游泳運動對APP/PS1轉基因小鼠學習記憶能力的影響
2017-12-12 06:14:51樊宗山敬廣偉趙海鵬
四川動物 2017年6期
樊宗山, 敬廣偉, 趙海鵬
(1. 河南農業職業學院,河南中牟451450; 2. 河南師范大學體育學院,河南新鄉453002; 3. 河南雞公山國家級自然保護區管理局,河南信陽464000; 4. 河南大學生命科學學院,河南開封475001)
游泳運動對APP/PS1轉基因小鼠學習記憶能力的影響
樊宗山1, 2, 敬廣偉3, 趙海鵬4*
(1. 河南農業職業學院,河南中牟451450; 2. 河南師范大學體育學院,河南新鄉453002; 3. 河南雞公山國家級自然保護區管理局,河南信陽464000; 4. 河南大學生命科學學院,河南開封475001)
探討游泳運動對APP/PS1轉基因小鼠學習記憶能力的影響。選擇11月齡的雄性APP/PS1轉基因小鼠,隨機平均分為對照組和游泳組,對照組常規飼養而游泳組進行1個月的游泳運動訓練。分別采用剛果紅染色、Tunel檢測、Western Blot和Morris水迷宮等實驗方法觀察小鼠大腦皮層Aβ斑形成、神經元凋亡、線粒體生成相關蛋白表達和學習記憶能力的變化情況。結果發現,游泳運動可以減少APP/PS1轉基因小鼠大腦皮層Aβ斑的形成、抑制神經元凋亡、促進線粒體生成、增強小鼠的學習記憶能力。由此可見,游泳運動可作為一項防治阿爾茨海默病的行為治療候選方案。
APP/PS1轉基因小鼠; 游泳運動; 學習記憶
阿爾茨海默病(Alzheimer’s disease,AD)是一種中樞神經退行性疾病,典型的神經病理變化是大腦內存在β樣淀粉(amyloid-β,Aβ)沉淀形成的老年斑以及神經元纖維纏結(neurofibrillary tangles),最終導致神經元丟失和神經功能障礙。神經元由于其特殊的功能決定其只能依靠氧化磷酸化提供能量,而線粒體是細胞內產生能量的主要細胞器,因此線粒體的質量體積占神經元的體積超過30%(王來,祝世功,2016)。線粒體結構破壞、活性降低、功能紊亂等所導致的突觸降解、神經元凋亡或丟失是AD發生的重要病理機制(Armand-Ugonetal.,2017;Wangetal.,2017;Yuetal.,2017)。研究表明,Aβ可富集到線粒體內,誘導線粒體分裂,促進線粒體片段化和瞬時通透孔(mitochondrial permeability transition)的開放,導致病理癥狀。抑制線粒體分裂可降低神經元的線粒體功能紊亂、突觸降解及動物的認知能力降低,促進線粒體功能則可以緩解AD的病理癥狀(Yeetal.,2015;Valasanietal.,2016;Baeketal.,2017;Reddyetal.,2017)。過氧化物酶體增殖物激活受體γ輔助活化因子1α(peroxisome proliferator-activated receptor γ coactivator 1α,PGC-1α)能促進線粒體生成,增強線粒體功能(王來,祝世功,2016),因而對維持神經元功能可能發揮重要作用。游泳運動能提高機體的認知能力,抑制疾病或衰老導致的記憶能力降低,促進神經功能恢復(崔建梅等,2013;欒海云等,2016)。本研究觀察游泳訓練的APP/PS1轉基因小鼠的學習記憶能力及其大腦神經元細胞的凋亡情況,探討游泳運動對APP/PS1轉基因小鼠學習記憶能力的影響及影響機制。
1 材料和方法
1.1動物來源
APP/PS1轉基因小鼠購自南京大學模式動物研究所并在本實驗室繁育,實驗動物生產許可證號:J004462,實驗動物使用許可證號:SCXK(蘇)2010-0001。將雄性APP/PS1轉基因小鼠雜合體(+/-)與野生型雌鼠(-/-)合籠繁殖,幼鼠出生后2周,提取小鼠DNA并做PCR鑒定仔鼠基因型。APP基因上游引物為5’-GACTGACCACTCGACCAGGTTCTG-3’,下游引物為5’-CTTGTAAGTTGGATTCTCATATCCG-3’,PCR產物為350 bp;PS1基因上游引物為5’-AATAGAGAACGGCAGGAGCA-3’,下游引物為5’-GCCATGAGGGCACTAATCAT-3’,PCR產物為608 bp。PCR反應條件為:95 ℃預變性5 min;94 ℃變性30 s,63 ℃退火45 s,72 ℃延伸1 min,35個循環。APP/PS1轉基因陽性小鼠在306 bp和608 bp處有2條條帶,野生型小鼠無條帶。APP/PS1轉基因小鼠在室溫22~25 ℃、相對濕度60%~80%的環境下飼養,以出生當天為P0,直至飼養至游泳運動結束,共12個月。
1.2試劑
Tunel細胞凋亡檢測試劑盒購自Promega公司;DAPI試劑、剛果紅染色試劑盒購自索萊寶生物科技有限公司;小鼠抗PGC-1α抗體、兔抗細胞色素c氧化酶Ⅳ(cytochrome c oxidase Ⅳ,COX Ⅳ)抗體和小鼠抗β-actin抗體購自Santa公司;辣根過氧化物標記的二抗購自北京中杉金橋生物科技有限公司;BCA蛋白定量試劑盒購自美國Life Technologies公司;ECL發光試劑盒購自Millipore公司;RIPA裂解液購自康為世紀生物科技有限公司;其他試劑均為分析純。
2 方法
2.1游泳運動訓練
雄性APP/PS1轉基因小鼠20只隨機平均分為對照組和游泳組。將游泳組小鼠放入游泳池,游泳池四周光滑且池壁高出水面20 cm,水溫30 ℃±1 ℃(等于或稍低于體溫時游泳時間最長),每次游泳15 min,每天1次,每周5 d,周二、四休息,共持續4周。如果小鼠漂浮在水中不游,用水溫計驅趕其尾部,使其不停游動。
2.2剛果紅染色Aβ斑
將對照組和游泳組小鼠用水合氯醛腹腔麻醉,冰上灌流、取腦,置于4%多聚甲醛中室溫浸泡后固定24 h。固定后的大腦進行梯度脫水處理:50%酒精,2 h;75%酒精,過夜;90%酒精,1 h;95%酒精Ⅰ,1 h;95%酒精Ⅱ,1 h;無水乙醇Ⅰ,1 h;無水乙醇Ⅱ,1 h;二甲苯Ⅰ,1 h;二甲苯Ⅱ,1 h;最后放入冬青油中過夜。脫水后的大腦用生物組織包埋機(天津天利航空機電有限公司)包埋制成蠟塊。石蠟切片機將蠟塊切成4 μm的薄片,貼附于涂有多聚賴氨酸的載玻片上,60 ℃烘箱中干燥2 h,脫蠟復水。蘇木素染色液浸染5 min,酸性分化液分化5 s,自來水沖洗2 min返藍,自來水沖洗2 min,剛果紅染色液浸染20~30 min,無水乙醇快速沖洗3次,二甲苯透明,中性樹膠封片,用熒光顯微鏡(BX61,Olympus,Japan)觀察并拍照。
2.3Tunel檢測皮層神經元的凋亡
取對照組和游泳組小鼠大腦石蠟切片,根據Tunel細胞凋亡檢測試劑盒說明書依次添加相應試劑。用熒光顯微鏡(BX61,Olympus,Japan)觀察并拍照。
2.4WesternBlot
10%水合氯醛腹腔麻醉小鼠,預冷的0.01 mol·L-1磷酸鹽緩沖液(PBS)冰上進行心臟灌注,至右心房流出清亮液體,斷頭取腦,迅速將小鼠大腦置于預冷的PBS中,小心剝離大腦皮質,將其置于添加有苯甲基磺酰氟的RIPA裂解液中,電動勻漿器勻漿,4 ℃、12 000 r·min-1離心10 min,所得上清即為皮層總蛋白。BCA法進行蛋白定量。樣品經10%SDS-PAGE分離目的蛋白、轉膜、5%的脫脂牛奶室溫封閉1 h后加入一抗,4 ℃孵育過夜。次日,用1×TBST漂洗3次,每次15 min,之后加入HRP標記的二抗于室溫孵育1 h,用1×TBST漂洗3次,每次15 min。ECL試劑盒顯影曝片。Quntity One 4.62進行條帶灰度分析。
2.5Morris水迷宮檢測
游泳前進行Morris水迷宮實驗測試小鼠空間學習記憶能力。實驗分為2個部分:(1)定位航行實驗:測量小鼠對水迷宮空間學習和記憶的獲取能力。實驗歷時7 d,第1~6天,每只小鼠每天訓練3次。在水迷宮的不同象限貼上標識,將小鼠按著標識面向池壁放入水中,記錄小鼠尋找并爬上平臺時所需時間(即逃避潛伏期)。如果小鼠在60 s內未找到平臺,用導引棒將其引至平臺,并在平臺上停留10 s,這時潛伏期記為60 s。第7天測試;(2)空間搜索實驗:在第8天撤除水下平臺,在同一入水點將小鼠面向池壁放入水中,記錄小鼠進入原先放置平臺的象限所花的時間(目標象限搜索時間百分比)和進入該象限的次數(通過目標區域次數),作為空間記憶的檢測指標。
2.6數據分析
用GraphPad Prism 6.0進行數據統計分析。實驗數據以平均數標準差(MeanSD)表示,組間比較采用配對t檢驗。P<0.05為差異有統計學意義。
3 結果
3.1APP/PS1轉基因小鼠鑒定
PCR結果顯示,APP/PS1轉基因小鼠分別約在350 bp和608 bp處觀察到APP和PS1目的帶,該位置DNA片段的大小與理論結果一致,而野生型小鼠沒有這2條目的帶,說明獲得APP/PS1轉基因小鼠(圖1)。
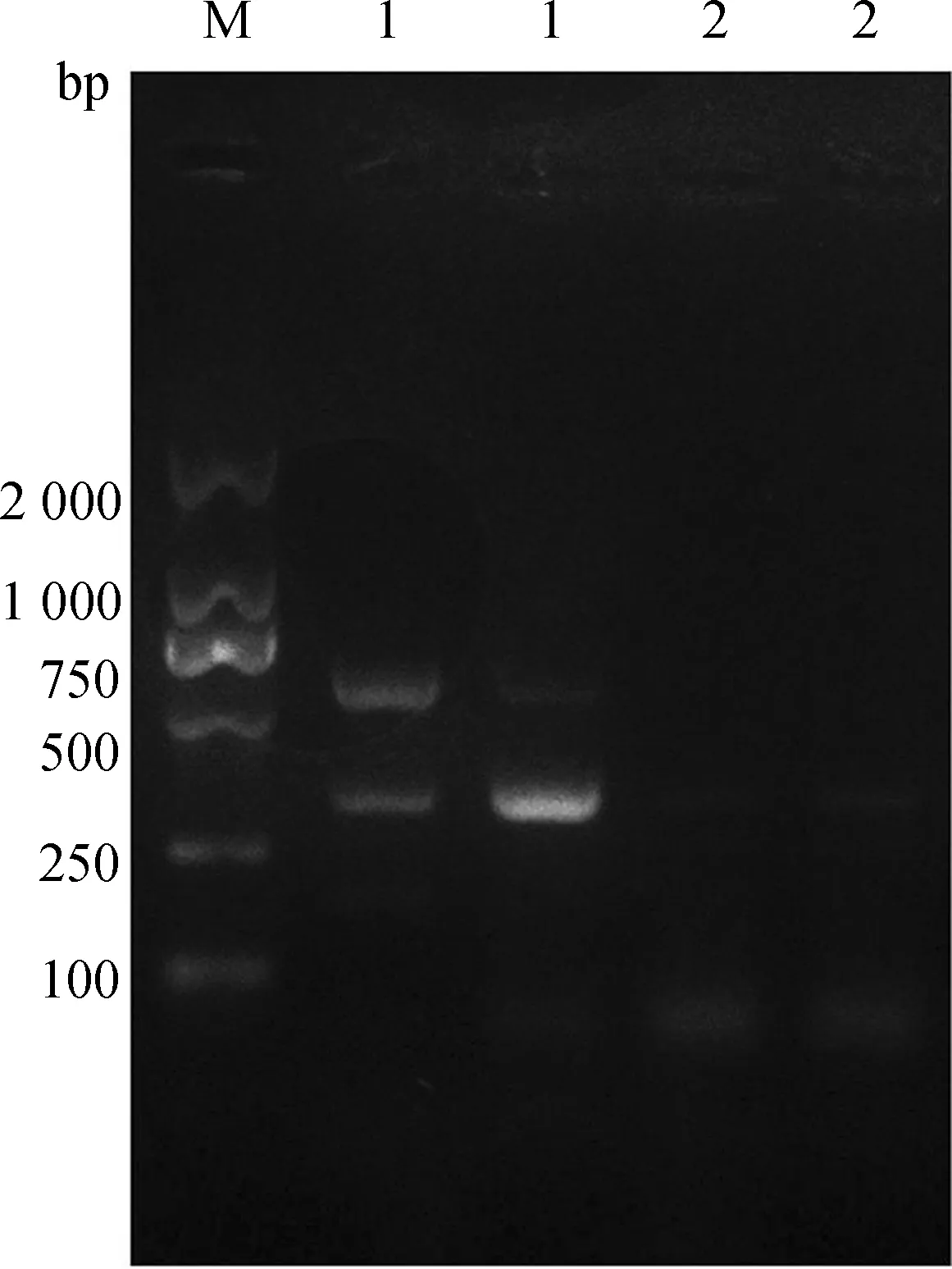
圖1 PCR鑒定APP/PS1轉基因小鼠Fig. 1 APP/PS1 transgenic mice identified by PCR
M. Marker, 1.APP/PS1 轉基因小鼠APP/PS1 transgenic mouse, 2. 野生型小鼠wild type mouse.
3.2游泳運動對Aβ斑的影響
剛果紅對Aβ斑的染色結果顯示,12月齡的對照組小鼠皮層內形成大量的Aβ斑,然而通過1個月的游泳運動,游泳組小鼠皮層內Aβ斑的數量極顯著減少(P<0.01)(圖2)。
3.3游泳運動對皮層神經元的影響
Tunel對神經元凋亡的檢測結果顯示,12月齡的對照組小鼠皮層內存在大量的神經元凋亡,而經過游泳運動的小鼠,皮層內神經元凋亡的數量極顯著減少(P<0.01)(圖3)。
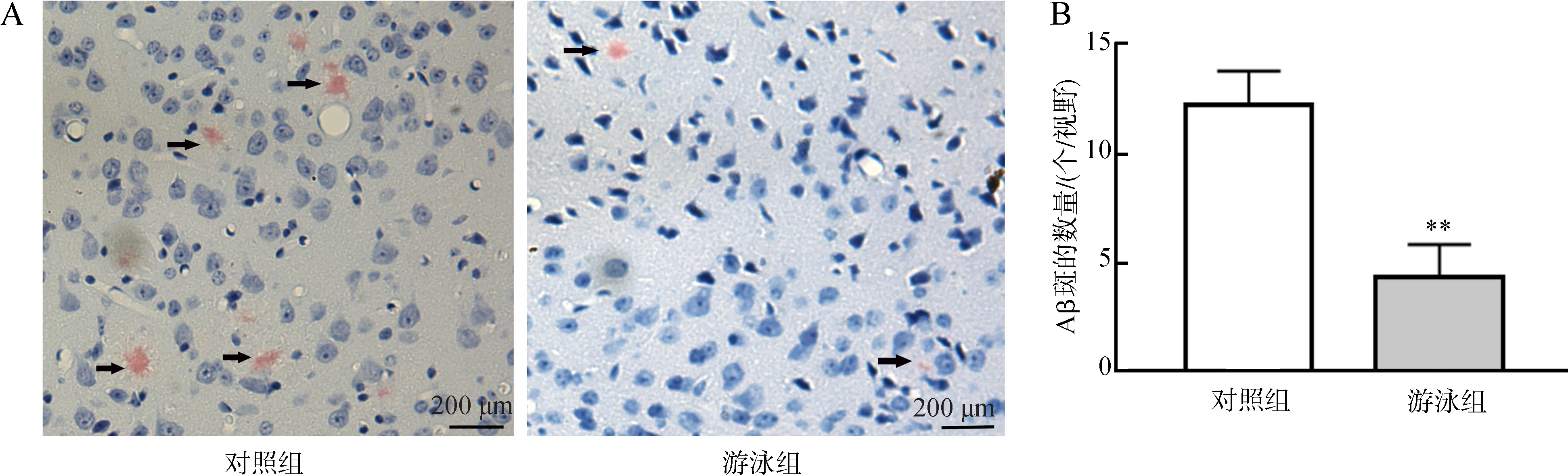
圖2 游泳運動對Aβ斑的影響Fig. 2 Effects of swimming on Aβ plaques
A. 剛果紅標記的Aβ斑, B. Aβ斑的統計分析;**P<0.01。
A. Aβ plaques marked by Congo red staining, B. statistical analysis of Aβ plaques;**P<0.01.
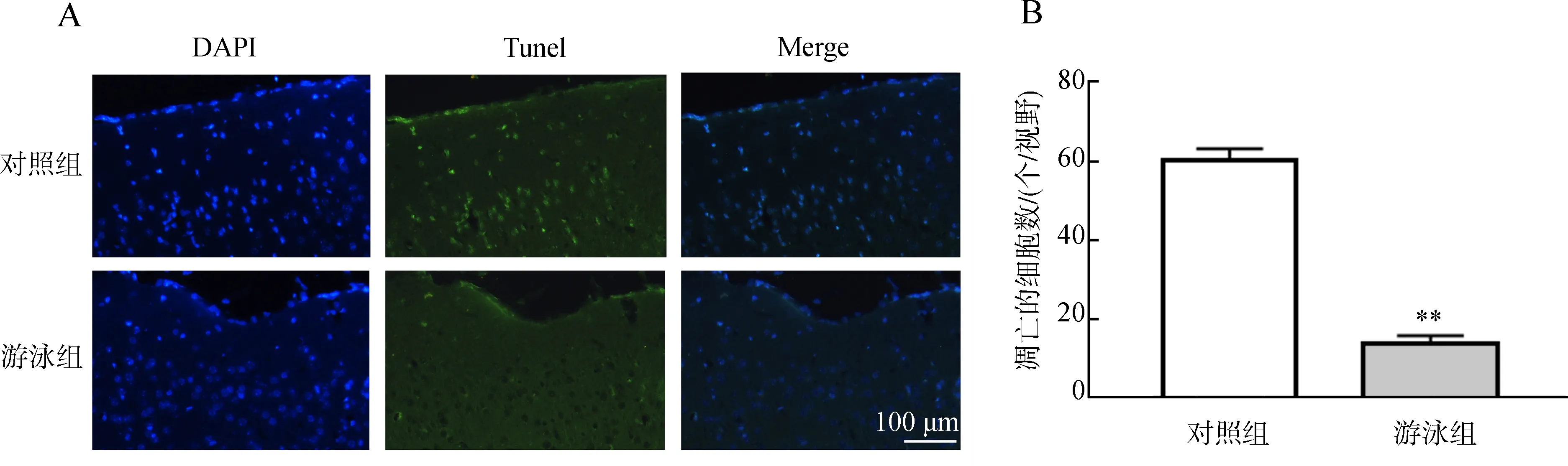
圖3 游泳運動對皮層神經元的影響Fig. 3 Effects of swimming on cortical neurons
A. Tunel標記的凋亡細胞, B. 凋亡細胞的統計分析;**P<0.01; DAPI. 染細胞核, Tunel. 斷裂的DNA, Merge. 凋亡細胞。
A. apoptotic cells identified by Tunel, B. statistical analysis of apoptotic cells;**P<0.01; DAPI. nucleus, Tunel. broken DNA, Merge. apoptotic cells.
3.4游泳運動對神經元PGC-1α和COXⅣ表達的影響
Western Blot檢測大腦皮質內PGC-1α和COX Ⅳ的表達。結果顯示,游泳運動促進了小鼠皮質內PGC-1α和COX Ⅳ表達的上調,說明游泳運動能夠促進線粒體的生成(圖4)。
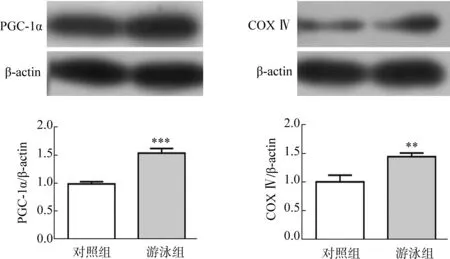
圖4 游泳運動對神經元PGC-1α和COX Ⅳ表達的影響Fig. 4 Effects of swimming on the expression of PGC-1α and COX Ⅳ in neurons
**P<0.01,***P<0.001.
3.5游泳運動對APP/PS1轉基因小鼠學習記憶能力的影響
在定位航行實驗中,前3 d內對照組和游泳組小鼠的逃避潛伏期都比較長,且差異無統計學意義,從第4天開始,游泳組小鼠所需要的時間比對照組少,且差異具有統計學意義(圖5:A)。在第8天撤出平臺,觀察小鼠的空間探索能力,發現游泳組小鼠穿過平臺所在象限的次數多于對照組小鼠,差異具有統計學意義(圖5:B)。觀察水迷宮的軌跡圖發現,對照組小鼠的運動軌跡比較復雜、沒有規律、耗時較多,而游泳組小鼠運動軌跡的規律比較明顯、耗時較少(圖5:C)。
4 討論
AD作為老年期退行性且不可逆的中樞神經系統性疾病,常導致患者進行性認知功能障礙、記憶力減退以及性格和行為改變等。目前,Aβ沉淀形成老年斑所導致的神經元凋亡或丟失是解釋AD發生的重要病理機制。APP/PS1轉基因小鼠攜帶有由小鼠朊病毒蛋白啟動子啟動表達的突變人類淀粉樣前蛋白(APPswe)和人類早老素(DeltaE9)基因,最早在3月齡時即可出現Aβ沉積形成老年斑的病理變化,且隨著月齡的增加,出現AD病理癥狀的表現越加明顯,因此該模型小鼠是用來研究AD較理想的動物模型(Jinetal.,2015;Zhaoetal.,2016)。本研究發現,12月齡的APP/PS1轉基因小鼠大腦皮層內形成大量由Aβ沉積所導致的老年斑,并觀察到神經元大量凋亡,與AD出現大量Aβ斑沉積和神經元凋亡或丟失的病理癥狀相吻合。
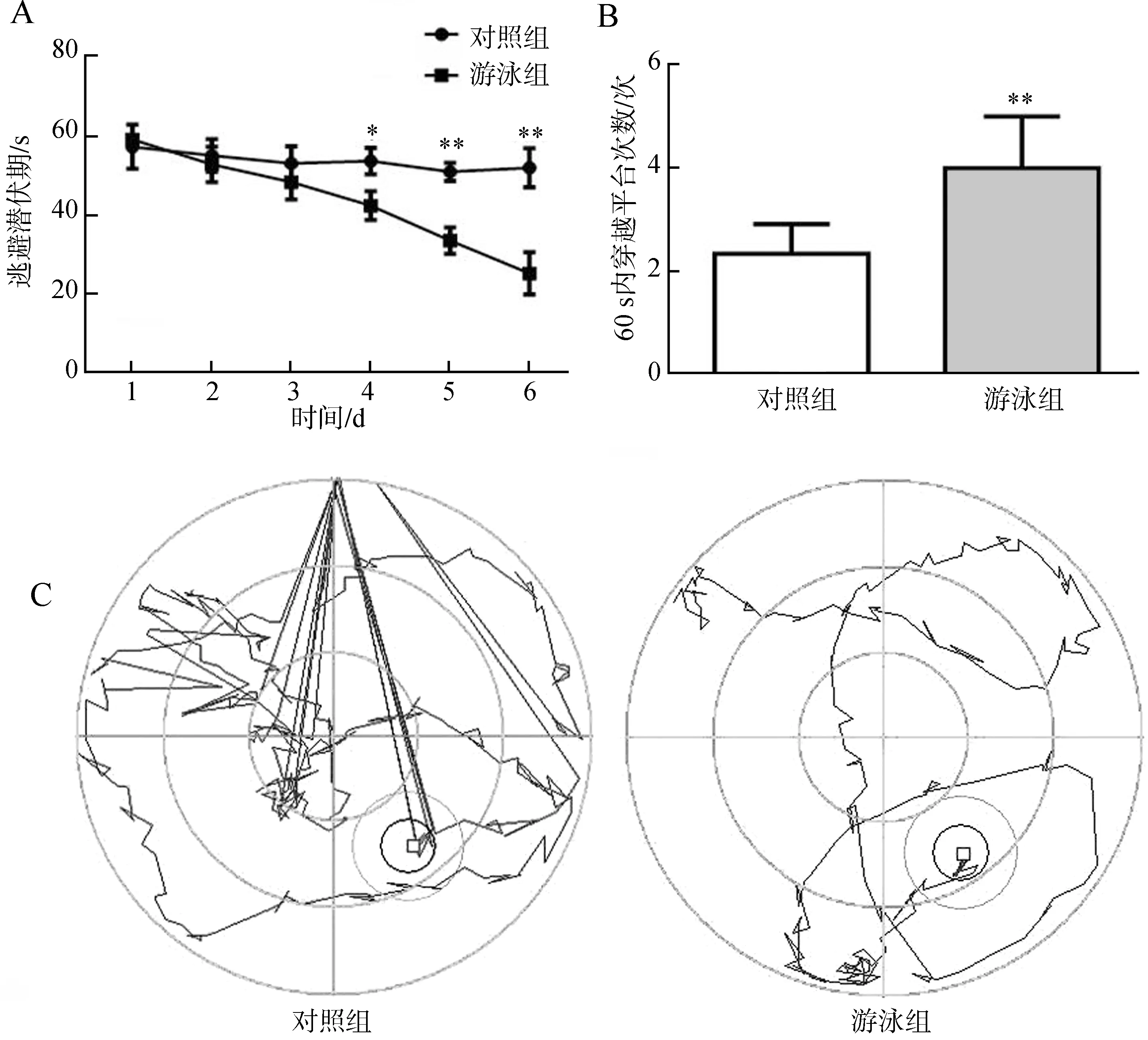
圖5 游泳運動對APP/PS1轉基因小鼠學習記憶能力的影響Fig. 5 Effects of swimming on learning and memory abilities in APP/PS1 transgenic mice
A.逃避潛伏期, B. 穿過平臺的次數, C. 游泳軌跡;*P<0.05,**P<0.01。
A. time of latency, B. passing times across the platform quadrant, C. swim-tracking path;*P<0.05,**P<0.01.
最近研究發現,Aβ可富集在線粒體內,導致線粒體氧化損傷、生成能力降低、動力學紊亂、結構破壞和功能降低等,這些是AD發生的早期事件,因此,促進線粒體的功能、抑制神經元凋亡或丟失可能是防治AD的策略之一(Piconeetal.,2014;Wuetal.,2014;劉艷萍等,2015)。對缺血再灌注的研究表明,促進線粒體生成可以增強線粒體的功能從而抑制神經元凋亡,發揮神經保護效應(Wangetal.,2014)。孕期的游泳運動可以誘導子代大腦線粒體生成,降低新生鼠缺氧缺血性腦損傷、增加運動能力、促進長時記憶能力,說明游泳運動行為可以通過促進線粒體生成發揮神經保護效應(Marcelinoetal.,2013,2016)。PGC-1α是線粒體生成的重要調控因子,促進PGC-1α的表達可以增強線粒體的體積分數、增加線粒體DNA的拷貝數、上調線粒體自身蛋白的表達,如COX Ⅳ促進線粒體生成,增強線粒體的功能發揮細胞保護效應(Abrahan & Ash,2016;王來,祝世功,2016)。本研究發現,12月齡的APP/PS1轉基因小鼠經過游泳運動后,大腦皮層PGC-1α和COX Ⅳ的表達上調,說明游泳運動能促進線粒體的生成。病理學的檢測發現,游泳組小鼠大腦皮質內Aβ沉積形成的老年斑數量減少、神經元凋亡數量降低,說明游泳運動可以降低Aβ斑的形成,抑制神經元凋亡或丟失。Morris水迷宮檢測APP/PS1轉基因小鼠的學習記憶能力表明,游泳組小鼠的逃避潛伏期小于對照組,60 s內穿過平臺次數多于對照組,對水迷宮軌跡圖的分析表明,對照組小鼠軌跡比較復雜、凌亂,而游泳組軌跡較為簡單、有規律,充分說明游泳運動可以抑制APP/PS1轉基因小鼠學習記憶能力的降低。
總之,本研究結果表明,游泳運動可以上調APP/PS1轉基因小鼠的PGC-1α的表達、促進線粒體的生成、抑制Aβ斑的形成、抑制神經元的凋亡或丟失、增強小鼠的學習記憶能力,該研究結果為探討AD的運動行為治療提供了依據。
崔建梅, 李小利, 付芳, 等. 2013. 游泳運動對衰老大鼠學習記憶、前額葉皮質氨基酸水平及nNOS表達的影響[J]. 吉林大學學報(醫學版), 39(4): 737-742.
劉艷萍, 陸征宇, 董強. 2015. 阿爾茨海默病病理改變的線粒體機制[J]. 神經損傷與功能重建, 10(3): 239-241.
欒海云, 李娜, 王桂華, 等. 2016. 游泳運動對D-半乳糖致衰老小鼠學習記憶和膽堿能神經系統功能的影響[J]. 中國運動醫學雜志, 35(1): 36-39.
王來, 祝世功. 2016. 線粒體生成與腦缺血再灌注損傷的研究進展[J]. 中國病理生理雜志, 32(8): 1478-1483.
Abrahan C, Ash JD. 2016. The potential use of PGC-1α and PGC-1β to protect the retina by stimulating mitochondrial repair[J]. Advances in Experimental Medicine & Biology, 854: 403-409.
Armand-Ugon M, Ansoleaga B, Berjaoui S,etal. 2017. Reduced mitochondrial activity is early and steady in the entorhinal cortex but it is mainly unmodified in the frontal cortex in Alzheimer's disease[J]. Current Alzheimer Research, 4. DOI: 10.2174/1567205014666170505095921.
Baek SH, Park SJ, Jeong JI,etal. 2017. Inhibition of drp1 ameliorates synaptic depression, Aβ deposition, and cognitive impairment in an Alzheimer’s disease model[J]. Journal of Neuroscience the Official Journal of the Society for Neuroscience, 37(20): 5099-5110.
Jin JL, Liou AK, Shi Y,etal. 2015. CART treatment improves memory and synaptic structure in APP/PS1 mice[J]. Scientific Reports, 5: 10224.
Marcelino TB, deLemos Rodrigues PI, Klein CP,etal. 2016. Behavioral benefits of maternal swimming are counteracted by neonatal hypoxia-ischemia in the offspring[J]. Behavioural Brain Research, 312: 30-38.
Marcelino TB, Longoni A, Kudo KY,etal. 2013. Evidences that maternal swimming exercise improves antioxidant defenses and induces mitochondrial biogenesis in the brain of young Wistar rats[J]. Neuroscience, 246(5): 28-39.
Picone P, Nuzzo D, Caruana L,etal. 2014. Mitochondrial dysfunction: different routes to Alzheimer’s disease therapy[J]. Oxidative Medicine & Cellular Longevity, 2014(2): 780179.
Reddy PH, Manczak M, Yin X. 2017. Mitochondria-division inhibitor 1 protects against amyloid-β induced mitochondrial fragmentation and synaptic damage in Alzheimer’s disease[J]. Journal of Alzheimers Disease Jad, 58(1): 147-162.
Valasani KR, Sun Q, Fang D,etal. 2016. Identification of a small molecule cyclophilin D inhibitor for rescuing Aβ-mediated mitochondrial dysfunction[J]. ACS Medicinal Chemistry Letters, 7(3): 294-299.
Wang D, Liu X, Liu Y,etal. 2017. The effects of cardiotrophin-1 on early synaptic mitochondrial dysfunction and synaptic pathology in APPswe/PS1dE9 mice[J]. Journal of Alzheimers Disease Jad, 59(4): 1255.
Wang L, Chen M, Yuan L,etal. 2014. 14, 15-EET promotes mitochondrial biogenesis and protects cortical neurons against oxygen/glucose deprivation-induced apoptosis[J]. Biochemical & Biophysical Research Communications, 450(1): 604-609.
Wu Z, Zhu Y, Cao X,etal. 2014. Mitochondrial toxic effects of Aβ through mitofusins in the early pathogenesis of Alzheimer’s disease[J]. Molecular Neurobiology, 50(3): 986-996.
Ye CY, Lei Y, Tang XC,etal. 2015. Donepezil attenuates Aβ-associated mitochondrial dysfunction and reduces mitochondrial Aβ accumulationinvivoandinvitro[J]. Neuropharmacology, 95: 29-36.
Yu Q, Du F, Douglas JT,etal. 2017. Mitochondrial dysfunction triggers synaptic deficits via activation of p38 MAP kinase signaling in differentiated alzheimer's disease trans-mitochondrial cybrid cells[J]. Journal of Alzheimers Disease Jad, 59(1): 223-239.
Zhao FL, Qiao PF, Yan N,etal. 2016. Hydrogen sulfide selectively inhibits γ-secretase activity and decreases mitochondrial aβ production in neurons from APP/PS1 transgenic mice[J]. Neurochemical Research, 41(5): 1145-1159.
EffectsofSwimmingExerciseonLearningandMemoryAbilityofAPP/PS1TransgenicMice
FAN Zongshan1, 2, JING Guangwei3, ZHAO Haipeng4*
(1. Henan Vocational College of Agriculture, Zhongmou, Henan Province 451450, China; 2. Physical Education Institute of Henan Normal University, Xinxiang, Henan Province 453002, China; 3. Henan Jigong Mountain National Nature Reserve Management Bureau, Xinyang, Henan Province 464000, China; 4. School of Life Science, Henan University, Kaifeng, Henan Province 475001, China)
To investigate the effects of swimming on learning and memory ability of APP/PS1 transgenic mice. 11-month-old male APP/PS1 transgenic mice were randomly divided into control group and swimming group. The control group had no exercise, and the swimming group had swimming training for 1 month. The expression of Aβ plaques, neuron apoptosis, mitochondria biogenesis-related protein expression and the learning and memory of APP/PS1 transgenic mice were determined by Congo red staining, Tunel assay, Western Blot and Morris water maze. The result showed that swimming exercise reduced the formation of Aβ plaques and neuronal apoptosis, promoted mitochondria biogenesis in the cerebral cortex of APP/PS1 transgenic mice, and enhanced the ability of learning and memory of APP/PS1 transgenic mice. This study suggested that swimming can promote the learning and memory ability of APP/PS1 transgenic mice by facilitating mitochondria biogenesis, and thus can be used as a treatment for the prevention and treatment of Alzheimer’s disease.
APP/PS1 transgenic mice; swimming; learning and memory
10.11984/j.issn.1000-7083.20170243
2017-08-11接受日期2017-11-10
樊宗山(1979—), 男, 碩士研究生, 主要研究方向為運動生理與體育教學, E-mail:fanzs161@sohu.com
*通信作者Corresponding author, 男, 博士, 副教授, 研究方向:動物學, E-mail:hpzhao1980@qq.com
R339.4
A
1000-7083(2017)06-0663-06
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
發明與創新(2022年30期)2022-10-03 08:40:56
學苑創造·A版(2020年9期)2020-10-13 09:41:02
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
作文周刊·小學一年級版(2016年27期)2017-06-03 23:21:17
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
海外文摘(2016年4期)2016-04-15 22:28:55
小星星·閱讀100分(低年級)(2015年10期)2015-10-22 08:30:04
