pBabe-GLS1真核表達載體構建及其對結腸癌細胞增殖、谷氨酰胺攝取的影響
2017-12-11 05:23:52李雅莉黃建玲金鑫張波
山東醫藥 2017年41期
關鍵詞:結腸癌
李雅莉,黃建玲,金鑫,張波
(西安交通大學醫學院附屬3201醫院,陜西漢中723000)
pBabe-GLS1真核表達載體構建及其對結腸癌細胞增殖、谷氨酰胺攝取的影響
李雅莉,黃建玲,金鑫,張波
(西安交通大學醫學院附屬3201醫院,陜西漢中723000)
目的構建pBabe-GLS1真核表達載體,觀察谷氨酰胺酶1(GLS1)對結腸癌細胞增殖及谷氨酰胺攝取的影響。方法運用反轉錄PCR擴增GLS1基因片段,通過BamH Ⅰ和Sal Ⅰ雙酶切目的片段并通過T4 DNA連接酶將GLSⅠ基因連接至pBabe真核表達載體中;經BamH Ⅰ和Sal Ⅰ雙酶切法、PCR法測序鑒定重組DNA分子;轉染組將構建的pBabe-GLS1真核表達載體瞬時轉染結直腸癌HT29細胞,陰性對照組轉染空質粒pBabe,采用Western blotting法檢測GLS1蛋白表達,通過谷氨酰胺含量檢測試驗和蛋白定量實驗檢測其對結腸癌細胞的生物學影響。結果成功構建了pBabe-GLS1真核表達載體,且在HT29細胞中高表達GLS1蛋白;轉染組HT29細胞增殖能力高于陰性對照組(Plt;0.05),對谷氨酰胺的攝取能力增加(Plt;0.05)。結論成功構建了pBabe-GLS1真核表達載體,構建的pBabe-GLS1真核表達載體能促進結腸癌細胞增殖及谷氨酰胺攝取。
結腸癌細胞;谷氨酰胺酶1;細胞增殖;谷氨酰胺
葡萄糖和谷氨酰胺是細胞賴以生存的營養物質,對腫瘤來說,其代謝方式的改變是判斷其惡性程度的重要標志之一。在有氧條件下,腫瘤細胞仍大量攝取葡萄糖,且通過糖酵解方式產能,這種有氧糖酵解的代謝特征稱為Warburg效應,亦是腫瘤特有的十大標志特征之一[1,2]。而谷氨酰胺是腫瘤除葡萄糖外的另一重要營養物質。谷氨酰胺酶1(GLS1)是調節谷氨酰胺代謝的關鍵酶之一,負責將谷氨酰胺轉變為谷氨酸,生成的谷氨酸可進一步轉變成α-酮戊二酸進入TCA循環或為谷胱甘肽提供還原當量[3]。2017年2~5月,我們構建了GLS1的過表達載體,并探討其對結腸癌細胞增殖、谷氨酰胺攝取的影響。
1 材料與方法
1.1 材料 高糖DMEM、無酚紅DMEM和PBS購自HyClone公司;人結腸癌細胞系HT29細胞購自中國科學院細胞庫;大腸桿菌DH5α、限制性內切酶BamH Ⅰ和Sal Ⅰ購自Takara公司;pBabe-puro質粒購自Addgene公司;RT-PCR試劑購自Invitrogen公司;GLS1和β-actin一抗購自武漢博士德公司;人抗兔二抗購自CST公司。
1.2 GLS1目的片段的擴增 采用PCR法。從人結腸癌細胞HT29中提取mRNA,然后用 RT-PCR技術擴增GLS1片段。正向引物:3′-GGATCCATGATGCGGCTGCGAGGCT-5′;反向引物:3′-GTCGACTTACAACAATCCATCAAG-5′(下劃線部分為酶切位點)。PCR反應體系:30 μL,變性溫度95 ℃ 10 s,退火溫度55 ℃ 10 s,延伸溫度72 ℃ 30 s,反應30個循壞。將PCR產物通過瓊脂糖凝膠電泳分離,在2 kb左右的位置觀察到特異性條帶,將其回收酶切,并經測序鑒定。
1.3 pBabe-GLS1過表達載體的構建 將上述2 kb左右的片段利用BamH Ⅰ和Sal Ⅰ進行酶切,酶切回收后,利用T4 DNA連接酶將目的片段連接至pBabe真核表達載體。將此連接產物轉化至感受態DH5α,挑取陽性克隆,通過BamH Ⅰ和Sal Ⅰ雙酶切鑒定。
1.4 pBabe-GLS1轉染HT29細胞 將2×105個細胞接種于6孔板,待貼壁后長至60%密度時進行轉染:pBabe-GLS1質粒2 μg、脂質體5 μL,將兩者與無血清opti-MEM混勻后加至細胞培養液中,轉染6 h后換成含10% FBS的DMEM培養48 h。
1.5 GLS1蛋白表達檢測 采用Western blotting法。收集轉染pBabe-GLS1的HT29細胞,加入RIPA蛋白裂解液,超聲裂解,4 ℃離心取上清液,收集總蛋白。經10% SDS-PAGE膠分離后,電轉(200 mA、2 h)至NC膜上。利用含5 %脫脂奶粉的TBST封閉NC膜;2 h后,用1∶1 000稀釋的GLS1抗體4 ℃孵育過夜,次日加入二抗孵育1 h,通過ECL發光檢測劑顯示結果。
1.6 細胞活力檢測 采用MTT法。將結腸癌細胞HT29分為轉染組與陰性對照組,分別轉染pBabe-GLS1、空質粒pBabe,24 h后,將各組細胞用含10%血清的培養液制成單個細胞懸液,以每孔104個細胞接種到96孔板;分別于培養0、1、2、3 d時進行MTT實驗:每孔加入MTT溶液至終濃度為5 mg/mL,孵育4 h后通過DMSO溶解結晶物;酶聯免疫檢測儀于490 nm波長處測定各孔光密度(OD)值,并繪制細胞生長曲線。
1.7 谷氨酰胺檢測 將結腸癌細胞HT29分為轉染組與陰性對照組,分別轉染pBabe-GLS1、空質粒pBabe,分別按照5×105細胞/孔接種于6孔板。待細胞貼壁后,棄培養基,更換為無血清無酚紅DMEM,繼續培養24 h。收集培養上清。用細胞刮收集細胞,溶于200 μL的PBS溶液,超聲裂解后取細胞裂解產物。通過BCA法確定蛋白濃度。通過NH3來計算谷氨酰胺的量。根據試劑盒說明書,得到相應的數值,以無細胞對照孔培養液檢測值為初始濃度,通過計算獲得谷氨酰胺含量。即細胞攝取的谷氨酰胺=無酚紅DMEM中谷氨酰胺初始濃度-培養上清中谷氨酰胺濃度。
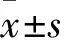
2 結果
2.1 構建pBabe-GLS1真核表達載體的鑒定 成功得到GLS1基因片段,經瓊脂糖凝膠電泳證實片段大小約為2 kb(圖1)。回收該片段,并用BamH Ⅰ和Sal Ⅰ雙酶切,發現1號和3號克隆可以切得一條2 kb大小的片段,說明其含有GLS1目的片段,經測序后鑒定正確。

圖1 GLS1片段10%瓊脂糖凝膠電泳圖
2.2 轉染pBabe-GLS1表達的鑒定 Western blotting實驗檢測發現,陰性對照組HT29細胞中GLS1表達相對較低,而轉染組GLS1蛋白呈高表達(圖2),證實構建的重組質粒pBabe-GLS1能在真核細胞中正確表達。
2.3 GLS1的過表達對細胞增殖能力及對谷氨酰胺攝取的影響 描繪的細胞生長曲線見圖3,轉染組HT29細胞增殖能力增加,與陰性對照組相比,Plt;0.05;轉染組、陰性對照組HT29細胞谷氨酰胺攝取量分別為6.902、4.139 mmol/g蛋白,兩組相比,Plt;0.05。

圖2 Western blotting鑒定GLS1在HT29細胞中表達

圖3 MMT法描繪細胞生長曲線
3 討論
谷氨酰胺是腫瘤細胞利用并依賴的重要能源物質,其作用包括:為細胞增殖提供足夠的碳源、氮源和ATP[4]、為缺少葡萄糖來源的線粒體TCA循環的維持提供補給[5]、為細胞內抗氧化系統的運行提供谷胱甘肽等還原當量[6]。研究[7]發現,通過有氧酵解方式代謝的腫瘤細胞相對正常細胞更依賴谷氨酰胺的攝入。主要原因:一方面,高度惡性的腫瘤細胞多伴隨重要癌基因的活化,如Myc、HIF-1α等,通過調節谷氨酰胺代謝的關鍵酶表達和活性,促進谷氨酰胺的攝入;另一方面,有氧酵解的腫瘤細胞增殖旺盛,通過利用谷氨酰胺分解產生的ATP、核酸、氨基酸等物質,滿足其自身生長需要。此外,谷氨酰胺還能調節細胞內重要癌基因和信號通路,如PI3K/Akt、mTOR等,促進腫瘤的惡性演進過程[8]。
GLS主要有兩種亞型,GLS1和GLS2。其中GLS1在調節谷氨酰胺代謝中扮演重要角色,通過將谷氨酰胺轉變為谷氨酸,生成的谷氨酸可進一步轉變成α-酮戊二酸進入TCA循環或為谷胱甘肽提供還原當量[3]。研究[9,10]發現,GLS1在多類腫瘤組織中呈高表達,如乳腺癌、前列腺癌、白血病等。基于代謝重編程與腫瘤惡性程度的相關性,以調節代謝的關鍵分子為靶點的腫瘤診斷和治療工作一直是研究熱點。PET-CT技術便是利用腫瘤細胞大量攝取葡萄糖的特點,通過18F等放射性核素標記葡萄糖分子進行顯像,目前已廣泛用于各類腫瘤的診斷和實驗室研究工作中[11]。在治療領域,諸多基于GLS1的抑制劑正處于實驗室研究或臨床試驗階段,如BPTES,CB-839等,并取得了令人滿意的效果[12,13]。
本研究中,我們成功構建了pBabe-GLS1真核表達質粒,通過在結腸癌細胞HT29中驗證其正確高表達后,利用谷氨酰胺檢測實驗證明GLS1的高表達可促進HT29細胞增殖及其對谷氨酰胺的攝取,為后續研究GLS1在腫瘤代謝中的作用及相關機制奠定了基礎。
[1] Wu W, Zhao S. Metabolic changes in cancer: beyond the Warburg effect[J]. Acta Biochim Biophys Sin (Shanghai), 2013,45(1):18-26.
[2] Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation[J]. Cell, 2011,144(5):646-674.
[3] Jones NP, Schulze A. Targeting cancer metabolism--aiming at a tumour′s sweet-spot[J]. Drug Discov Today, 2012,17(5-6):232-241.
[4] Reitzer LJ, Wice BM, Kennell D. Evidence that glutamine, not sugar, is the major energy source for cultured HeLa cells[J]. J Biol Chem, 1979,254(8):2669-2676.
[5] Le A, Lane AN, Hamaker M, et al. Glucose-independent glutamine metabolism via TCA cycling for proliferation and survival in B cells[J]. Cell Metab, 2012,15(1):110-121.
[6] Curthoys NP, Watford M. Regulation of glutaminase activity and glutamine metabolism[J]. Annu Rev Nutr, 1995,15(1):133-159.
[7] Lu W, Pelicano H, Huang P. Cancer metabolism: is glutamine sweeter than glucose[J]. Cancer Cell, 2010,18(3):199-200.
[8] Hosios AM, Vander Heiden MG. Acetate metabolism in cancer cells[J]. Cancer Metab, 2014,2(1):27.
[9] Pan T, Gao L, Wu G, et al. Elevated expression of glutaminase confers glucose utilization via glutaminolysis in prostate cancer[J]. Biochem Biophys Res Commun, 2015,456(1):452-458.
[10] Qie S, Chu C, Li W, et al. ErbB2 activation upregulates glutaminase 1 expression which promotes breast cancer cell proliferation[J]. J Cell Biochem, 2014,115(3):498-509.
[11] Garcia Vicente AM, Soriano Castrejon A, Cruz Mora MA, et al. Dual time point 2-deoxy-2-[18F]fluoro-D-glucose PET/CT: nodal staging in locally advanced breast cancer[J]. Rev Esp Med Nucl Imagen Mol, 2014,33(1):1-5.
[12] Gross MI, Demo SD, Dennison JB, et al. Antitumor activity of the glutaminase inhibitor CB-839 in triple-negative breast cancer[J]. Mol Cancer Ther, 2014,13(4):890-901.
[13] Elgogary A, Xu Q, Poore B, et al. Combination therapy with BPTES nanoparticles and metformin targets the metabolic heterogeneity of pancreatic cancer[J]. Proc Natl Acad Sci U S A, 2016,113(36):E5328-E5336.
10.3969/j.issn.1002-266X.2017.41.010
R735.35
A
1002-266X(2017)41-0033-03
2017-03-14)
猜你喜歡
天津醫科大學學報(2019年6期)2019-08-13 07:04:32
西南國防醫藥(2016年7期)2016-12-01 06:01:15
腹腔鏡外科雜志(2016年10期)2016-06-01 12:10:08
中國衛生標準管理(2015年6期)2016-01-14 05:17:12
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
中國當代醫藥(2015年26期)2015-03-01 02:06:57
西南軍醫(2015年6期)2015-01-23 01:25:50
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
中華介入放射學電子雜志(2014年1期)2014-02-02 05:24:06
