常重要的碳匯[2-8],例如溫帶和熱帶草地每年吸收0.5 Pg碳[5],青藏高原人工草地不同年際間凈固碳量的范圍為129~513 g·m-2·年-1[6],內蒙古羊草(Leymuschinensis)草地年CO2凈吸收量為126~195 g·m-2[8]。亦有研究表明草地生態系統的碳收支趨于平衡[2,7],例如美國德克薩斯州牧草草地的CO2凈吸收量為0[2]。也有研究發現草地生態系統在年尺度上表現為碳源,例如溫帶半干旱草地和青海三江源退化草地的年CO2釋放量分別為482[9]和30 g·m-2[6]。世界天然草地生態系統中的80%處于中緯度干旱半干旱地區,往往受到水分條件的限制,相對于其他陸地生態系統而言,其碳循環過程對降水(強度、次數和時間分布)變化的響應更為敏感,它所表現的弱源或者弱匯特征在降水的擾動下容易發生方向性的逆轉,從而使得碳的源/匯功能表現出極大的不確定性[10],因此各研究區得出結論也不盡一致。
沙質草地是我國北方干旱半干旱區重要的土地資源,近年來,由于氣候變化和人為干擾等因素的影響使得該區域土地沙漠化日趨嚴重[11],造成植被與土壤碳的流失[12];加之干旱半干旱地區降水條件的限制,可能導致生態系統由碳匯轉變為碳源。對沙質草地生態系統CO2通量動態和碳收支狀況的長期監測和研究,有助于確定沙質草地生態系統的源/匯狀況,為區域尺度碳收支的核算提供基礎數據和科學依據。本研究以地處我國北方半干旱農牧交錯帶的科爾沁沙質草地為研究對象,基于連續兩年的碳通量觀測數據,通過認識沙質草地生態系統凈CO2交換量(NEE)在不同時間尺度的變化特征及其對環境因子變化的響應規律,為區域生態系統碳循環的深入研究提供理論基礎。
1 材料與方法
1.1 研究區概況
研究區位于內蒙古東部科爾沁沙地的奈曼旗境內,依托中國科學院奈曼沙漠化研究站(42°55′ N,120°42′ E,海拔345 m)開展試驗。研究區屬于溫帶大陸性半干旱氣候,年均溫6.4 ℃,≥10 ℃的年積溫為3000~3400 ℃;多年平均降水量360 mm左右[13];無霜期151 d,年均潛在蒸發量1972.8 mm;年平均風速為3.5 m·s-1,大風日數20~60 d。地帶性土壤為沙質栗鈣土,但在風蝕作用下多已退化為風沙土。
1.2 樣地類型
研究樣地為科爾沁典型的沙質草地,地勢平坦,植被較為均一,2015和2016年生長高峰期(8月)的植物生物量如表1所列(地下生物量采樣深度為30 cm)。表層0~20 cm平均土壤有機碳含量為1.27 g·kg-1,全氮含量為0.21 g·kg-1,pH值為7.42,電導率為16.04 μS·cm-1;土壤粗砂(0.10~2.00 mm)含量為92.76%,細砂(0.05~0.10 mm)含量為3.28%,粘粉粒(<0.05 mm)含量為3.96%。主要優勢植物有狗尾草(Setariaviridis)、豬毛菜(Salsolacollina)和五星蒿(Bassiadasyphylla)等。

表1 2015和2016年生長高峰期植物生物量Table 1 The plant biomass during peak growth periods in 2015 and 2016 (g·m-2)
1.3 生態系統碳通量觀測
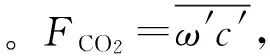
對渦動相關法觀測到的10 Hz原始湍流數據運用Logger Net軟件進行格式轉換(TOB1),然后利用美國Li-cor公司研發的EddyPro軟件進行系列校正計算,包括異常值和野點剔除、坐標旋轉修正、頻率損失修正、超聲虛溫修正、空氣密度效應修正等基本質量控制,初步得到間隔為30 min的通量數據,然后依據下列原則進一步剔除異常數據:1)剔除降水期間的數據;2)根據3倍標準差剔除異常突出數據,由原始時間序列x求相鄰點之差Δx的總體標準差(σΔx),逐點檢查,如某點Δx≥3σΔx,則為野點值,若連續6點都符合以上判斷,則不做野點值處理;3)利用凈輻射(Rn)小于1 W·m-2判斷出夜間數據,對夜間數據,用一個給定的臨界閾值μc*加以檢驗,當μ*<μc*時,則剔除該通量數據,μc*一般的取值范圍為0.10~0.20 m·s-1,本研究取0.10 m·s-1。通過上述質量控制后,對缺失的CO2通量數據進行插補:對于缺失小于2 h的數據采用線性內插法,缺失大于2 h的數據采用平均日變化法(夜間以7 d為周期,白天以14 d為周期)[15],最后得到以30 min為間隔,連續的、質量可靠的數據集。
1.4 數據統計分析
應用SPSS 19.0軟件進行數據統計分析,NEE與環境因子(空氣溫度、5 cm土壤溫度、5 cm土壤體積含水量)之間的關系用皮爾遜相關法進行分析。采用Microsoft Excel 2010軟件繪圖。
2 結果與分析
2.1 環境因子變化
溫度和降水均表現出明顯的季節變化(圖1a)。2015和2016年空氣溫度(Ta)日均值的變化范圍分別為-17.64~27.81 ℃和-26.02~30.29 ℃;最低月(1月)平均氣溫分別為-14.66和-10.02 ℃,最高月(7月)平均氣溫分別為23.23和24.15 ℃。年均溫2015年(7.80 ℃)稍高于2016年(7.25 ℃)。5 cm土壤溫度(Ts)日均值的變化范圍分別為-14.55~29.69 ℃和-18.07~31.36 ℃,1月的平均溫度分別為-8.86和-13.67 ℃,7月的平均溫度分別為26.56和27.49 ℃;Ts全年均值2015和2016年分別為9.51和9.39 ℃。
2015和2016年降水總量分別為212.3和277.2 mm(圖1b),均明顯低于該地區歷史多年平均降水量(360 mm);降水頻率2015年(70次)低于2016年(85次);降水分配主要集中在植物生長期5-9月(>80%)。5 cm土壤體積含水量(SWC):2015年的最低值(1.66%)出現在1月,最高值(7.07%)出現在9月;2016年的最低值(1.73%)出現在12月,最高值(8.81%)出現在5月;年均值分別為3.37%和3.77%。
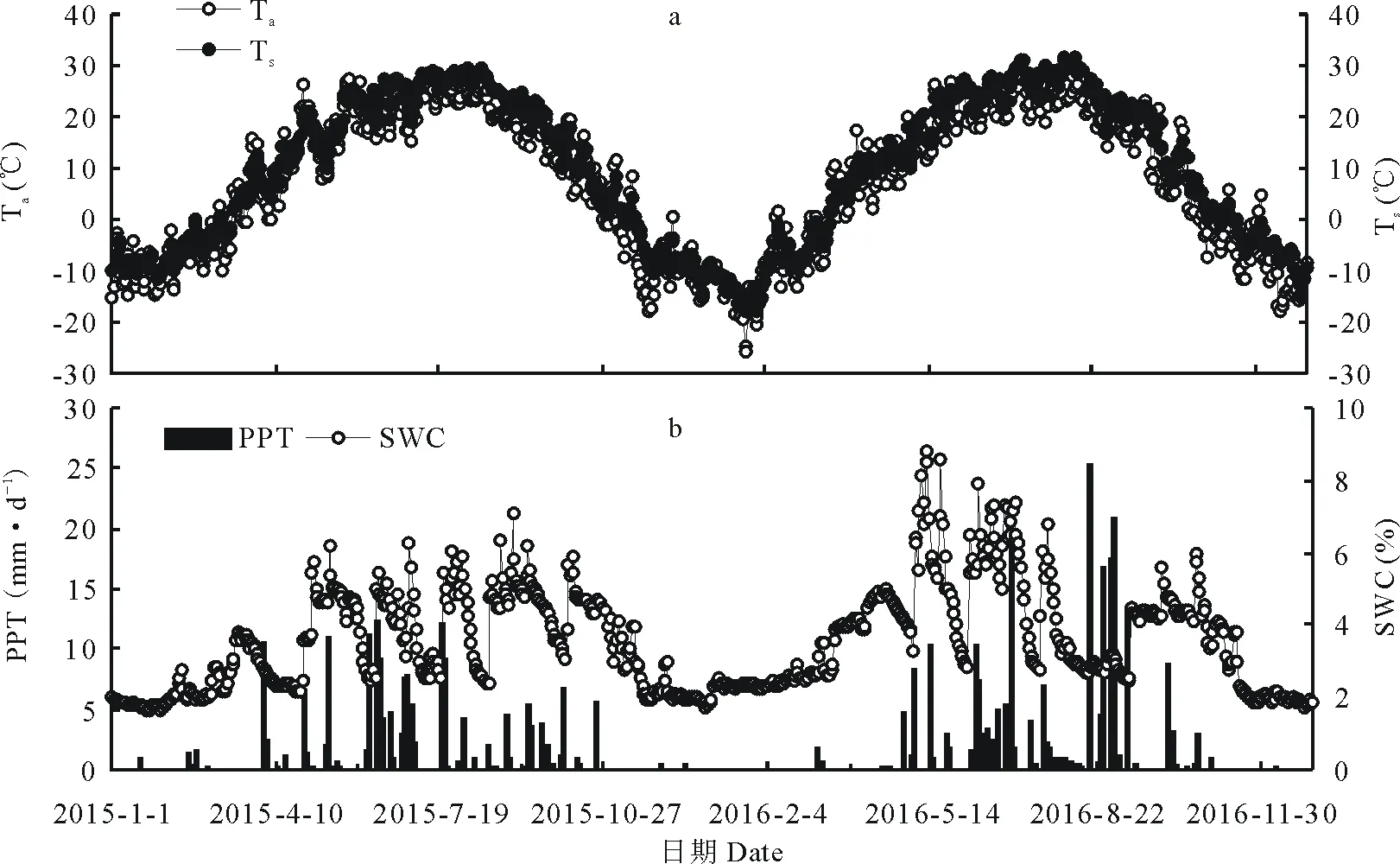
圖1 空氣溫度(Ta)、5 cm土壤溫度(Ts)、5 cm土壤體積含水量(SWC)和日降水量(PPT)的季節變化Fig.1 Seasonal variation of air temperature (Ta), soil temperature at 5 cm depth (Ts), soil water content at 5 cm depth (SWC) and precipitation (PPT) in sandy grassland ecosystem
2.2 沙質草地NEE日變化
2015和2016年,沙質草地生態系統最大日均CO2吸收速率分別為-6.68(6月20日)和-9.58 g·m-2·d-1(7月14日),最大日均CO2釋放速率分別為5.69(9月19日)和5.21 g·m-2·d-1(9月18日),年日平均NEE分別為0.30和0.20 g·m-2·d-1(圖2)。
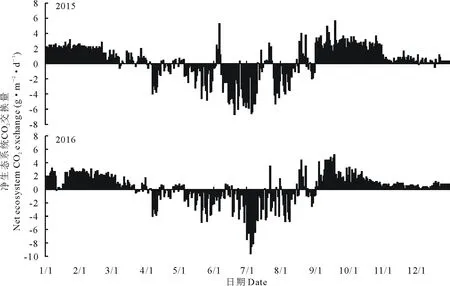
圖2 2015和2016年生態系統凈CO2交換量(NEE)逐日變化Fig.2 Variation of diurnal net ecosystem CO2 exchange (NEE) during 2015 and 2016
2.3 沙質草地NEE月動態
從不同年份逐月總量來看(圖3),2015和2016年生長季(5-9月)沙質草地生態系統均表現為凈碳吸收,吸收總量分別為120.54和139.83 g·m-2;最大月分別在6和7月,凈吸收量為74.96和103.58 g·m-2。非生長季均表現為碳排放,排放總量分別為230.33和212.82 g·m-2;最大月均出現在9月,排放量分別為88.36和78.13 g·m-2。從年際尺度凈CO2通量來看,沙質草地生態系統兩年均表現為大氣CO2的源,2015年碳釋放量(109.79 g·m-2·年-1)高于2016年(72.99 g·m-2·年-1)。
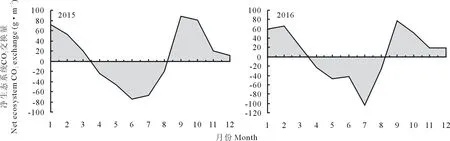
圖3 2015和2016年生態系統凈CO2交換量(NEE)月變化Fig.3 Variation of monthly net ecosystem CO2 exchange (NEE) during 2015 and 2016
2.4 NEE與環境因子之間的關系
以生長季和非生長季為周期,分年度對NEE與環境因子之間的關系進行相關分析,結果表明NEE與環境因子(Ta、Ts和SWC)之間總體上存在負相關關系(表2),除2015年非生長季的Ta和2016年非生長季的SWC與NEE之間的相關性不顯著外(P>0.05),其余時段NEE與環境因子均表現出顯著的負相關關系,說明隨著溫度和濕度的增加,生態系統固定CO2的能力增加。

表2 NEE與環境因子之間的皮爾遜相關系數Table 2 The Pearson’s correlation coefficients between NEE and environment factors
注:**表示相關系數通過0.01顯著性檢驗;*表示通過0.05顯著性檢驗。Ta:空氣溫度;Ts:5 cm土壤溫度; SWC:5 cm土壤體積含水量。
Note: **The correlation coefficient was tested by 0.01 significance test;*The correlation coefficient was tested by 0.05 significance test. Ta: Air temperature;Ts: Soil temperature at 5 cm depth; SWC: Soil water content at 5 cm depth.
3 討論
本研究中沙質草地生態系統2015和2016年碳吸收速率的日均最大值分別為6.68和9.58 g·m-2·d-1,與西藏當雄草原化嵩草(Kobresiamyosuroides)草甸(7.43 g·m-2·d-1)[16]和蒙古國干旱草原(8.44 g·m-2·d-1)[17]相當,稍低于三江源退化高寒草地(11.01 g·m-2·d-1)、三江源人工草地(10.64 g·m-2·d-1)[6]和海北高寒草甸(11.74 g·m-2·d-1)[18],但遠低于加拿大濕草地(18.35 g·m-2·d-1)[19]和美國的奧克拉馬高原草原(36.70 g·m-2·d-1)[20]。
在日尺度上,兩個生長季內生態系統均出現了NEE為正值(即碳排放)的情形(圖2)。對應圖1b可知,NEE為正的日期均出現在降水發生后,生長季發生于半干旱地區的陣發式降水能夠強烈地激發土壤中CO2的釋放量[21-23],導致生態系統CO2的日排放量超過固定量,因而NEE表現為正值。而在非生長季NEE出現負值(即碳吸收)的情形,其原因可能是冬季環境溫度低,土壤呼吸微弱,但是CO2在水中的溶解度隨溫度下降而升高,此時土壤孔隙中的CO2易被土壤水分吸收而積聚在凍土層中,大氣中的CO2濃度因高于土壤孔隙中的CO2濃度而向土壤擴散[24],表現為碳吸收。
本研究中沙質草地生態系統生長季凈碳吸收量(2015和2016年分別為120.54和139.83 g·m-2)低于以往類似的研究結果,例如內蒙古錫林浩特羊草草原為183.50~249.56 g·m-2[25]、海北高寒灌叢為500.00~611.00 g·m-2[1]、北美大草原為279.00~345.00 g·m-2[18]。同樣,本研究中兩個非生長季日平均CO2凈釋放速率(分別為1.62和1.47 g·m-2·d-1)相對較低,例如海北高寒灌叢為1.82~2.05 g·m-2·d-1[1]、美國堪薩斯州的高桿草大草原為3.00 g·m-2·d-1[7]。
生態系統碳通量受生物、非生物等諸多因子的影響,即使是同一個生態系統不同年際間碳收支也存在很大的波動。例如三江源退化高寒草甸在相鄰年際間分別表現為弱碳源和弱碳匯[6];吉林通榆退化草原也出現了類似的現象,該生態系統出現碳釋放的年份降水異常偏少,日均凈碳交換速率較低且生長季凈碳吸收過程較短[25];內蒙古典型草原在濕潤年呈弱碳匯,而在干旱年迅速轉變為碳源[26]。水分的虧缺一方面會嚴重的抑制草地生態系統的碳吸收能力;另一方面影響著植物的生長發育,在降水減少情況下,植物的光合速率和蒸騰速率會下降[6],而且干旱可以導致植物生長期縮短,從而引起系統向碳源方向轉變[20,27]。本研究中連續兩年的觀測表明在年際尺度上沙質草地生態系統均表現為碳源,其根本原因在于降水偏少,但要充分認識該類型生態系統碳源/匯功能特征變化對于降水的響應規律及機制,需要更長時間序列上的連續觀測。
4 結論
沙質草地生態系統日均碳通量變化幅度較小,2015和2016年最大CO2吸收速率分別為-6.68和-9.58 g·m-2·d-1,最大日CO2釋放速率分別為5.69和5.21 g·m-2·d-1。
沙質草地生態系統在相對干旱的年份表現為碳源,2015和2016年凈碳釋放量分別為109.79和72.99 g·m-2·年-1。
總體而言,NEE與空氣溫度、土壤溫度及土壤含水量呈現負相關關系,即隨著環境溫度和水分的增加,生態系統固定CO2的能力增強。
References:
[1] Qiao C L, Li J M, Wang J H,etal. Annual carbon dioxide flux variation of alpine shrub ecosystem in the Qinghai-Tibet plateau. Acta Agrestia Sinica, 2011, 19(6): 910-916.
喬春連, 李婧梅, 王基恒, 等. 青藏高原高寒灌叢生態系統CO2通量年變化特征研究. 草地學報, 2011, 19(6): 910-916.
[2] Dugas W A, Heuer M L, Mayeux H S. Carbon dioxide fluxes over bermudagrass, native prairie, and sorghum. Agricultural and Forest Meteorology, 1999, 93(2): 121-139.
[3] Suyker A E, Verma S B. Year-round observations of the net ecosystem exchange of carbon dioxide in a native tallgrass prairie. Global Change Biology, 2001, 7(3): 279-289.
[4] Valentini R, Gamon J A, Field C B. Ecosystem gas exchange in a California grassland: Seasonal patterns and implications for scaling. Ecology, 1995, 76(6): 1940-1952.
[5] Jmo S, Hall D O. The global carbon sink: a grassland perspective. Global Change Biology, 1998, 4(2): 229-233.
[6] Wang B. Study on the Carbon Flux and Its Controlling Mechanism of Degraded Alpine Meadow and Artificial Pasture in the Three-Rive Source Region of the Qinghai-Tibet Plateau. Tianjin: Nankai University, 2014.
王斌. 三江源區退化和人工草地生態系統CO2通量及其影響機制的研究. 天津: 南開大學, 2014.
[7] Kim J, Verma S B, Clement R J. Carbon dioxide budget in a temperate grassland ecosystem. Geophysical Research, 1992, 97(D5): 6057-6063.
[8] Chen Z Z, Wang S P. Typical Grassland Ecosystem in China. Beijing: Science Press, 2000.
陳佐忠, 汪詩平. 中國典型草原生態系統. 北京: 科學出版社, 2000.
[9] Balogh J, Czóbel S, Juhász A,etal. Seasonal carbon-balance of a semi-desert temperate grassland ecosystem over a year period. Acta Biologica Szegediensis, 2002, 46(3): 221-222.
[10] Peng Q, Qi Y C, Dong Y S,etal. Responses of carbon cycling key processes to precipitation changes in arid and semiarid grassland ecosystems: a review. Progress in Geography, 2012, 31(11): 1510-1518.
彭琴, 齊玉春, 董云社, 等. 干旱半干旱地區草地碳循環關鍵過程對降雨變化的響應. 地理科學進展, 2012, 31(11): 1510-1518.
[11] Shang W, Li Y Q, Wang S K,etal. Dynamic changes of surface soil organic carbon and light-fraction organic carbon after mobile dune afforestation with Mongolian pine in Horqin Sandy Land. Chinese Journal of Applied Ecology, 2011, 22(8): 2069-2074.
尚雯, 李玉強, 王少昆, 等. 科爾沁沙地流動沙丘造林后表層土壤有機碳和輕組有機碳的變化. 應用生態學報, 2011, 22(8): 2069-2074.
[12] Zhao H L, Li Y Q, Zhou R L. Effects of desertification on C and N storage in grassland ecosystem on Horqin Sandy Land. Chinese Journal of Applied Ecology, 2007, 18(11): 2412-2417.
趙哈林, 李玉強, 周瑞蓮. 沙漠化對科爾沁沙質草地生態系統碳氮儲量的影響. 應用生態學報, 2007, 18(11): 2412-2417.
[13] Liu X P, He Y H, Zhao X Y,etal. Characteristics of precipitation in Nanman region of Horqin sandy land. Research of Soil and Water Conservation, 2011, 18(2): 155-158.
劉新平, 何玉惠, 趙學勇, 等. 科爾沁沙地奈曼地區降水變化特征分析. 水土保持研究, 2011, 18(2): 155-158.
[14] Zhu Z L, Sun X M, Zhang R H,etal. The estimate of energy and mass exchange in Inner Mongolia semi-arid grassland micrometeorological methods. Climatic and Environmental Research, 2002, 7(3): 351-358.
朱治林, 孫曉敏, 張仁華, 等. 內蒙古半干旱草原能量物質交換的微氣象方法估算. 氣候與環境研究, 2002, 7(3): 351-358.
[15] Falge E, Baldocchi D, Olson R,etal. Gap filling strategies for long term energy flux data sets. Agricultural and Forest Meteorology, 2001, 107(1): 71-77.
[16] Kato T, Tang Y, Gu S,etal. Temperature and biomass influences on interannual changes in CO2exchange in an alpine meadow on the Qinghai-Tibetan Plateau. Global Change Biology, 2006, 12(7): 1285-1298.
[17] Li S G, Asanuma J, Eugster W,etal. Net ecosystem carbon dioxide exchange over grazed steppe in central Mongolia. Global Change Biology, 2005, 11(11): 1941-1955.
[18] Shi P L, Sun X M, Xu L L,etal. The net carbon flux and its influencing factors in Tibetan alpine steppe meadow ecosystem. Science in China: Earth Science, 2006, 36(S1): 194-203.
石培禮, 孫曉敏, 徐玲玲, 等. 西藏高原草原化嵩草草甸生態系統CO2凈交換及其影響因子. 中國科學: 地球科學, 2006, 36(S1): 194-203.
[19] Frank A B. Carbon dioxide fluxes over a grazed prairie and seeded pasture in the Northern Great Plains. Environmental Pollution, 2002, 116(3): 397-403.
[20] Suyker A E, Verma S B, Burba G G. Interannual variability in net CO2exchange of a native tallgrass prairie. Global Change Biology, 2003, 9(2): 255-265.
[21] Saetre P, Stark J M. Microbial dynamic sand carbon and nitrogen cycling following rewetting of soils beneath two semi-arid plant species. Oecologia, 2005, 142: 247-260.
[22] Wang X Y, Li Y L, Zhao X Y,etal. Responses of soil respiration to different environment factors in semi-arid and arid areas. Acta Ecologica Sinica, 2012, 32(15): 4890-4901.
王新源, 李玉霖, 趙學勇, 等. 干旱半干旱區不同環境因素對土壤呼吸影響研究進展. 生態學報, 2012, 32(15): 4890-4901.
[23] Hunt J E, Kelliher F M, Mcseveny T M,etal. Evaporation and carbon dioxide exchange between the atmosphere and a tussock grassland during a summer drought. Agricultural & Forest Meteorology, 2002, 111(1): 65-82.
[24] Zhang J X, Cao G M, Zhou D W,etal. The carbon storage and carbon cycle among the atmosphere, soil, vegetation and animal in theKobresiahumilisalpine meadow ecosystem. Acta Ecologica Sinica, 2003, 23(4): 627-634.
張金霞, 曹廣民, 周黨衛, 等. 高寒矮嵩草草甸大氣-土壤-植被-動物系統碳素儲量及碳素循環. 生態學報, 2003, 23(4): 627-634.
[25] Du Q, Liu H Z, Feng J W,etal. Carbon dioxide exchange processes over the grassland ecosystems in semiarid areas of China. Science in China: Earth Science, 2012, 55(4): 644-655.
杜群, 劉輝志, 馮健武, 等. 半干旱區草原生態系統的碳交換特征. 中國科學: 地球科學, 2012, 55(4): 644-655.
[26] Hao Y, Wang Y, Sun X,etal. Seasonal variation in carbon exchange and its ecological analysis overLeymuschinensissteppe in Inner Mongolia. Science in China: Earth Science, 2006, 49(Supple 2): 186-195.
[27] Meyers T P. A comparison of summertime water and CO2fluxes over rangeland for well watered and drought conditions. Agricultural & Forest Meteorology, 2001, 106(3): 205-214.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
