家雞G蛋白偶聯受體161的基因克隆、分子進化和組織表達
2017-12-12 06:26:08祝國強莫春橫李正陽王亞軍李娟
四川動物 2017年6期
關鍵詞:小鼠
祝國強, 莫春橫, 李正陽, 王亞軍, 李娟
(四川大學生命科學學院,生物資源與生態環境教育部重點實驗室,成都610065)
家雞G蛋白偶聯受體161的基因克隆、分子進化和組織表達
祝國強, 莫春橫, 李正陽, 王亞軍, 李娟*
(四川大學生命科學學院,生物資源與生態環境教育部重點實驗室,成都610065)
G蛋白偶聯受體161(GPR161)是G蛋白偶聯受體家族孤兒受體家族成員,在哺乳動物晶狀體發育調節和神經胚形成中具有重要作用。近年來研究發現該蛋白罕見地擁有類似支架蛋白的結構特征,暗示其信號轉導機制不同于其他G蛋白偶聯受體。本研究以家雞Gallusgallus為動物模型,探究GPR161基因序列信息、分子遺傳進化關系以及其組織表達圖譜。家雞GPR161基因編碼區序列全長1 566 bp,編碼具521個氨基酸的前體蛋白。序列分析顯示,家雞GPR161基因編碼區與人Homosapiens、小鼠Musmusculus、斑馬魚Daniorerio的氨基酸相似度分別為83.0%、82.6%、65.8%。分子進化遺傳分析結果顯示,GPR161基因在家雞與斑馬魚的進化關系比家雞與人或小鼠都更為疏遠。利用熒光定量PCR探究家雞GPR161基因在各組織的表達分布,結果顯示,家雞GPR161基因mRNA在精巢或卵巢、大腦、心臟、肌肉中有較高表達。本研究是鳥類中關于GPR161基因的首次報道,研究結果為進一步探究GPR161基因在鳥類中的生理效應提供參考。
家雞;GPR161; 基因克隆; 分子進化; 組織表達
G蛋白偶聯受體是細胞表面受體中最大并最具多樣性的家族,其家族成員均具典型7次跨膜結構,介導視覺、嗅覺、行為等眾多生理活動(Morris & Malbon,1999)。在人中已發現并獲得鑒定的G蛋白偶聯受體超過1 000種,約占人類基因總數的5%(Consortium,2004;Zhangetal.,2006),截至目前,該數量仍在緩慢增加,凸顯G蛋白偶聯受體在細胞功能實現中的重要作用。
G蛋白偶聯受體161(G protein-coupled receptor 161,GPR161)屬于G蛋白偶聯受體家族孤兒受體家族成員,這一類受體家族成員的內源性配體有待鑒定(Gainetdinovetal.,2004)。盡管缺乏其內源性配體信息,但GPR161被報道在調節晶狀體發育和神經胚形成中具有重要作用。空泡化晶狀體癥小鼠突變體帶有先天白內障和神經管畸形,研究發現該表型源于GPR161基因8 bp堿基缺失(Mattesonetal.,2008)。原位雜交實驗結果進一步顯示,GPR161基因的表達伴隨著晶狀體發育的完整階段,側面印證GPR161功能缺失可能是白內障的成因(Mattesonetal.,2008)。進一步的研究發現,GPR161的生理功能不僅涉及空泡化晶狀體癥,還參與調控左右心室形成、乳腺癌細胞的增殖以及垂體柄阻斷綜合癥的發生(Leungetal.,2008;Feiginetal.,2014;Karacaetal.,2014)。
在傳統的G蛋白偶聯受體信號通路中,胞外配體結合受體激活G蛋白,激活的亞基包括G蛋白α亞基以及β和γ亞基進一步激活下游效應器,包括各種激酶等,促進通路持續(Morris & Malbon,1999)。在信號轉導進程中,支架蛋白成為重要的連接器。各種GTP酶、激酶、磷酸化酶等信號蛋白在支架蛋白介導下獲得次序激活,掌控多樣生理效應。以支架蛋白A類激酶錨定蛋白為例,不同的A類激酶錨定蛋白錨定Ⅰ型或Ⅱ型蛋白激酶A,釋放二級信號分子,最終激活環磷酸腺苷感受信號通路(Wong & Scott,2004;Langeberg & Scott,2015)。此外,支架蛋白與G蛋白偶聯受體具有物理連接,對G蛋白偶聯受體的定位和活性調節有重要意義(Ritter & Hall,2009)。研究表明,GPR161基因除了具G蛋白偶聯受體經典的結構特征,其C末端還罕見地包含作為支架蛋白A類激酶錨定蛋白共有的兩性分子螺旋,提示該蛋白可以直接錨定、富集Ⅰ型蛋白激酶A,在信號轉導進程中發揮更為重要的主導作用,其結構意義及潛在的效應機制備受關注(Mukhopadhyayetal.,2013;Bachmannetal.,2016)。
綜上,GPR161基因參與介導機體重要生理效應,其結構特征亦值得進一步探究。至今,針對GPR161基因的研究報道仍然相對匱乏,且GPR161基因序列在鳥類中至今仍未獲得鑒定。家雞Gallusgallus是鳥類模式生物,是脊椎動物進化中最為重要的過渡類群。本研究首次報道家雞GPR161基因的編碼區序列,預測其二級結構,同時也檢測其在成體家雞中的組織表達圖譜,研究結果為闡釋GPR161基因在脊椎動物中的生理效應提供基礎信息。
1 材料與方法
1.1材料
本實驗所用動物為羅曼粉成年家雞,購自四川牧星養雞場;引物合成、DNA測序工作由成都擎科公司完成;組織保存和RNA提取所用的RNAzol購自Molecular Research Central;MMLV逆轉錄酶、dNTP、KOD-Fx高保真聚合酶、Easy-Taq酶、pTA-2載體、限制性內切酶、T4 DNA連接酶均購自大連寶生物工程有限公司(TaKaRa);分子克隆宿主大腸桿菌EscherichiacoliDH5α由本實驗室制備保存;熒光定量PCR儀Bio-Rad CFX96、熒光染料Eva Green、96孔板、塑料封膜購自Bio-Rad。
1.2方法
1.2.1總RNA提取取家雞組織包括脂肪、腎上腺、腎臟、肝臟、肺、肌肉、卵巢、胰腺、皮膚、脾臟、精巢等,迅速放入液氮中速凍,并用研缽研棒保證組織浸泡在液氮中充分研磨成粉末。后續步驟嚴格按照RNA extraction kits說明書(TaKaRa)提取總RNA:取約60 μg組織粉末,與600 μL RNAzol混合;補充240 μL DEPC滅菌水,用渦旋儀混勻約15 s;在高速冷凍離心機中,于4 ℃、12 000 r·min-1離心15 min;取上清,加入3 μL阿司咪唑,渦旋15 s后于4 ℃、12 000 r·min-1冷凍離心10 min;取上清,加入等體積異丙醇,于4 ℃、12 000 r·min-1冷凍離心10 min;小心吸取上清并棄掉,加入75%乙醇漂洗總RNA沉淀,于4 ℃、8 000 r·min-1冷凍離心3 min;重復上一步,漂洗RNA沉淀;棄上清,用20 μL DEPC滅菌水溶解沉淀,及時構建cDNA文庫或于-80 ℃冰箱保存。
1.2.2cDNA模板制備以各組織總RNA為模板進行逆轉錄構建cDNA文庫,具體步驟如下:取2 μg各組織的總RNA樣品,與1 μL Oligo-dT混合;補DEPC滅菌水至總體積5 μL (Oligo-dT終濃度為0.5 μg·μL-1),混勻;于PCR儀中70 ℃加熱10 min后,立即取出放置于冰上10 min;加入2 μL 5×RT buffer,0.5 μL MMLV逆轉錄酶,0.5 μL dNTPs,補充DEPC滅菌水至總體積10 μL,混勻;于PCR儀中42 ℃反應1.5 h,后又70 ℃反應10 min結束;取反應所得產物加入70 μL滅菌水,所得混合溶液即為cDNA模板。
1.2.3載體構建根據Ensemble數據庫提供的家雞基因組信息,遵循引物設計原理并使用DNAMAN 8.0(Lynnon Biosoft)設計家雞GPR161基因編碼區全長引物(上游引物cGPR161-U1和下游引物cGPR161-L1)和組織表達所需的熒光定量引物(上游引物cGPR161-qU1和下游引物cGPR161-qL1)。以家雞脂肪組織cDNA為模板,使用編碼區全長引物對GPR161基因的開放閱讀框(ORF)區域進行擴增。PCR擴增體系如下:5 μL 2×KOD緩沖液,2 μL脂肪組織cDNA模板,2 μL dNTPs,上游引物cGPR161-U1和下游引物cGPR161-L1各0.1 μL,0.5 μL KOD-Fx聚合酶,0.3 μL去離子滅菌水。PCR擴增條件如下:94 ℃預變性2 min;98 ℃變性10 s,60 ℃退火30 s,68 ℃延伸2 min,34個循環;72 ℃最后延伸10 min。然后分別取3 μL反應產物,用1%瓊脂糖凝膠電泳檢測PCR擴增結果。
取4.5 μL PCR產物與0.5 μL 10×A-attachment Mix混合,置于PCR儀中60 ℃反應30 min,保證KOD-Fx聚合酶擴增的平末端產物的3’末端加上堿基A;反應混合液與pTA-2載體進行4 ℃過夜連接;將連接產物轉入DH5α感受態細胞,并均勻涂布在加有抗氨芐的LB固體培養皿中進行藍白斑篩選;利用pTA-2載體通用引物挑選特異性陽性單克隆,送成都擎科公司進行雙向測序。
1.2.4生物信息分析對獲得的家雞GPR161氨基酸序列與NCBI數據庫中其他物種已知或預測序列,使用DNAMAN 8.0(Lynnon Biosoft)和在線網站ESPript(Easy Sequencing in PostScript)進行氨基酸相似度比對(Robert & Gouet,2014),分子進化遺傳學分析采用最大似然法(Maximum Likelihood,ML)依托MEGA 6.0完成(Tamuraetal.,2013),基因功能預測使用在線BioGRID數據庫(Biological General Repository for Interaction Datasets)(Chatr-aryamontrietal.,2016)。
1.2.5組織表達采用家雞不同組織的cDNA模板,以熒光定量PCR方法檢測各組織的表達情況。熒光定量PCR的擴增體系如下:2%DMSO,9.5 μL去離子滅菌水,2 μL 10×buffer,6 μL cDNA模板,0.4 μL dNTPs,上游引物cGPR161-qU1和下游引物cGPR161-qL1各0.2 μL,1 μL熒光染料Eva Green,1 μL Easy-Taq酶,總體積為20 μL。熒光定量PCR的擴增條件如下:94 ℃預變性2 min;94 ℃變性20 s,60 ℃退火15 s,72 ℃延伸20 s,39個循環,然后按照0.5 ℃/5 s的速度從70 ℃到95 ℃向上升溫熔解,生成熔解曲線。熒光定量PCR的結果按照比較CT值法(2-ΔΔCT法)進行數據處理(Schmittgen & Livak,2008),步驟如下:首先將各組織樣品結果GPR161的CT值減去對應組織內參基因β-actin的CT值,得到各組織的ΔCT值;然后將GPR161在大腦中的表達量選作參考,并將上一步分別所得的各組織的ΔCT值減去大腦的ΔCT值,得到ΔΔCT值;最后計算2-ΔΔCT值,得到各組織中GPR161相對大腦中GPR161的表達量倍數,即GPR161在各組織間的相對表達量。
2 結果
2.1家雞GPR161基因的克隆
參照NCBI數據庫中預測的家雞GPR161基因序列和Ensemble數據庫中的家雞全基因組設計上、下游PCR引物(表1),采用家雞脂肪組織cDNA為模板,使用高保真酶KOD擴增家雞GPR161序列。凝膠電泳顯示,擴增條帶大小與預期一致,略大于1 500 bp(圖1)。將該片段與pTA-2載體連接后轉化到DH5α中,經藍白斑篩選得到陽性單克隆。提取質粒,酶切檢驗確保重組質粒的完整性后,選取3個單克隆質粒送公司測序。
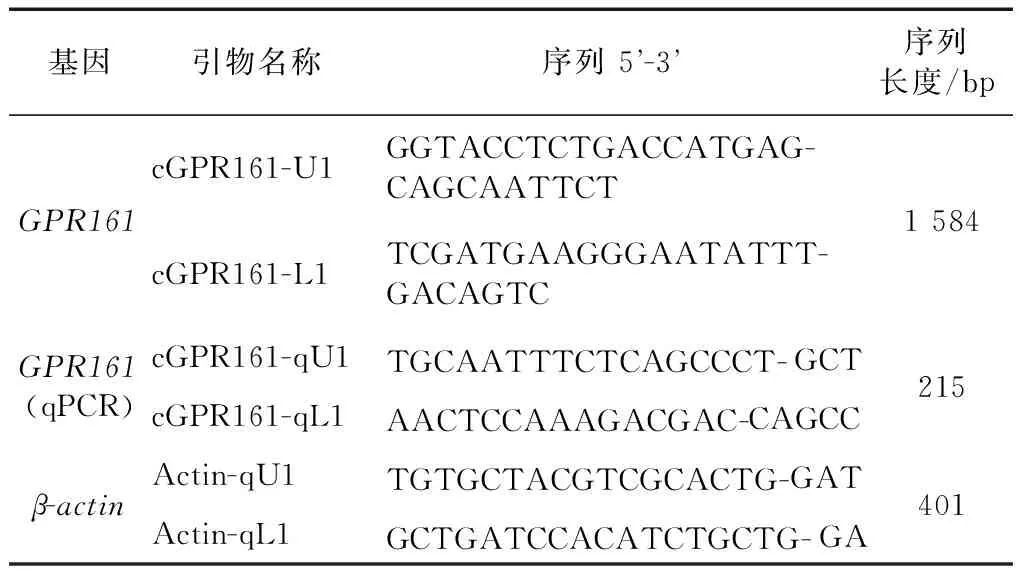
表1 引物序列Table 1 Primer sequence
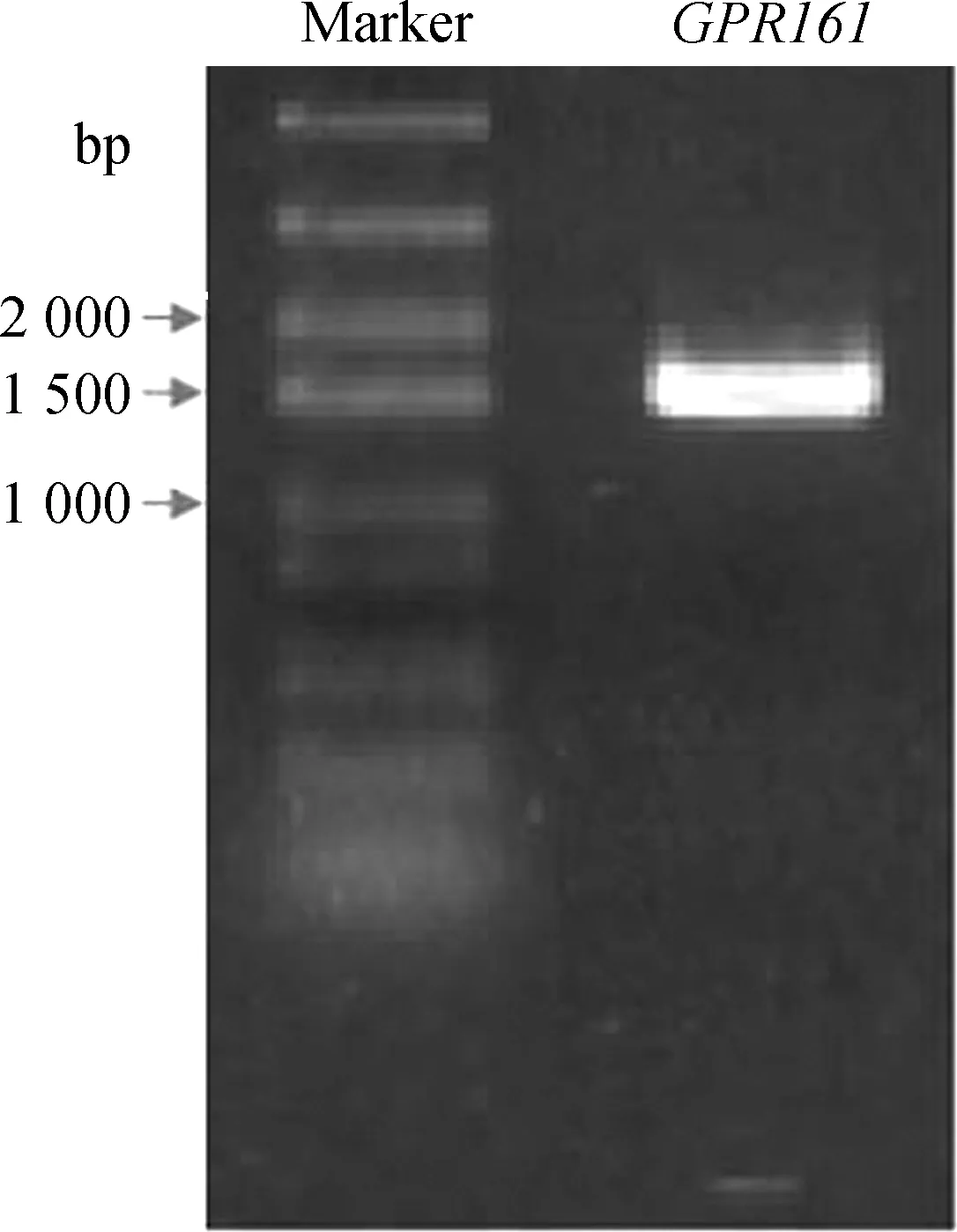
圖1 家雞GPR161基因擴增Fig. 1 PCR amplification of chicken GPR161 gene
2.2家雞GPR161基因的序列分析
測序結果顯示,家雞GPR161基因編碼區ORF長度為1 566 bp,編碼521個氨基酸。與家雞全基因組序列比對后發現,家雞GPR161基因位于家雞1號染色體上,編碼區沒有內含子。將家雞GPR161基因編碼區氨基酸序列與人Homosapiens、小鼠Musmusculus、斑馬魚Daniorerio的進行比對(圖2),發現其與人、小鼠、斑馬魚的相似度分別為83.0%、82.6%、65.8%。家雞GPR161基因含有G蛋白偶聯受體的7次跨膜區經典結構,同時還含有支架蛋白A類激酶錨定蛋白的兩性分子螺旋結構(圖2)。

圖2 家雞GPR161與斑馬魚、人、小鼠的基因編碼區氨基酸序列比對Fig. 2 Alignment of the amino acid sequence of GPR161 gene from chicken, zebrafish, human and mouse
TM1~7示經典跨膜區域, 兩性分子螺旋結構在方框中標識。
TM1-7 shows the typical transmembrane domains, the amphipathic helix is indicated.
2.3家雞GPR161基因的分子進化遺傳學分析
盡管GPR161基因序列信息目前只在人、小鼠、斑馬魚和家雞中得到鑒定,但是隨著測序技術的發展和越來越多物種的基因組被破譯,不同物種中未經實驗數據驗證的GPR161基因預測序列或片段也能夠從Ensemble數據庫獲得。為了更加全面地探究GPR161基因在不同物種間的分子遺傳學進化,分子系統進化樹結果顯示,家雞與火雞Meleagrisgallopavo的同源性最高,且與爬行類中華鱉Pelodiscussinensis、鳥類斑胸草雀Taeniopygiaguttata分子進化關系也較為接近,人和小鼠次之,與斑馬魚進化關系則相對較遠(圖3)。
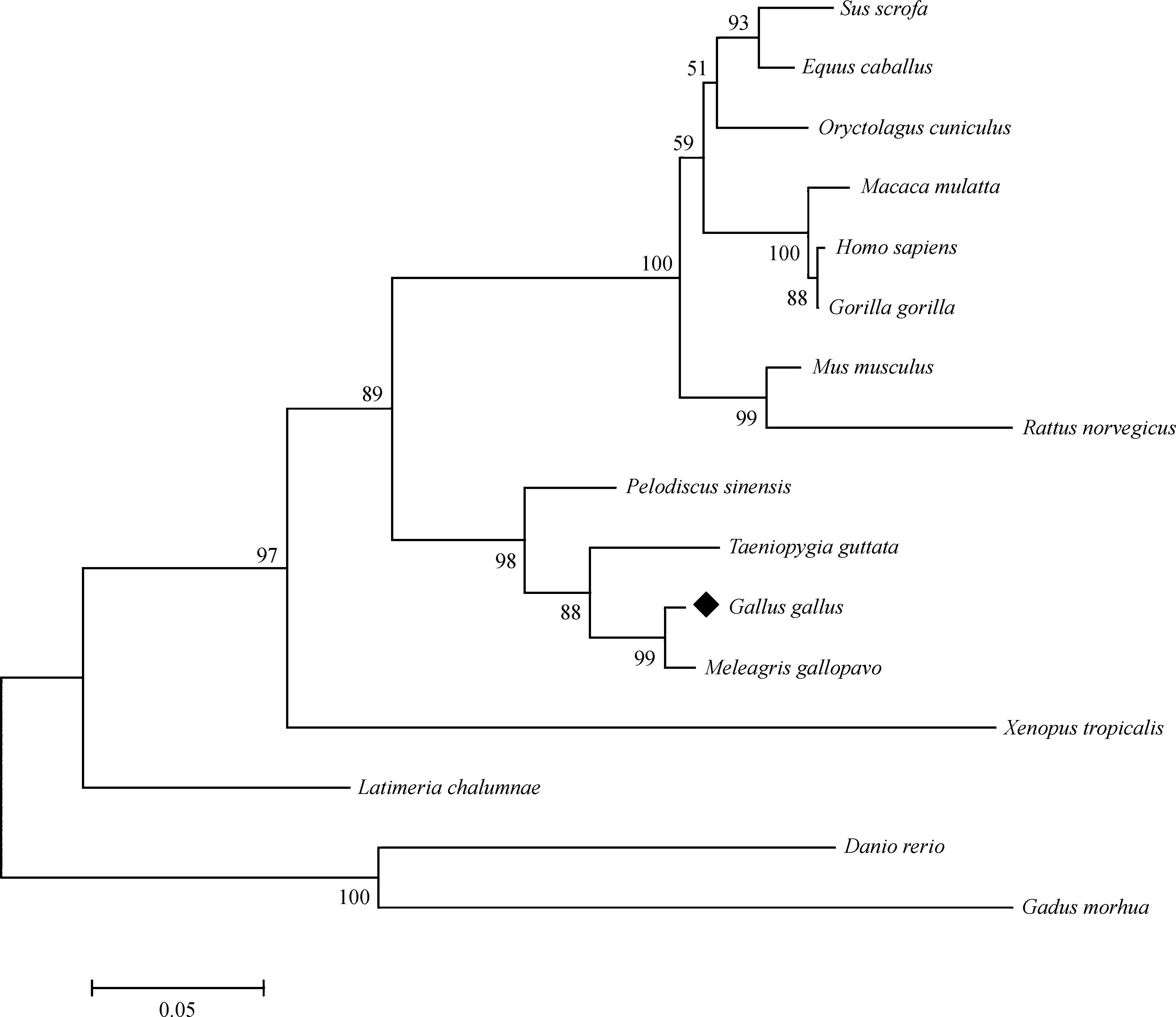
圖3 GPR161基因的分子系統進化樹Fig. 3 Molecular phylogenetic tree of GPR161 gene
2.4家雞GPR161基因的功能預測
依托于測序產生的大數據,對某個基因或基因家族的生物信息學進行功能注釋,目前已逐漸成為分析和預測基因功能的重要技術。在線BioGRID數據庫預測了可能與GPR161基因有蛋白相互作用的基因(圖4)。對蛋白互作網絡的GO功能注釋分析顯示,GPR161主要參與G蛋白偶聯受體信號通路、腹側神經管平滑信號通路的負調控、cAMP生物合成的正調控、初級纖毛的生成等生理功能,比如PPKACA、PPKACB、PPKACG是蛋白激酶A的組成亞基,直接介導cAMP信號。
2.5家雞GPR161基因mRNA組織表達分析
為探究GPR161基因在家雞各組織中的生理學效應差異,以成體家雞各組織cDNA為模板,用熒光定量PCR方法檢測GPR161基因在家雞各組織中的mRNA水平表達分布情況。各組織的樣本數均大于或等于4。結果顯示,GPR161基因mRNA在家雞的精巢或卵巢、大腦、心臟、肌肉中有較高的表達,但在脂肪、腎臟、肺中表達量相對較低,在肝臟、胰腺、皮膚、脾臟中則檢測到更弱的表達信號(圖5)。
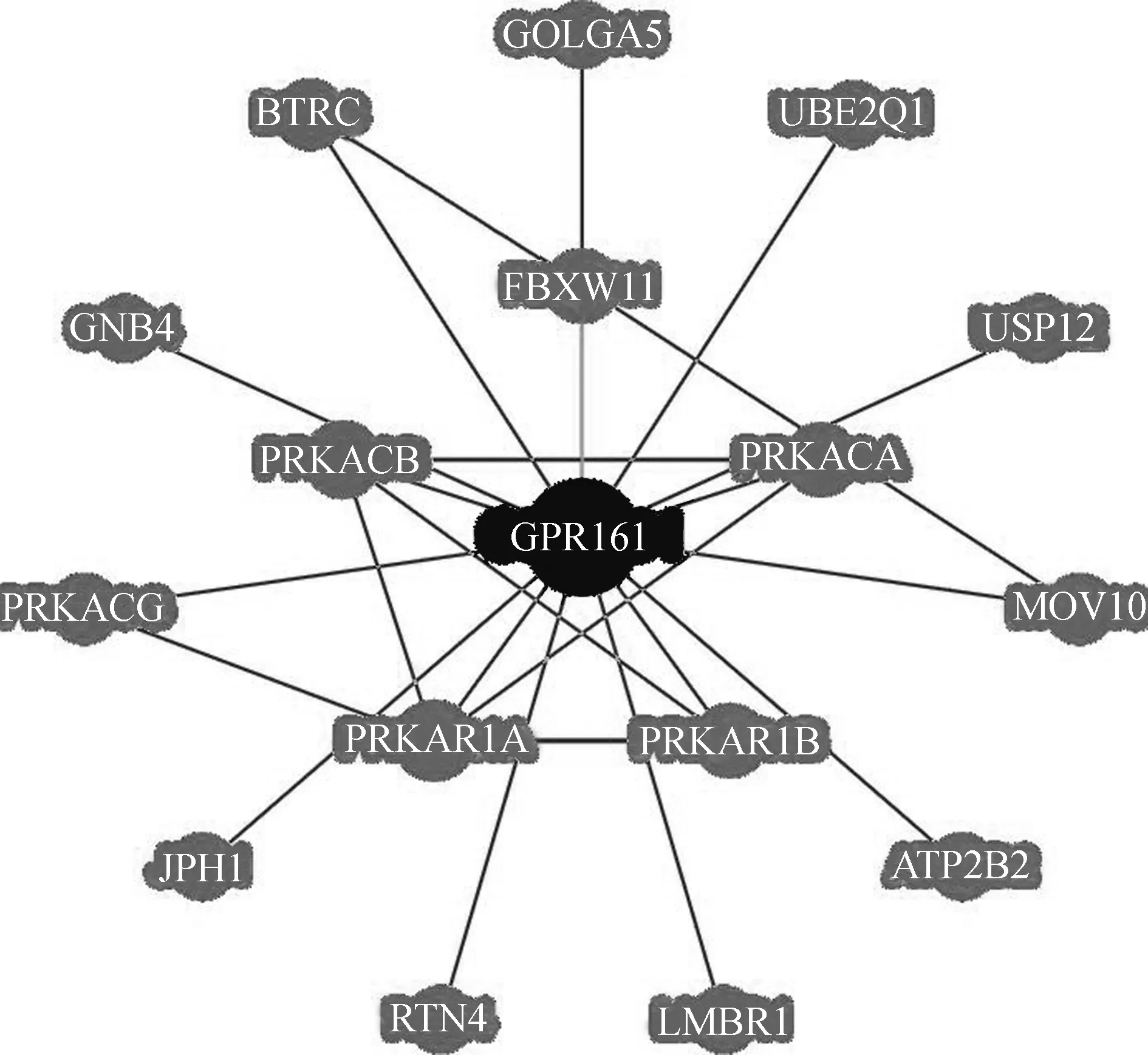
圖4 GPR161基因蛋白-蛋白間互作網絡Fig. 4 The network of protein-protein interactions with GPR161 gene
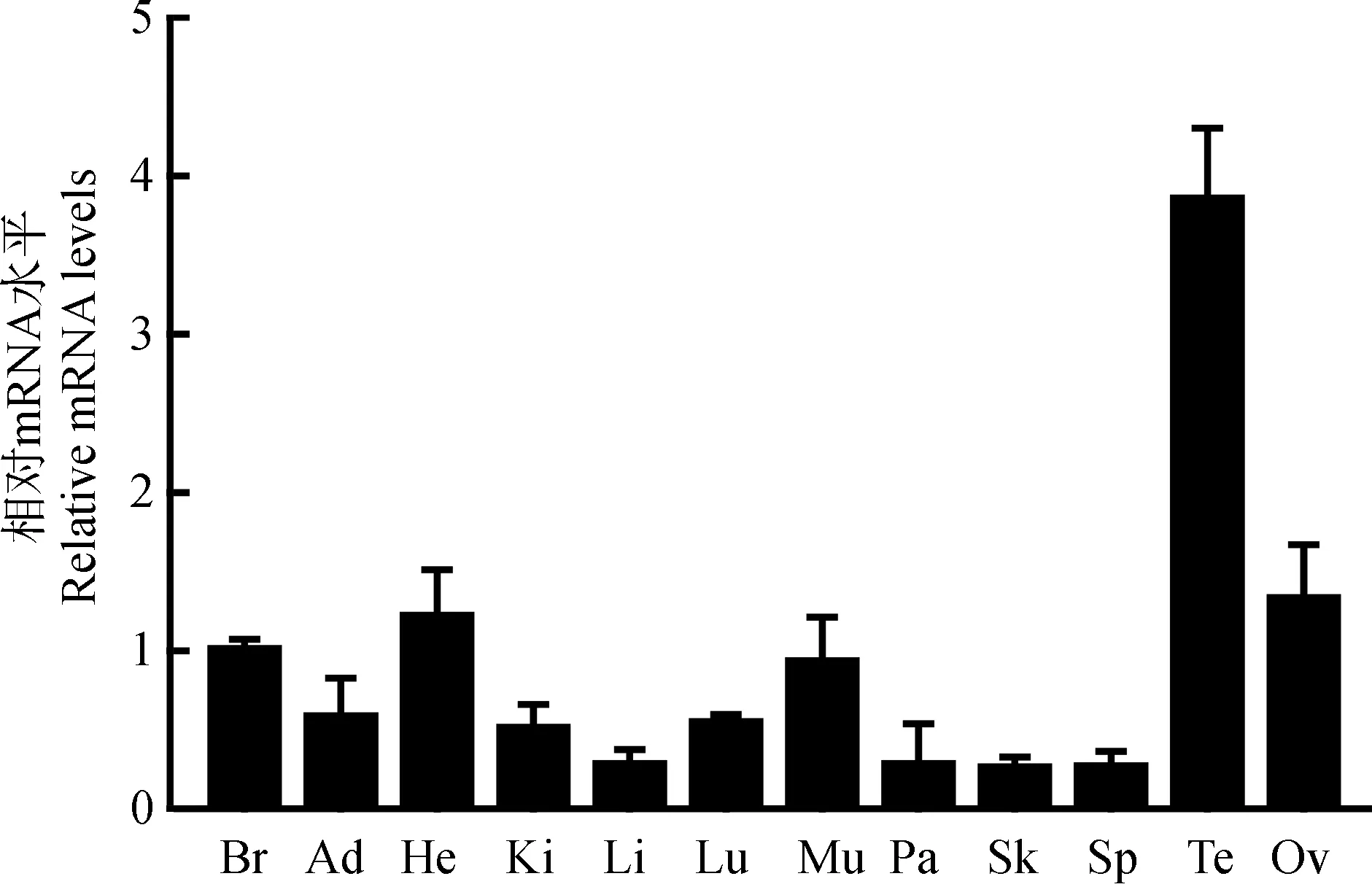
圖5 熒光定量PCR檢測GPR161基因mRNA在家雞各組織中的表達Fig. 5 qPCR detection of GPR161 gene mRNA expression in chicken tissues
Br. 大腦, Ad.脂肪, He. 心臟, Ki. 腎臟, Li. 肝臟, Lu. 肺, Mu. 肌肉, Pa. 胰腺, Sk. 皮膚, Sp. 脾臟, Te. 精巢, Ov. 卵巢。
Br. brain, Ad. adipose, He. heart, Ki. kidney, Li. liver, Lu. lung, Mu. muscle, Pa. pancreas, Sk. skin, Sp. spleen, Te. testis, Ov. Ovary.
3 討論
GPR161基因被報道在晶狀體發育、神經胚形成、左右心室形成、乳腺癌細胞增殖、垂體柄阻斷綜合癥的發生等疾病中扮演重要角色(Leungetal.,2008;Mattesonetal.,2008;Feiginetal.,2014;Karacaetal.,2014)。該基因在結構上罕見地擁有支架蛋白A類激酶錨定蛋白類似結構,凸顯其獨特結構特征并暗示其特別的調控機制。本研究首次報道家雞GPR161基因的序列信息、分子進化關系以及其組織表達圖譜。
家雞GPR161基因編碼區ORF長度為1 566 bp,編碼521個氨基酸。序列分析顯示,家雞GPR161氨基酸序列與哺乳動物具有較高的序列相似性(人:83.0%;小鼠:82.6%),而與斑馬魚的相似度相對較低(斑馬魚:65.8%)。分子進化遺傳分析表明,GPR161基因在家雞與火雞的同源性最高,且與爬行類中華鱉、鳥類斑胸草雀分子進化關系也較為接近,人和小鼠次之,而與斑馬魚進化關系相對較遠。
本研究也對GPR161的生理功能做了初步預測。通過在線BioGRID數據庫預測了GPR161參與G蛋白偶聯受體信號通路、cAMP生物合成的正調控等生理功能,這與目前在初級纖毛的發育等研究報道一致(Mukhopadhyayetal.,2013;Bachmannetal.,2016;Paletal.,2016)。利用熒光定量PCR分析技術解析獲得家雞GPR161基因組織表達圖譜,為探究其在各器官的生理學意義提供了參考依據。家雞GPR161基因主要在精巢或卵巢、大腦、心臟、肌肉組織中高表達,該研究結果與人、小鼠、斑馬魚的報道基本一致。在人和小鼠的大腦、神經管、垂體等神經系統中,GPR161基因mRNA被檢測到高水平表達(Mukhopadhyayetal.,2013;Karacaetal.,2014)。在斑馬魚的胚胎發育中,GPR161基因mRNA的高水平表達信號亦在心臟中被檢測到(Leungetal.,2008)。GPR161基因在家雞大腦與心臟組織中的高表達,暗示該基因在家雞中很可能介導與哺乳動物和魚類類似的生理效應,即參與促進神經胚的形成及左右心室的發育等(Leungetal.,2008;Mattesonetal.,2008;Lietal.,2015)。與之相對的是,家雞GPR161基因在腎臟、脂肪、肺中的表達量相對較低,在肝臟、胰腺、皮膚、脾臟中表達信號更弱。本研究結果為闡釋GPR161基因在脊椎動物中的生理效應提供了基礎信息。
Bachmann VA, Mayrhofer JE, Ilouz R,etal. 2016. Gpr161 anchoring of PKA consolidates GPCR and cAMP signaling[J]. Proceedings of the National Academy of Sciences, 113(28): 7786-7791.
Chatr-aryamontri A, Oughtred R, Boucher L,etal. 2016. The BioGRID interaction database: 2017 update[J]. Nucleic Acids Research, 45(D1): D369-D379.
Consortium IHGS. 2004. Finishing the euchromatic sequence of the human genome[J]. Nature, 431(7011): 931-945.
Feigin ME, Xue B, Hammell MC,etal. 2014. G-protein-coupled receptor GPR161 is overexpressed in breast cancer and is a promoter of cell proliferation and invasion[J]. Proceedings of the National Academy of Sciences, 111(11): 4191-4196.
Gainetdinov RR, Premont RT, Bohn LM,etal. 2004. Desensitization of G protein-coupled receptors and neuronal functions[J]. Annual Review of Neuroscience, 27: 107-144.
Karaca E, Buyukkaya R, Pehlivan D,etal. 2014. Whole-exome sequencing identifies homozygous GPR161 mutation in a family with pituitary stalk interruption syndrome[J]. The Journal of Clinical Endocrinology & Metabolism, 100(1): e140-e147. DOI: 10.1210/jc.2014-1984.
Langeberg LK, Scott JD. 2015. Signalling scaffolds and local organization of cellular behaviour[J]. Nature Reviews Molecular Cell Biology, 16(4): 232-244.
Leung T, Humbert JE, Stauffer AM,etal. 2008. The orphan G protein-coupled receptor 161 is required for left-right patterning[J]. Developmental Biology, 323(1): 31-40.
Li BI, Matteson PG, Ababon MF,etal. 2015. The orphan GPCR, Gpr161, regulates the retinoic acid and canonical Wnt pathways during neurulation[J]. Developmental Biology, 402(1): 17-31.
Matteson PG, Desai J, Korstanje R,etal. 2008. The orphan G protein-coupled receptor, Gpr161, encodes the vacuolated lens locus and controls neurulation and lens development[J]. Proceedings of the National Academy of Sciences, 105(6): 2088-2093.
Morris AJ, Malbon CC. 1999. Physiological regulation of G protein-linked signaling[J]. Physiological Reviews, 79(4): 1373-1430.
Mukhopadhyay S, Wen X, Ratti N,etal. 2013. The ciliary G-protein-coupled receptor Gpr161 negatively regulates the sonic hedgehog pathway via cAMP signaling[J]. Cell, 152(1): 210-223.
Pal K, Hwang S, Somatilaka B,etal. 2016. Smoothened determines β-arrestin-mediated removal of the G protein-coupled receptor Gpr161 from the primary cilium[J]. The Journal of Cell Biology, 212(7): 861-875.
Ritter SL, Hall RA. 2009. Fine-tuning of GPCR activity by receptor-interacting proteins[J]. Nature Reviews Molecular Cell Biology, 10(12): 819-830.
Robert X, Gouet P. 2014. Deciphering key features in protein structures with the new ENDscript server[J]. Nucleic Acids Research, 42(W1): W320-W324.
Schmittgen TD, Livak KJ. 2008. Analyzing real-time PCR data by the comparative CT method[J]. Nature Protocols, 3(6): 1101-1108.
Tamura K, Stecher G, Peterson D,etal. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0[J]. Molecular Biology and Evolution, 30(12): 2725-2729.
Wong W, Scott JD. 2004. AKAP signalling complexes: focal points in space and time[J]. Nature Reviews Molecular Cell Biology, 5(12): 959-970.
Zhang Y, DeVries ME, Skolnick J. 2006. Structure modeling of all identified G protein-coupled receptors in the human genome[J]. PLoS Computational Biology, 2(2): e13-e29. DOI: 10.1371/journal.pcbi.0020013.
GeneCloning,MolecularEvolutionandTissueExpressionofGProtein-coupledReceptor161inChicken
ZHU Guoqiang, MO Chunheng, LI Zhengyang, WANG Yajun, LI Juan*
(Key Laboratory of Bio-Resources and Eco-Environment of Ministry of Education, College of Life Sciences,Sichuan University, Chengdu 610065, China)
G protein-coupled receptor 161 (GPR161) is a member of GPCRs, and this orphan receptor plays an important role in the lens development and neurulation of mammals. Recent study showed that the structure of GPR161 was similar with scaffolding proteins and thus may had a potential function in signal transduction. Using chicken as experimental model, the cDNA sequence, molecular evolution and tissue expression of chickenGPR161 gene were investigated in this study. The full-length coding sequence of chickenGPR161 gene was 1 566 bp in length and encoding 521 amino acids. Sequence analysis revealed thatGPR161 of chicken had high sequence similarity with that of human (83.0%), mouse (82.6%) or zebrafish (65.8%). Molecular evolutionary genetical analysis showed that the relationship ofGPR161 among chicken, human and mouse were closer than that of chicken and zebrafish. The result of real-time quantitative PCR showed that the mRNA level ofGPR161 gene was abundant in testis or ovary, brain, heart and muscle. Our data provide a basis for the study of the physiological function ofGPR161 gene in birds.
chicken;GPR161; gene cloning; molecular evolution; tissue expression
10.11984/j.issn.1000-7083.20170140
2017-05-02接受日期2017-07-22
祝國強(1992—), 碩士, 主要從事動物分子遺傳研究, E-mail:872139941@qq.com
*通信作者Corresponding author, E-mail:lijuanscuhk@163.com
Q959.7; Q78
A
1000-7083(2017)06-0632-07
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
