科爾沁沙地草地植物群落功能性狀對封育和放牧的響應
2018-01-09 08:35:44左小安岳喜元劉良旭何釗全
農業工程學報 2017年24期
張 晶,左小安,楊 陽,岳喜元,張 婧,呂 朋,周 欣,連 杰,劉良旭,何釗全
?
科爾沁沙地草地植物群落功能性狀對封育和放牧的響應
張 晶1,2,3,左小安1,2※,楊 陽1,岳喜元1,張 婧1,呂 朋1,周 欣2,連 杰2,劉良旭1,何釗全2
(1. 中國科學院西北生態環境資源研究院烏拉特荒漠草原站,蘭州 730000; 2. 中國科學院西北生態環境資源研究院奈曼沙漠化研究站,蘭州 730000;3. 中國科學院大學,北京 100049)
該研究以科爾沁沙地中草甸(meadow)、針茅草原(steppe)、疏林草地(scattered tree grassland)和沙質草地(sandy grassland)4種草地的18種優勢植物為研究對象,分別測定了不同草地植物群落在封育和放牧下的高度(height)、比葉面積(specific leaf area,SLA)、葉片干物質含量(leaf dry matter content,LDMC)、葉片面積(leaf area,LA)、葉片氮含量(leaf nitrogen content,LNC)和葉片碳同位素(leaf carbon isotopes)6個相關于植物生理生態過程的功能性狀指標。結果表明:1)長期放牧增加了草地中一年生植物和C4植物的豐富度和重要值,降低了沙質草地中多年生植物的相對生物量;2)沙質草地植物群落的SLA和δ13C高于其他3種草地,LDMC小于其他3種草地;3)長期放牧降低了草地植物群落的高度和LA,增加了草地植物群落的δ13C,SLA、LDMC和LNC則不受放牧的影響;4)植物功能性狀中,SLA與LDMC極顯著負相關且不受放牧的干擾;而長期放牧導致了高度與SLA和δ13C顯著負相關以及SLA和δ13C顯著正相關。科爾沁沙地長期放牧的草地中一年生植物和C4植物比例相對增加,且優勢植物具有較低的高度和LA以及較高的δ13C,表明了草地植物群落通過物種競爭結構和功能性狀權衡策略的改變來適應長期放牧。因此,減輕放牧壓力,改善植物群落的功能性狀,對于該區域退化草地的恢復及持續管理具有一定的重要意義。
植物;生長;生理學;草地;放牧;草地植物群落;封育;不同草地類型
0 引 言
草地是陸地上分布最廣的生態系統之一,中國天然草地總面積約為4.0×108hm2,約占國土總面積的41.7%[1]。草地生態系統的植物群落結構及生態功能不僅取決于氣候和土壤變化等因素,并且受到土地利用變化以及人類活動的影響[2]。放牧是草地利用的主要方式之一,主要是通過家畜的采食與踐踏影響土壤的物理結構[3],同時家畜對營養物質的攝取轉化及排泄物歸還等影響草地營養物質的循環,導致草地土壤化學成分的變化,從而進一步影響植物的生長[4-5]。放牧不僅影響著草地植物的葉片、根系和種子庫等植被特征,而且也對土壤容重、土壤孔隙結構及土壤肥力(碳、氮含量、氮的礦化、微生物活性)等也具有重要的的影響[6-8]。長期超載放牧,使草地植被覆蓋率和生產力急劇降低、土壤貧瘠化,最終草畜平衡被打破,草地逐漸開始退化。植物功能性狀被看成是植物適應環境變化與干擾的不同反映,是植物與環境相互作用的結果[9]。已有研究表明,植物和土壤對放牧敏感性的順序為:地上植株>根系>土壤[10]。葉片性狀作為植物與外界環境直接接觸的器官,可以最大限度地提供有關植物生長和適應環境的重要信息,反映植物適應環境的生存對策[11-13]。在群落水平上,由于食草動物采食可選擇范圍變寬,而且不同植物種對放牧干擾的響應也不一致,因而放牧對植物群落組成的影響遠比對單一植物種群內植株個體的影響復雜[14]。因此,基于生態系統中物種相對生物量結合相關功能性狀的研究,可以很好的將個體水平放大到群落水平,凸顯放牧對于草地植物群落的影響,為中國北方草地的管理提供一定的科學依據。
植物功能性狀是植物長期適應外部環境過程中經遺傳變異和自然選擇而形成的穩定指標,可以較好地反映植物生長狀況、資源利用策略和植物對環境變化的響應及適應[15-16]。植物高度(height)會影響光來源、熱量負荷、濕度和種子擴散[17];較高的植物個體,表明植物生長速率較快、生物量較大[18-20]。葉面積(LA)能很好地反映植物對周圍環境的適應特征,是植物比較生態學研究中的重要指標[21-24]。比葉面積(SLA)、葉片干物質含量(LDMC)以及葉片氮含量(LNC),均與植物的結構和組織特性密切相關,極大地影響著植物的生長[25-27]。高SLA的植物,往往光合速率高,有著較強的碳貯存能力和氮素利用效率[28];高的LDMC植物,具有較強的忍耐性和抗壓能力[29],在養分限制區,植物往往采用高LDMC、低SLA的資源保守策略,并通過存儲充足的氮來完成生活史[25,30-31]。植物葉片碳同位素(δ13C)反映了與植物光合、蒸騰強度相關聯的水分利用率,指示著植物的長期水分利用效率[32];葉片的δ13C與氮含量的變化對于植物適應干旱環境具有非常重要的作用[33]。因此,研究不同草地植物功能性狀對封育和放牧的響應,有助于揭示不同草地植物群落的構建及其對環境變化的適應機理,對于退化草地植被恢復和管理具有重要的理論及實踐意義。
科爾沁沙地地處中國溫帶半干旱草原區,由于過度的開墾、放牧和持續的干旱,導致了該區域生物資源銳減和土地生產力下降[34]。近些年,隨著退耕還草和封育禁牧等政策的實施,沙漠化蔓延速度減緩,退化植被得以恢復,沙化土地質量有所改善[35]。有關封育和放牧對該區域沙丘生境影響的研究報道較多,主要集中在對沙丘土壤呼吸、植被特征以及物種豐富度等方面[36-37]。然而,由于人類活動和氣候變化的影響,該區域不僅存在各類型沙丘,也發育了以多年生禾本科植物為主的濕草甸、針茅草原、疏林草地以及一年生植物為主的沙質草地[38]。本文以上述4種草地的植物為研究對象,探討長期封育和放牧對草地植物重要值、生物量以及植物群落6項功能性狀(植物高度、比葉面積、葉片干物質含量、葉面積、葉片氮含量、葉片碳同位素)指標的影響,以期揭示:1)封育和放牧對不同草地植物重要值和相對生物量的影響;2)不同植物群落功能性狀對草地類型的差異化響應;3)不同草地植物群落功能性狀對封育和放牧差異化的響應;4)草地植物群落功能性狀差異的驅動機制,為進一步加強對草地的管理、遏制草地的退化和促進草地生態系統可持續發展提供一定的理論支撐。
1 材料與方法
1.1 研究區概況
研究區位于科爾沁沙地42°55′-44°41′ N,120°42′-121°33′E之間,海拔200~360 m。屬溫帶半干旱大陸性氣候,夏季炎熱,冬季寒冷;年均溫3~7 ℃,≥10 ℃年積溫2 200~3 200 ℃;年均降水量350.0 mm,80%的降水集中在6-9月。草地類型主要有草甸(M)、針茅草原(SS)、疏林草地(SG)和沙質草地(G)。主要植物種有針茅()羊草()糙隱子草()賴草()、蘆葦()、興安胡枝子()虎尾草()狗尾草()、毛馬唐()等。其中草甸主要以灰色草甸土為主,針茅草原和疏林草地主要以典型栗鈣土為主,沙質草地主要以沙質栗鈣土為主。
1.2 研究方法
1.2.1 取樣方法
2014年8月,在該研究區域內選取長期保護封育和放牧下的草甸(M)、針茅草原(SS)、疏林草地(SG)和沙質草地(G)4種草地為研究對象。4種封育草地圍欄保護大約20~30 a,冬季偶有放牧或割草,干擾程度較輕;放牧草地均為封育草地附近無圍欄保護的長期公共自由放牧區,放牧強度大約每公頃2頭牛、6只羊,屬于重度放牧[2,39]。在每個封育草地中設置6個面積為20 m× 20 m的典型樣地,樣地間距最小為500 m。每個樣地隨機設置3個1 m×1 m樣方,調查樣方中優勢植物種的生物量。
1.2.2 試驗方法
參照植物功能性狀取樣和測定標準方法[40-42]對樣方內所有的物種進行葉功能性狀的取樣和測定,植物功能性狀包括:height(植物高度),cm;SLA(比葉面積),m2/kg;LDMC(葉片干物質含量),g/kg;LA(葉面積),cm2;LNC(葉片N含量)、碳同位素(δ13C)等。
每個樣方內植物取5~10株,選擇10片無病蟲害的完全伸展葉,用剪刀剪下,置于濕潤的濾紙之間,迅速放入密封袋貯藏于便攜式車載冰箱內(內部溫度<5 ℃)。回到實驗室將葉片去葉柄后浸入水中,置于黑暗環境中(5 ℃)冷藏12 h。取出后迅速吸干葉表水分,在百萬分之一的電子天平上稱得葉鮮質量。然后將葉片平鋪在掃描儀上掃描,再用ImageJ1.48v軟件進行處理,得到葉面積(LA)。最后,將掃描后的葉片放入60 ℃烘箱內烘48 h,取出稱得葉干質量。LDMC=葉干質量/葉鮮質量,SLA=葉面積/葉干質量。
為保證每種植物有足夠的葉片用以測定樣品N和δ13C,在每個樣方附近獲取部分植物的葉,帶回實驗室烘干稱質量后,與原來每個樣方烘干稱質量后的樣品混合,制成分析植物葉片N和δ13C的樣品。再將每個植物葉的樣品混合粉碎,過0.25 mm篩,存儲備用。分析前在65 ℃的干燥箱中烘干1 h,放入干燥器中保存待測。采用元素分析儀(Costech ECS 4010,意大利)測定樣方中每種植物葉的N和δ13C含量。
1.3 數據分析
1.3.1 物種相對生物量(P)
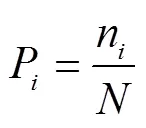
式中N是物種在群落內的生物量,是群落中全部物種的生物量。
1.3.2 植物群落功能性狀(CWM)
植物群落功能性狀(CWM),即就某一性狀而言,群落內每一物種該性狀的值與該物種相對生物量積的總和[43],表示群落內某功能性狀的均值,常用來評估群落動態和生態系統特征[43]。CWM由下列公式計算[44]

式中是物種在群落內的相對生物量,trait是物種的性狀值。
1.3.3 物種重要值
根據植物群落的數量特征,計算每種類型草地每個物種的重要值,公式如下[45]
物種重要值=(相對高度+相對蓋度+相對生物量)/3
1.4 統計分析
采用SPSS 17.0進行數據分析,單因素方差分析比較封育和放牧下不同草地CWM的差異;雙因素方差分析比較放牧和草地類型對CWM的影響,多重比較采用最小顯著性差異法(LSD)。
2 結果與分析
2.1 放牧對不同草地優勢物種重要值和相對生物量的影響
表1表明,4種草地在封育和放牧下,其優勢植物多樣性和豐富度都有顯著的差異。在封育下,草甸中優勢植物為多年生的賴草、蘆葦等,重要值為22.71、12.35;針茅草原優勢植物為多年生的針茅和羊草等,重要值為44.22、14.86;疏林草地優勢植物為多年生的糙隱子草、一二年生的狗尾草、黃蒿以及半灌木興安胡枝子等,重要值依次為31.50、13.03、10.31、12.42;沙質草地優勢植物為一二年生的狗尾草、黃蒿和多年生的糙隱子草、蘆葦等,重要值依次為22.30、9.79、12.39、9.72。而在長期放牧干擾下,草甸中優勢植物為一年生的虎尾草和多年生的賴草、蘆葦及皺果苔草等,重要值依次為16.58、14.17、13.18、9.72;針茅草原優勢植物為半灌木興安胡枝子、多年生的糙隱子草和針茅等,重要值為28.68、25.84、12.01;疏林草地優勢植物為一二年生的虎尾草和毛馬唐等,重要值為29.04、11.90;沙質草地優勢植物為一年生的馬唐、虎尾草、黃蒿和畫眉草等,重要值為16.63、13.59、11.34、10.23。由此表明,4種草地在長期封育下,草地植物群落的優勢植物主要為多年生禾本科植物,但在長期放牧影響下,草地植物群落中一二年生植物和灌木的優勢度有所增加。此外,C4植物的多樣性和重要值有所增加,尤其在疏林草地和沙質草地中有較大幅度的增加。這表明,放牧明顯導致植物種群內個體組成變化,改變了群落的競爭結構。

表1 不同草地優勢物種重要值
注:M:草甸;SS:針茅草原;SG,疏林草地;G:沙質草地;AS:一二年生植物;PS:多年生植物;C3:C3植物;C4:C4植物。
Note: M: Meadow; SS:steppe; SG:Scattered tree grassland; G: Sandy grassland; AS: Annual or two species; PS: Perennial species; C3: C3species; C4:C4species.
封育和放牧下草甸、疏林草地和沙質草地一年生植物的相對生物量無顯著差異(圖1a),放牧干擾下,4種草地多年生植物相對生物量發生了顯著的變化,沙質草地中多年生植物的相對生物量在放牧干擾下顯著降低(<0.05)(圖1b)。
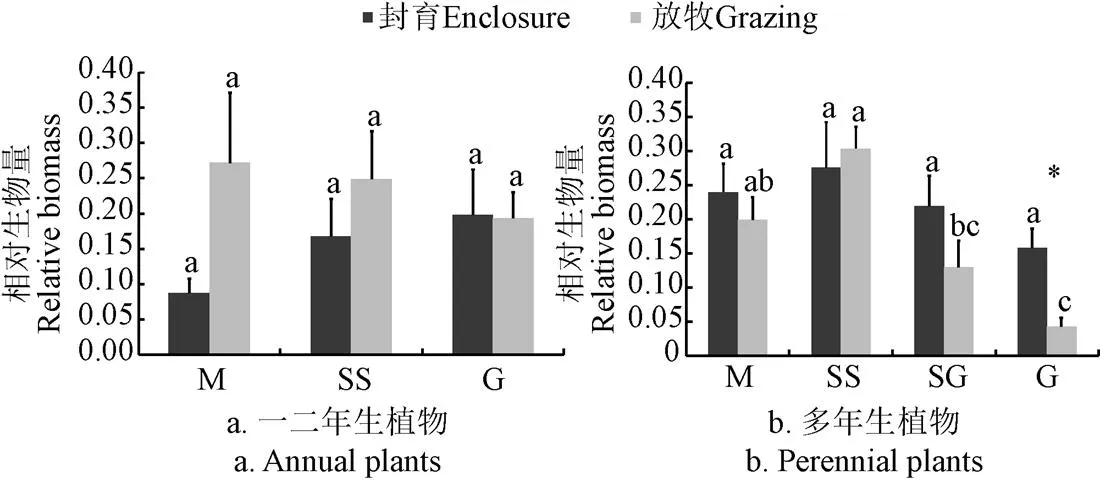
注:不同字母表示同一管理下不同草地植物相對生物量在P<0.05水平上的差異;*表示不同管理下同一草地植物相對生物量在P<0.05水平上的差異。
2.2 放牧對不同草地CWM的影響
由方差分析表2看出,不同草地植物群落的height、SLA、LDMC、LA和δ13C差異極顯著(<0.01),長期放牧極顯著影響height、LA和δ13C(<0.01),二者交互作用下height、SLA、LDMC和LA差異顯著(<0.05)。
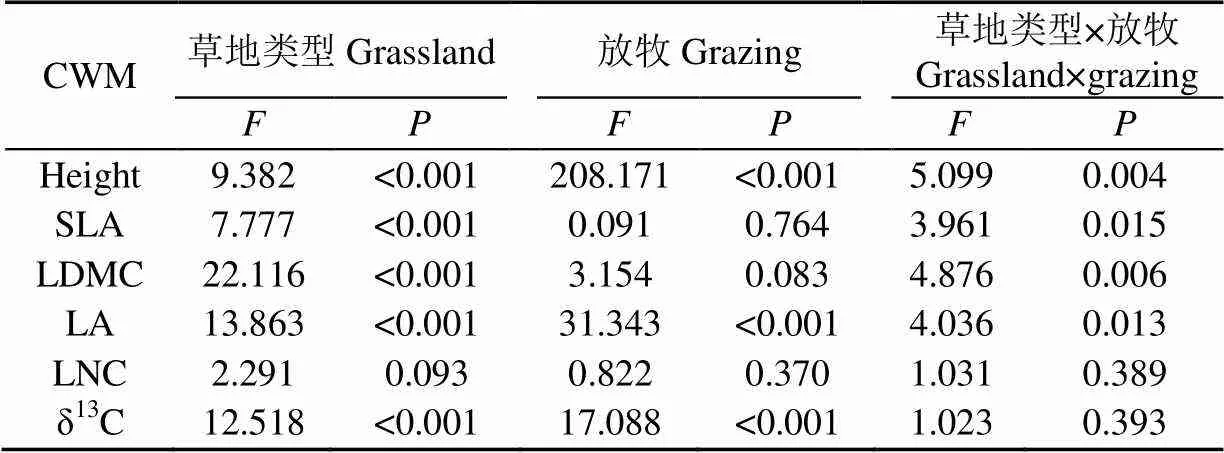
表2 植物群落功能性狀CWM方差分析表
從表3可以看出,針茅草原植物群落的height大于其他3種草地(<0.05),而長期放牧降低了草地植物群落高度(<0.05);沙質草地植物群落的SLA大于其他3種草地(<0.05),而LDMC小于其他3種草地(<0.05),放牧對于草地植物群落SLA和LDMC無顯著影響;草甸植物群落的LA大于其他3種草地,放牧減小了草地植物群落的LA;針茅草原植物群落的LNC大于草甸,而放牧對于草地植物群落的LNC無顯著影響;沙質草地植物群落δ13C高于其他3種草地(<0.05),且放牧提高了草地植物群落的δ13C(<0.01)。
從表4看出,封育和放牧下植物群落height最大的分別為針茅草原和草甸(<0.05),放牧極顯著降低了4種草地植物群落的height(<0.01)。封育下,針茅草原植物群落的SLA小于其他3種草地(<0.05),而在放牧干擾下,沙質草地植物群落的SLA大于其他3種草地(<0.05),其中,草甸植物群落的SLA顯著降低(<0.05)。封育下,針茅草原和疏林草地植物群落的LDMC大于草甸和沙質草地(<0.05),而在放牧干擾下,針茅草原植物群落的LDMC高于其他3種草地(<0.05),其中,草甸植物群落的LDMC顯著升高(<0.05)。封育下,草甸和沙質草地植物群落的LA大于針茅草原和疏林草地(<0.05),而在放牧干擾下,只有草甸植物群落的LA大于其他3種草地(<0.05),放牧顯著降低了草甸(<0.01)、針茅草原(<0.01)和沙質草地植物群落的LA(<0.05)。封育和放牧下沙質草地植物群落的δ13C大于其他3種草地(<0.05),其中,放牧極顯著提高了疏林草地植物群落的δ13C(<0.01)。

表3 草地類型和管理方式對CWM的影響
注:不同小寫字母表示不同草地CWM在<0.05水平上的差異;不同大寫字母表示不同管理方式下CWM在<0.05水平上的差異,下同。
Note: Different small letters indicate multiple comparison of plant functional traits in community level in different grasslands at 0.05 level, different capital letters indicate multiple comparison of plant functional traits in community level in different management modes at 0.05 level, the same as below.

表4 放牧對不同草地CWM的影響
注:*, **分別表示不同管理下同一草地CWM在<0.05和<0.01水平上的差異。
Note: *, ** express multiple comparison of CWM of different treatment in same grassland at 0.05 and 0.01 level.
由表5可以看出,草地CWM之間有顯著的相關關系。在封育和放牧下,植物群落SLA與LDMC極顯著負相關(<0.01)。封育下,LA與LDMC極顯著負相關(<0.01),LNC與LDMC顯著正相關(<0.05),而放牧消除了以上的相關關系。在放牧干擾下,SLA與height、δ13C與height極顯著負相關(<0.01),δ13C與SLA極顯著正相關(<0.01)。
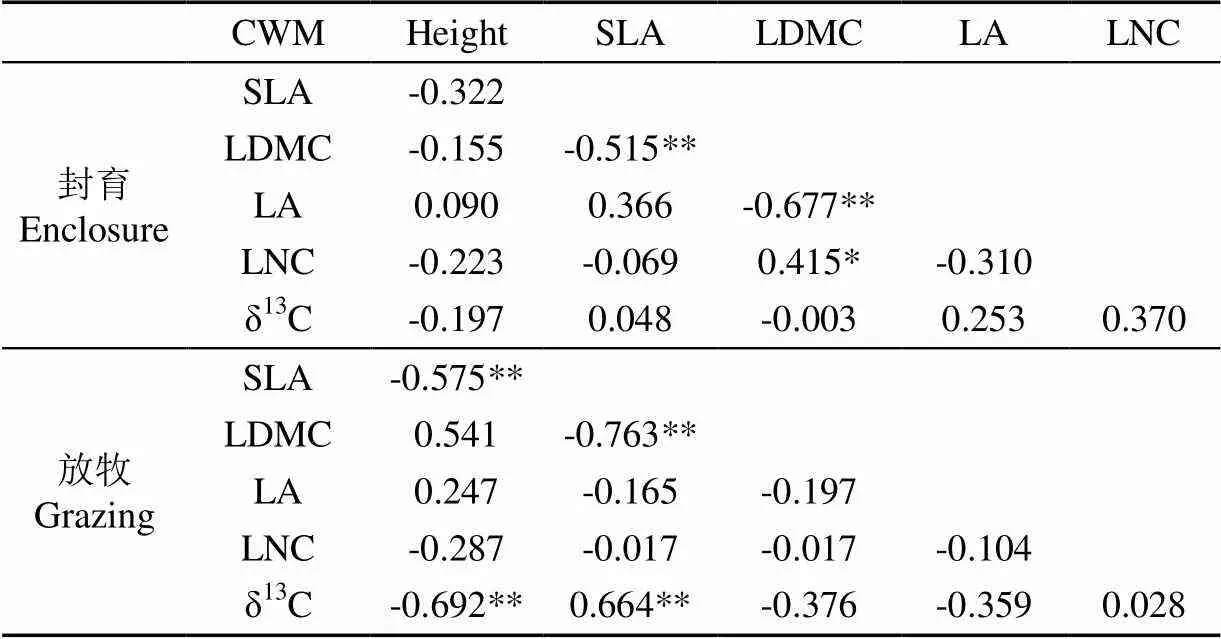
表5 草地CWM相關關系
3 討 論
科爾沁沙地生態環境演化過程中,優勢植物通過不斷地權衡資源的分配和調節生理生態過程,從而在植物個體大小和葉片形態上形成了能夠適應不同環境的植物功能性狀[38,46]。我們的研究結果表明,不同草地植物群落功能性狀之間有著明顯的差異,說明了草地植物功能性狀對生境變化發生了協同性響應,這與Zheng等的研究結果相一致[47]。科爾沁沙地中不同草地及其生境間的土壤特性存在較大的差異[48],導致了植物群落功能性狀有所不同。針茅草原中優勢物種為“針形”葉的多年生禾本科的大針茅,因而其植物群落高度較高、葉面積較小。沙質草地生產力低、土壤貧瘠,其優勢植物大多為一二年生C4植物,較多年生C3植物而言,一二年生C4植物具有較強的光合生產能力和資源利用效率[49],其高SLA低LDMC的特點有助于利用有限的資源迅速完成生活史[50]。草甸中水肥條件相對良好,植物生長過程中受到的水分和養分限制較小,因而植物群落葉面積相對較大。在水分有限的條件下,植物為了生長和生存,從而提高了植物的水分利用效率[51-52]。沙質草地土壤含水量相對較低[48],植物群落中占優勢的一二年生植物為了快速生長和繁殖,從而提高了葉片的δ13C值來適應相對干旱的土壤環境。
已有研究表明,長期放牧極大地改變著草地植物的物種組成和生活型變化[48]。本研究中,草甸、疏林草地和沙質草地在放牧干擾下出現了虎尾草、馬唐和狗尾草等一二年生C4植物,說明了科爾沁沙地中長期的放牧明顯地增加了草地生態系統中一二年生植物的優勢,這與前人的研究結果相一致[7,53]。長期的放牧也極大地影響草地植物的功能性狀[14],而且不同草地植物群落功能性狀對放牧的響應有所不同[47]。草原植物矮化是長期過度放牧下植株變矮、葉片變短變窄、節間縮短等性狀的集合[54]。在本研究中,不同草地植物群落在長期放牧下植物高度和葉面積顯著降低,支持了草地植物應對踐踏和啃食而產生的防御性響應[14]。長期放牧也導致了土壤保水和持水能力降低,從而提高草地植物群落的δ13C值。
此外,Cruz等的研究發現,15 a放牧處理后草地中以C4植物為優勢,這些植物群落通過低的葉組織密度、高的比葉面積、高的光和效率,快速增長來恢復損失的組織,增強地上部的生長能力,從而使得植物具有更強的耐牧性[55],表現出植物對放牧干擾的“快速資源獲得型策略”;也有研究表明,放牧導致了植物葉片具有低的SLA、葉片氮含量和高的LDMC,從而使得植物具有低的光合能力和養分周轉速率,表現出植物對資源利用的“保守性策略”[14]。本研究的草甸植物群落在放牧干擾下,SLA顯著降低,LDMC顯著升高,屬于“資源保持型”策略,主要原因可能是草甸中優勢物種為多年生的禾本科植物,其植物群落對于放牧的抵抗性較強,主要通過降低光合速率來應對干擾。同時,LDMC較高的葉片,木質素和纖維素成分相應較高,葉片相對較硬,適口性較差且不利于反芻動物的消化代謝[14]。本研究中草甸植物群落在放牧干擾下LDMC升高,也是草地植物“避牧”的一種生存策略。植物群落化學計量特征對于放牧的響應存在明顯的滯后性,植物群落作為一個整體需要在很長的時間尺度上做出反應[56-57],這也是本研究中葉片氮含量在草地類型和放牧下無顯著差異的主要原因。
在以往的研究中SLA與LDMC之間表現出顯著的負相關關系[23],其原因是LDMC越大,葉組織密度越高,限制了光和CO2在葉組織中的傳導[58],導致光合速率降低、SLA變小。本研究中封育和放牧下草地植物群落SLA與LDMC的極顯著負相關關系,與李玉霖等的研究結果相一致[24],說明SLA與LDMC的負相關關系沒有受到放牧的干擾。長期放牧導致了土壤風蝕和土壤的貧瘠化,草地中植物為了適應土壤環境變化而具有較低height和較高水分的生長策略[32],因而形成了草地中SLA與height、δ13C與height的負相關關系以及δ13C與SLA的正相關關系。這些結果表明了科爾沁沙地中草地植物為了適應放牧干擾過程,采取了資源利用率和水分利用率之間的權衡策略。本研究僅以一年的試驗數據揭示了不同草地植物群落功能性狀對生境變化和放牧響應的差異性,但是一年的試驗周期相對較短,其研究結果可能存在偶然性。因此,有必要進行更長時間的觀測和調查,進一步探究長期放牧和環境變化對不同草地植物功能性狀及其功能型組成的影響。
4 結 論
放牧降低了草地植物群落的高度和葉面積,降低了沙質草地中多年生植物的相對生物量,提高了草地植物群落的水分利用率。在土壤資源相對貧瘠和生產力較低的沙質草地中,植物通過高的比葉面積、低葉干物質含量的策略來充分利用有限的資源而迅速完成生活史。放牧干擾下草甸植物群落“耐牧”和“避牧”的策略為低的比葉面積和高的葉干物質含量。植物群落高度、比葉面積、葉干物質含量、葉面積、水分利用率為放牧干擾的敏感性狀,而植物群落葉片氮含量在不同草地類型和管理水平上均無顯著差異。放牧導致草地植物群落高度降低、水分利用效率增加,進而形成了比葉面積與高度、水分利用率與高度的負相關關系以及水分利用率與比葉面積的正相關關系,但比葉面積與葉干物質含量間的負相關關系不受放牧的干擾,這些結果表明了草地植物群落功能性狀在應對放牧干擾時權衡并形成了特定的生長對策。然而,研究放牧對草地植物群落功能性狀的影響,也需要充分考慮草地生態系統的初始狀況。
[1] 方精云,楊元合,馬文紅,等. 中國草地生態系統碳庫及其變化[J]. 中國科學:生命科學,2010,40(7):566-576.
Fang Jingyun, Yang Yuanhe, Ma Wenhong, et al. Ecosystem carbon stocks and their changes in China’s grasslands[J]. Sci China Life Sci, 2010, 40(7): 566-576. (in Chinese with English abstract)
[2] 趙哈林,大黑俊哉,李玉霖,等. 科爾沁沙質草地植物群落的放牧退化及其自然恢復過程[J]. 中國沙漠,2009,29(2):229-235.
Zhao Halin, Okuro Toshiya, Li Yulin, et al. Changes of plant community in grazing and restoration processes in Horqin Sand Land, Inner Mongolia[J]. Journal of Desert Research, 2009, 29(2): 229-235. (in Chinese with English abstract)
[3] Manouchehr D, Gifford G F. Influence of vegetation, rock cover, and trampling on infiltration rates and sediment production[J]. Jawra Journal of the American Water Resources Association, 1980, 16(6): 979-986.
[4] Haynes R J, Williams P H. Nutrient cycling and soil fertility in the grazed pasture ecosystem[J]. Advances in Agronomy, 1993, 49(8): 119-199.
[5] Krzic M, Broersma K, Thompson D J, et al. Soil properties and species diversity of grazed crested wheatgrass and native rangelands[J]. Journal of Range Management, 2000, 53(3): 353-358.
[6] 侯扶江,楊中藝. 放牧對草地的作用[J]. 生態學報,2006,26(1):244-264.
Hou Fujiang, Yang Zhongyi. Effets of grazing of livestock on grassland[J]. Acta Ecologica Sinica, 2006, 26(1): 244-264. (in Chinese with English abstract)
[7] 孫大帥. 不同放牧強度對青藏高原東部高寒草甸植被和土壤影響的研究[D]. 蘭州:蘭州大學,2012.
Sun Dashuai. Studies on the Effects of Grazing Intensity on Vegetation and Soil in Alpine Meadow on the Eastern Qinghai-Tibetan Plateau[D]. Lanzhou: Lanzhou University, 2012. (in Chinese with English abstract)
[8] 高英志,韓興國,汪詩平. 放牧對草原土壤的影響[J]. 生態學報,2004,24(4):790-797.
Gao Yingzhi, Han Xingguo, Wang Shiping. The effects of grazing on grassland soils[J]. Acta Ecologica Sinica, 2004, 24(4): 790-797. (in Chinese with English abstract)
[9] Zheng S X, Ren H Y, Lan Z C, et al. Effects of grazing on leaf traits and ecosystem functioning in Inner Mongolia grasslands: Scaling from species to community[J]. Biogeosciences Discussions, 2009, 6(5): 1117-1132.
[10] 李西良,侯向陽,吳新宏,等. 草甸草原羊草莖葉功能性狀對長期過度放牧的可塑性響應[J]. 植物生態學報,
2014,38(5):440-451.
Li Xiliang, Hou Xiangyang, Wu Xinhong, et al. Plastic responses of stem and leaf functional traits into long-term grazing ina meadow steppe[J]. Chinese Journal of Plant Ecology, 2014, 38(5): 440-451. (in Chinese with English abstract)
[11] Bussotti F. Functional leaf traits, plant communities and acclimation processes in relation to oxidative stress in trees: A critical overview[J]. Global Change Biology, 2008, 14(11): 2727-2739.
[12] Cornelissen J H C. Foliar nutrients in relation to growth, allocation and leaf traits in seedlings of a wide range of woody plant species and types[J]. Oecologia, 1997, 111(4): 460-469.
[13] Garnier E, Laurent G, Bellmann A, et al. Consistency of species ranking based on functional leaf traits[J]. New Phytologist, 2001, 152(1): 69-83.
[14] 趙娜,趙新全,趙亮,等. 植物功能性狀對放牧干擾的響應[J]. 生態學雜志,2016,35(7):1916-1926.
Zhao Na, Zhao Xinquan, Zhao Liang, et al. Progress in researches of response of plant functional traits to grazing disturbance[J].Chinese Journal of Ecology, 2016, 35(7): 1916-1926. (in Chinese with English abstract)
[15] Reich P B, Walters M B, Ellsworth D S. Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems[J]. Ecological Monographs, 1992, 62(3): 365-392.
[16] 劉金環,曾德慧,Don Koo LEE. 科爾沁沙地東南部地區主要植物葉片性狀及其相互關系[J]. 生態學雜志,2006,25(8):921-925.
Liu Jinhuan, Zeng Dehui, Don Koo LEE. Leaf traits and their interrelationships of main plant species in southeast Horqin sandy land[J]. Chinese Journal of Ecology, 2006, 25(8): 921-925. (in Chinese with English abstract)
[17] Givnish T J. 1-plant stems: Biomechanical adaptation for energycapture and influence on species distributions[J]. Plant Stems, 1995: 3-49.
[18] Lavorel S, Grigulis K. How fundamental plant functional trait relationships scale-up to trade-offs and synergies in ecosystem services[J]. Journal of Ecology, 2012, 100(1): 128-140.
[19] Lienin P, Kleyer M. Plant trait responses to the environment and effects on ecosystem properties[J]. Basic & Applied Ecology, 2012, 13(4): 301-311.
[20] Conti G, Díaz S. Plant functional diversity and carbon storage-an empirical test in semi-arid forest ecosystems[J]. Journal of Ecology, 2013, 101(1): 18-28.
[21] Meziane D, Shipley B. Interacting determinants of specific leaf area in 22 herbaceous species: Effects of irradiance and nutrient availability[J]. Plant Cell & Environment, 1999, 22(5): 447-459.
[22] Poorter H, Jong R D. A comparison of specific leaf area, chemical composition and leaf construction cost of field plants from 15 habitats differing in productivity[J]. New Phytologist, 1999, 143(1): 163-176.
[23] Garnier E, Shipley B, Roumet C, et al. A standardized protocol for the determination of specific leaf area and leaf dry matter content[J]. Functional Ecology, 2001, 15(5): 688-695.
[24] 李玉霖,崔建垣,蘇永中. 不同沙丘生境主要植物比葉面積和葉干物質含量的比較[J]. 生態學報,2005,25(2):304-311.
Li Yulin, Cui Jianyuan, Su Yongzhong. Specific leaf area and leaf dry matter content of some plants in different dune habitats[J]. Acta Ecologica Sinica, 2005, 25(2): 304-311. (in Chinese with English abstract)
[25] Kazakou E, Vile D, Shipley B, et al. Co‐variations in litter decomposition, leaf traits and plant growth in species from a Mediterranean old-field succession[J]. Functional Ecology, 2006, 20(1): 21-30.
[26] Cortez J, Garnier E, Pérez-Harguindeguy N, et al. Plant traits, litter quality and decomposition in a Mediterranean old-field succession[J]. Plant & Soil, 2007, 296(1/2): 19-34.
[27] Dechaine J M, Brock M T, Iniguezluy F L, et al. Quantitative trait loci × environment interactions for plant morphology vary over ontogeny in Brassica rapa[J]. New Phytologist, 2014, 201(2): 657-669.
[28] Feng Y L, Fu G L, Zheng Y L. Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation, and use efficiencies between invasive and noninvasive alien congeners[J]. Planta, 2008, 228(3): 383.
[29] Wilson P J, Thompson K, Hodgson J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies[J]. New Phytologist, 1999, 143(1): 155-162.
[30] Poorter H, Niinemets U, Poorter L, et al. Causes and consequences of, variation in leaf mass per area (LMA): A meta-analysis[J]. New Phytologist, 2009, 182(3): 565.
[31] Milcu A, Roscher C, Gessler A, et al. Functional diversity of leaf nitrogen concentrations drives grassland carbon fluxes[J]. Ecology Letters, 2014, 17(4): 435-444.
[32] Knight J D, Livingston N J, Kessel C V. Carbon isotope discrimination and water-use efficiency of six crops grown under wet and dryland conditions[J]. Plant Cell & Environment, 1994, 17(2): 173-179.
[33] 苗艷明,呂金枝,畢潤成. 不同功能型植物葉氮含量與光合特性的關系研究[J]. 植物研究,2012(4):425-429.
Miao Yanming, Lv Jinzhi, Bi Runcheng. Relationships between leaf nitrogen content and photosynthetic characteristics in different plant functional types[J]. Bulletin of Botanical Research, 2012(4): 425-429. (in Chinese with English abstract)
[34] 趙哈林,趙學勇,張銅會,等. 科爾沁沙地沙漠化過程及其恢復機理[M]. 北京:海洋出版社,2003.
[35] 王濤,吳薇,薛嫻,等. 近50年來中國北方沙漠化土地的時空變化[J]. 地理學報,2004,59(2):203-212.
Wang Tao, Wu Wei, Xue Xian, et al. Spatial-temporal changes of sandy desertified land during last 5 decades in Northern China[J]. Acta Geographica Sinica, 2004, 59(2): 203-212. (in Chinese with English abstract)
[36] 孫殿超,李玉霖,趙學勇,等. 放牧及圍封對科爾沁沙質草地土壤呼吸的影響[J]. 中國沙漠,2015,35(6):1620-1627.
Sun Dianchao, Li Yulin, Zhao Xueyong, et al. Effects of grazing and enclosure on soil respiration rate in the Horqin sandy grassland[J]. Journal of Desert Research, 2015, 35(6): 1620-1627. (in Chinese with English abstract)
[37] 呂朋,左小安,張婧,等. 放牧強度對科爾沁沙地沙質草地植被的影響[J]. 中國沙漠,2016,36(1):34-39.
Lü Peng, Zuo Xiaoan, Zhang Jing, et al. Effects of grazing intensity on vegetation in sandy grassland of Horqin[J]. Journal of Desert Research, 2016, 36(1): 34-39. (in Chinese with English abstract)
[38] Zuo Xiaoan, Zhao Xueyong, Zhao Halin, et al. Scale dependent effects of environmental factors on vegetation pattern and composition in Horqin Sandy Land, Northern China[J]. Geoderma, 2012(173/174): 1-9.
[39] 張銅會,趙哈林,大黑俊哉,等. 沙質草地連續放牧后某些土壤性質的變化[J]. 中國草地學報,2003,25(1):9-12.
Zhang Tonghui, Zhao Halin, Okuro Toshiya, et al. Variations of soil characterics after successive grazing in keerqin sandy land, Inner Mongolia[J]. Grassland of China, 2003, 25(1): 9-12. (in Chinese with English abstract)
[40] Conti G, Diaz S. Plant functional diversity and carbon storage-an empirical test in semi-arid forest ecosystems[J]. Journal of Ecology, 2013, 101(1): 18-28.
[41] Jhc C, Lavorel S, Garnier E, et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide[J]. Australian Journal of Botany, 2003, 51(4): 335-380.
[42] Spasojevic M J, Grace J B, Harrison S, et al. Functional diversity supports the physiological tolerance hypothesis for plant species richness along climatic gradients[J]. Journal of Ecology, 2014, 102(2): 447-455.
[43] Garnier E, Cortez J, Billès G, et al. Plant Functional markers capture ecosystem properties during secondary succession[J]. Ecology, 2008, 85(9): 2630-2637.
[44] Naeem S, Li S. Biodiversity enhances ecosystem reliability[J]. Nature International Weekly Journal of Science, 1997, 390(6659): 507-509.
[45] Curtis J T, Mcintosh R P. An upland forest continuum in the Prairie-Forest Border Region of Wisconsin[J]. Ecology, 1951, 32(3): 476-496.
[46] Zuo X, Zhou X, Lv P, et al. Testing associations of plant functional diversity with carbon and nitrogen storage along a restoration gradient of sandy grassland[J]. Frontiers in Plant Science, 2016, 7(e17476): 189.
[47] Zheng Shuxia, Ren Haiyan, Lan Zhichun, et al. Effects of grazing on leaf traits and ecosystem functioning in Inner Mongolia grasslands: scaling from species to community[J]. Biogeosciences, 2010, 7(3): 1117-1132.
[48] Zhang Jing, Zuo Xiaoan, Zhou Xin, et al. Long-term grazing effects on vegetation characteristics and soil properties in a semiarid grassland, northern China[J]. Environmental Monitoring & Assessment, 2017, 189(5): 216.
[49] Ripley B, Martin T. Experimental investigation of fire ecology in the C3and C4subspecies of Alloteropsissemialata[J]. Journal of Ecology, 2010, 98(5): 1196-1203.
[50] 周欣,左小安,趙學勇,等. 科爾沁沙地中南部34種植物葉功能性狀及其相互關系[J]. 中國沙漠,2015,35(6):1489-1495.
Zhou Xin, Zuo Xiaoan, Zhao Xueyong, et al. Plant functional traits and interrelationships of 34 plant species in south central Horqin Sandy Land, China[J]. Journal of Desert Research, 2015, 35(6): 1489-1495. (in Chinese with English abstract)
[51] 阮成江,李代瓊. 黃土丘陵區沙棘氣孔導度及其影響因子[J]. 西北植物學報,2001,21(6):1078-1084.
Ruan Chengjiang, Li Daiqiong. Stomatal conductance and influence factors of seabuckthorn in Loess Hilly Region[J]. Acta Botanica Boreali-Occidentalia Sinica, 2001, 21(6): 1078-1084. (in Chinese with English abstract)
[52] 孫惠玲,馬劍英,王紹明,等. 準噶爾盆地荒漠植物碳同位素組成研究[J]. 中國沙漠,2007,27(6):972-976.
Sun Huiling, Ma Jianying, Wang Shaoming, et al. The study of stable carbon isotope composition in desert plants of Junggar Basin[J]. Journal of Desert Research, 2007, 27(6): 972-976. (in Chinese with English abstract)
[53] Díaz S, Lavorel S, Mcintyre S, et al. Plant trait responses to grazing- a global synthesis[J]. Global Change Biology, 2007, 13(2): 313-341.
[54] 王煒,梁存柱,劉鐘齡,等. 草原群落退化與恢復演替中的植物個體行為分析[J]. 植物生態學報,2000,24(3):268-274.
Wang Wei, Liang Cunzhu, Liu Zhongling, et al. Analysis of the plant individual behavior during the degradation and restoring succession in steppe community[J]. Acta Phytoecologica Sinica, 2000, 24(3): 268-274. (in Chinese with English abstract)
[55] Cruz P, Quadros F L F D, Theau J P, et al. Leaf traits as functional descriptors of the intensity of continuous grazing in native grasslands in the South of Brazil[J]. Rangeland Ecology & Management, 2010, 63(3): 350-358.
[56] Coffin D P, Laycock W A, Lauenroth W K. Disturbance intensity and above-and belowground herbivory effects on long-term (14y) recovery of a semiarid grassland[J]. Plant Ecology, 1998, 139(2): 221-233.
[57] Milchunas D G. Livestock Grazing: Animal and plant biodiversity of shortgrass steppe and the relationship to ecosystem function[J]. Oikos, 1998, 83(83): 65-74.
[58] Parkhurst DF. Diffusion of CO2and other gases inside leaves[J]. New Phytologist, 1994, 126(3): 449-479.
Response of plant community functional traits in different grasslands to enclosure and grazing in Horqin Sandy Land
Zhang Jing1,2,3, Zuo Xiaoan1,2※, Yang Yang1, Yue Xiyuan1, Zhang Jing1, Lü Peng1, Zhou Xin2, Lian Jie2, Liu Liangxu1, He Zhaoquan2
(1.,,730000,; 2.,,,730000,; 3.,100049)
Functional traits as a link between plants and environment, express plants’ physiological, phenological and morphological adaptive strategy when plants face environment changes. In recent years, some trait-based means show that plant functional traits are some of the most reliable predictors of ecosystem, increasingly used to assess and predict the ecosystem feature and succession. Grazing is one of the most common utilization methods for pasture, affecting the semiarid and arid grassland ecosystems in various ways.It not only alters individual plant growth and population dynamics, but also changes soil properties, ultimately changing the vegetation composition and structure. The influence of grazing on plant communities is much greater than that of individual plant.At the community level, herbivores have extensive range of food intake, and different plant species have different response to nibbling, and therefore, it is more reasonable to explore the influence of grazing at the community level.Based on the changes of 18 dominant species in 4 different types of grassland under grazing and enclosure in Horqin sandy land,in this study, we compared the plant community functional traits in the meadow,steppe, scattered tree grassland and sandy grassland. We measured the 6 functional traits related to the process of plant growth and photosynthesis in different types of grassland under the condition of enclosure and grazing, and these traits included the plant height, specific leaf area(SLA), leaf dry matter content(LDMC), leaf area(LA), leaf nitrogen content(LNN) and leaf carbon isotopes(δ13C). The results showed that:1) Long term grazing increased the species richness and dominance of annuals and C4species, as well as decreased the relative biomass of perennials; 2) The SLA and δ13C in sandy grassland were higher than the other 3 types of grassland, while LDMC showed a reversed trend; 3) Grazing decreased the plant height and LA and increased the δ13C, while the other functional traits in different grasslands did not differ due to grazing; 4) Correlation relationship analysis showed that SLA was significantly and negatively correlated with LDMC in different types of grassland under both grazing and enclosure, suggesting that long term grazing did not change the negative associations of SLA and LDMC; however, long term grazing resulted in the significant and negative correlations between plant height, SLA and δ13C, as well as a significant and positive correlation between SLA and δ13C. In Horqin sandy land, long term grazing increased the proportion of annuals and C4species, and these species had the lower plant height and LA and the higher water use efficiency, which illustrates that the grassland community adapts to the long term grazing through the change in species competition structure and function strategy trade off. Therefore, to improve the functional traits of plant community by reducing grazing pressure is helpful and meaningful to the vegetation restoration and sustainable management in the degraded grasslands.
plants; growth; physiology; grass land; grazing;plant community traits; enclosure
10.11975/j.issn.1002-6819.2017.24.034
X17
A
1002-6819(2017)-24-0261-08
2017-07-04
2017-12-14
國家自然科學基金項目(41622103,41571106);國家重點研發計劃重點專項課題(2016YFC0500506)
張 晶,女,甘肅榆中人,博士生,主要從事植物生態方面的研究。Email:zhangjing_881014@163.com
左小安,男,博士,研究員,主要研究方向為干旱區生態格局與過程。Email:zuoxa@lzb.ac.cn
張 晶,左小安,楊 陽,岳喜元,張 婧,呂 朋,周 欣,連 杰,劉良旭,何釗全. 科爾沁沙地草地植物群落功能性狀對封育和放牧的響應[J]. 農業工程學報,2017,33(24):261-268. doi:10.11975/j.issn.1002-6819.2017.24.034 http://www.tcsae.org
Zhang Jing, Zuo Xiaoan, Yang Yang, Yue Xiyuan, Zhang Jing, Lü Peng, Zhou Xin, Lian Jie, Liu Liangxu, He Zhaoquan. Response of plant community functional traits in different grasslands to enclosure and grazing in Horqin Sandy Land[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2017, 33(24): 261-268. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2017.24.034 http://www.tcsae.org
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
中國科技論壇(2017年7期)2017-07-25 08:49:53
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年2期)2017-02-21 01:21:24
爆笑show(2016年7期)2017-02-09 09:36:13
國際漢語學報(2016年1期)2017-01-20 08:21:20
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
