茶樹MADS-box轉錄因子基因的克隆與非生物脅迫響應分析
2017-12-11 10:08:08沈威滕瑞敏李輝劉志薇王永鑫王文麗莊靜
茶葉科學 2017年6期
沈威,滕瑞敏,李輝,劉志薇,王永鑫,王文麗,莊靜
南京農業大學園藝學院,茶葉科學研究所,江蘇 南京 210095
茶樹MADS-box轉錄因子基因的克隆與非生物脅迫響應分析
沈威,滕瑞敏,李輝,劉志薇,王永鑫,王文麗,莊靜*
南京農業大學園藝學院,茶葉科學研究所,江蘇 南京 210095
本研究基于茶樹轉錄組數據庫,以茶樹龍井43為試驗材料,通過RT-PCR方法從該茶樹的cDNA中克隆得到1個CsMADS1基因。序列分析表明:茶樹CsMADS1基因開放閱讀框長度為657 bp,編碼218個氨基酸,是典型的植物MADS-box家族轉錄因子。序列多重比對顯示,該序列與多個相關物種的MADS-box序列一致性為65.65%,含有高度保守的MADS結構域和半保守的K結構域。氨基酸理化性質、親疏水性、亞細胞定位預測、無序化分析,以及二級和三級結構分析顯示,CsMADS1轉錄因子是親水性蛋白,可能定位于細胞核中,無序化程度明顯,以 α-螺旋結構為主,并與人 MEF2蛋白具有相似的三級結構。利用實時熒光定量PCR方法分析了茶樹龍井43中CsMADS1基因在非生物脅迫下的表達。結果表明,茶樹中CsMADS1基因對高溫、低溫、干旱和高鹽等不同非生物脅迫有響應,且表達存在差異。
茶樹;MADS-box;轉錄因子;進化分析;非生物脅迫;表達分析
MADS-box基因廣泛存在于植物中,它所編碼的蛋白是一類轉錄因子,由于在植物生長發育方面發揮著非常重要的作用而被廣泛研究。在擬南芥(Arabidopsis thaliana)[1]、水稻(Oryza sativa)[2]、葡萄(Vitis vinifera)[3]、楊樹(Populus)[4]、二穗短柄草(Brachypodium distachyon)[5]、和蕪菁(Brassica rapa)[6]中,MADS-box家族轉錄因子分別有107、75、93、105、57和167個成員。
目前,MADS-box基因研究的熱點主要集中在植物花發育過程中,該基因在ABCDE模型中發揮重要的調控作用[7]。除此之外,MADS-box基因還在植物的根發育[8]、芽休眠[9]、果實成熟[10]及細胞衰老[11]等方面起著重要作用。MADS-box基因在植物逆境響應等方面也起著重要作用,例如:Cooper等建立的基因網絡證實了 MADS-box轉錄因子參與水稻的逆境調控[12];矮牽牛中的PMADS9基因在多種逆境脅迫條件下表達量均顯著提高[13];Yu等[14]發現在多種逆境處理下,MADS-box基因的表達顯著影響植物根系的生長與發育。但是,目前該基因在茶樹逆境方面的研究還鮮有報道。茶樹(Camellia sinensis)屬于山茶科山茶屬,是一種喜酸懼堿,喜溫怕寒的多年生常綠木本植物,在我國已經有幾千年的栽培歷史[15]。茶樹一生中受到多種非生物脅迫傷害,其中高溫、低溫、干旱、高鹽等嚴重影響著茶樹的生長發育,從而影響茶葉的產量和品質。因此,研究 MADS-box轉錄因子在茶樹生長發育和逆境調控中的作用具有重要意義。
本研究以茶樹材料龍井43的cDNA為模板克隆得到1個茶樹CsMADS1基因,并對該基因編碼的氨基酸序列、理化性質、進化樹和三級結構等方面進行了分析。利用熒光定量PCR,分析了龍井43中CsMADS1基因在高溫、低溫、干旱和高鹽等4種非生物脅迫條件下的表達特征。本研究以期通過生物技術,探尋與茶樹逆境響應相關的基因,為茶樹抗逆遺傳育種和生長發育調控提供理論基礎。
1 材料與方法
1.1 試驗材料和脅迫處理
試驗材料為茶樹龍井 43兩年生扦插幼苗,種植于南京農業大學茶葉科學研究所溫室。摘取健康茶樹植株的幼嫩葉片,進行總RNA提取并反轉錄為cDNA。對龍井43兩年生植株進行 4種非生物脅迫處理,分別為:38℃高溫、4℃低溫、20% PEG、200 mmol·L-1NaCl,處理時間分別為1、4、8、24 h;以未做任何處理的龍井43植株葉片為空白對照,每個處理設3個重復。其中溫度處理由光照培養箱控制,分別設為38℃、4℃,濕度設為70%;干旱和鹽處理采用葉面噴施的方式,分別在各時間段采集不同處理的第2葉,保存在-80℃冰箱,于第2天統一提取RNA后合成cDNA,作為實時熒光定量檢測的模板。
1.2 茶樹總RNA的提取和cDNA的合成
茶樹龍井43的RNA提取參考Quick RNA Isolation Kit試劑盒說明書完成(北京華越洋生物科技有限公司),RNA的樣品濃度和質量分別用微量紫外檢測儀Nano-Drop和1.2%瓊脂糖凝膠電泳檢測。RNA反轉錄成 cDNA按照Prime Script RT Reagent Kit試劑盒(大連TaKaRa公司)完成。
1.3 茶樹CsMADS1基因的克隆
根據本實驗室茶樹轉錄組數據[16],設計1對引物(CsMADS1-F:5′-ATGGTGAGACA GAGAATTCAGATC-3′和 CsMADS1-R:5′-GC TCTGATAAGGTAGGCCCAA-3′) 。 以 龍 井43葉片cDNA為模板進行PCR擴增。擴增體系為 20 μL:CsMADS1-F/R 引物各 1 μL、10 μLEx TaqMix 酶、1 μL 模板、7 μL ddH2O。PCR擴增程序為:94℃、5 min,94℃、30 s,52℃、30 s,72℃、1 min,35個循環;72℃,10 min。根據DNA回收試劑盒將經過1.2%的瓊脂糖凝膠電泳分離的 PCR產物回收,隨后連接到pMD19-T載體上,轉化至大腸桿菌DH5α。質粒酶切和鑒定之后送至通用生物系統(安徽)有限公司測序。
1.4 生物信息學分析
通過BioXM2.6軟件查找基因的開放閱讀框以及編碼氨基酸序列。在 NCBI網站(http://www.ncbi.nlm.nih.gov)對氨基酸序列進行同源性搜索和保守域預測。利用ExPASy-ProtParam(http://web.expasy.org/prot-param)和SMS(http://www.bio-soft.net/sms)分析相關物種MADS-box轉錄因子的氨基酸理化性質、理論等電點及組成成分等。使用最新版本MEGA 7軟件構建分子系統發育進化樹[17]。借助于 DNAMAN 8.0軟件進行茶樹CsMADS1基因推導氨基酸的多重對比和親疏水性分析。通過 LocTree3(https://rostlab.org/services/loctree3)和SoftBerry ProtComp 9.0(http://linux1. softberry.com/berry.phtml)完成亞細胞定位預測[18-19]。無序化特征分析利用Fold Index[20]程序(http://bip.weizmann.ac.il/fldbin/findex)完成。利用SOPMA[21]程序(http://npsa-pbil.
ibcp.fr/cgi-bin/npsa)和Swiss-Model[22]程序(https://swissmodel.expasy.org)進行蛋白質二級結構及三級結構預測分析,并在Swiss-Pdb Viewer軟件上編輯生成茶樹CsMADS1轉錄因子的三維結構圖形。
1.5 茶樹CsMADS1基因的表達特性分析
按照SYBR PremixEx Taq試劑盒(大連TaKaRa公司)操作說明,使用 Bio-CFX96 real-time PCR system儀器完成實時熒光定量PCR。茶樹GAPDH基因作為內參基因[23],與目標基因CsMADS1同時擴增。擴增體系為15 μL:酶 7.5 μL,模板 1.5 μL,上下游引物各 0.3 μL,ddH2O 5.4 μL。擴增程序為:95℃ 5 min;95℃5 s,55℃ 30 s,65℃ 10 s,40個循環。內參基因和目標基因CsMADS1的表達檢測引物分別 為 : 5′-TTGGCATCGTTGAGGGTCT-3′和5′-CAGTGGGAACACGGAAAGC-3′;5′-GGC ATAATCTACAACCACAG-3′和 5′-CCACGC ATTCTCTTCCAT-3′。用2-ΔΔCT法計算分析茶樹龍井43中CsMADS1相對表達量[24]。
2 結果與分析
2.1 龍井43茶樹中CsMADS1基因的克隆
以龍井43茶樹材料葉片的cDNA為模板,利用引物 CsMADS1-F和 CsMADS1-R進行PCR擴增反應,對擴增片段進行克隆、測序。將該序列在NCBI上進行Blast分析,結果顯示該序列與多種植物的MADS-box基因有較高同源性,為所需的目的序列。通過BioXM2.6軟件分析顯示,該基因開放閱讀框長度為657 bp,編碼 218個氨基酸(圖 1),命名為CsMADS1。

圖1 茶樹CsMADS1基因核苷酸序列與推測的氨基酸序列Fig. 1 The nucleotide and amino acid sequences of CsMADS1 gene from C. sinensis
2.2 茶樹CsMADS1基因編碼氨基酸序列比對及理化性質分析
對茶樹CsMADS1基因編碼氨基酸序列進行分析,結果顯示(圖2-A),該氨基酸序列在3~60位含有高度保守的MADS結構功能域,在94~169位含有半保守的結構功能域K-box,具有植物MADS轉錄因子家族典型的MIKC結構域。為進一步分析茶樹 CsMADS1的保守結構域,將它與其他相似度較高的多個物種MADS-box氨基酸序列進行多重比對(圖2-B),結果顯示一致性為65.65%,它們均含有高度保守的MADS結構域和半保守的K結構域。
對上述植物 MADS-box轉錄因子蛋白氨基酸進行理化性質分析。結果顯示(表1),這些植物中MADS-box蛋白殘基數在210~241之間,相對分子質量在23.72~27.26 kDa之間;酸性氨基酸比例略少于堿性氨基酸,理論等電點在 5.75~8.96之間。芳香族氨基酸占 4%左右,明顯少于脂肪族氨基酸;總平均疏水性在-0.7左右,表現出較強的親水性。

圖2 茶樹CsMADS1轉錄因子保守域及與其他植物氨基酸序列的多重比對Fig. 2 The conserved domain of CsMADS1 transcription factor and alignment of amino acid sequences from C. sinensis and other plant species

表1 不同植物中MADS-box家族轉錄因子氨基酸組成成分及理化性質分析Table 1Comparison of compositionand physicochemical properties of aminoacidsequences of MADS-box transcription factors amongplant species

表2 CsMADS1亞細胞定位預測Table 2The predictedsubcellular localization of CsMADS1
2.3 茶樹CsMADS1氨基酸序列進化樹分析
為了進一步分析茶樹 CsMADS1與MADS-box家族相關轉錄因子之間的進化關系,參考 Annette Becker等[25]的 MADS-box轉錄因子家族分類方法,并利用MEGA7構建茶樹CsMADS1與擬南芥MADS-box家族轉錄因子的同源進化樹(圖3),結果顯示,茶樹CsMADS1轉錄因子屬于MADS-box家族轉錄因子中的STMADS11亞族。
2.4 茶樹CsMADS1蛋白氨基酸親疏水性分析
利用DNAMAN 8.0軟件對茶樹CsMADS1轉錄因子進行親疏水性分析。結果顯示,茶樹CsMADS1轉錄因子親水性最強的位點是第155位的亮氨酸(Leu)(圖4-A);疏水性最強的位點是45位的纈氨酸(Val)(圖4-B)。由此可知,親水性氨基酸明顯占更多比例,推測CsMADS1轉錄因子屬于親水性蛋白。
2.5 茶樹CsMADS1蛋白亞細胞定位預測
通過在線預測軟件 SoftBerry ProtComp 9.0對CsMADS1蛋白進行亞細胞定位預測。結果顯示(表2),茶樹CsMADS1蛋白定位在細胞核的相應預測數值最高。進一步利用在線預測軟件LOCTREE3對CsMADS1蛋白進行亞細胞定位,結果顯示CsMADS1蛋白定位在細胞核。由此判斷,CsMADS1蛋白可能定位于細胞核中。
2.6 茶樹CsMADS1蛋白固有無序化預測與分析
利用FoldIndex程序對CsMADS1氨基酸序列進行了無序化分析。結果表明(圖5),茶樹CsMADS1氨基酸序列中無序化區域有2個,總的無序化氨基酸數目為143,無序化比例為 65.60%。所以 CsMADS1有明顯的無序化程度。
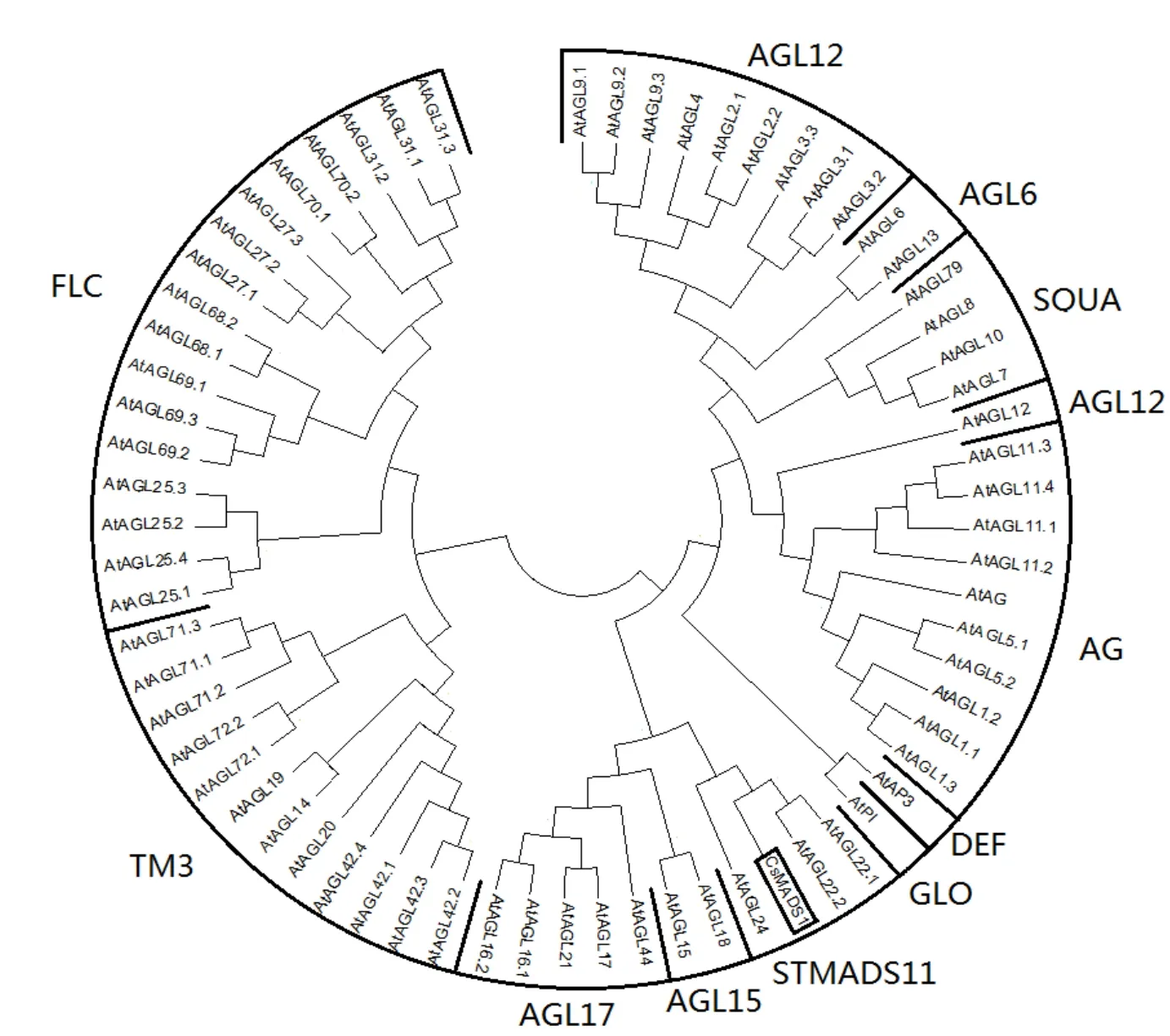
圖3 茶樹CsMADS1轉錄因子的進化分析Fig. 3 Phylogenetic analysis of CsMADS1 transcription factors from C. sinensis

圖4 茶樹CsMADS1氨基酸序列親疏水性分析Fig. 4 Analysis of hydrophilicity and hydrophobicity of CsMADS1 from C. sinensis
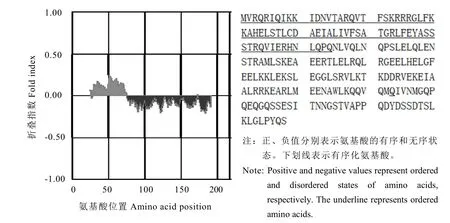
圖5 茶樹CsMADS1折疊狀態的分析Fig. 5 Analysis of the folding state of CsMADS1 from C. sinensis
2.7 茶樹CsMADS1蛋白推導的二級結構和三級結構預測分析
從茶樹 CsMADS1蛋白序列預測的二級結構中可以看出(圖 6),CsMADS1蛋白由53.21%的 α-螺旋(Alpha helix),14.68%的延伸主鏈(Extended strand),6.42%的 β-轉角(Beta turn)和25.69%的隨機卷曲(Random coil)組成。其中α-螺旋結構是CsMADS1蛋白的主要組成部分,而延伸主鏈和隨機卷曲結構則散布于蛋白序列中。
以人 MEF2轉錄因子為模板,對茶樹CsMADS1轉錄因子的三級結構進行建模分析,結果顯示相似度為48.65%。在結合域的α螺旋、β-折疊部位,CsMADS1與MEF2的差異位點共有19個,其中第1個和第2個α-螺旋分別有9個和8個差異位點,另外2個差異位點分別在兩個β-折疊部位(圖7),但這些差異位點對其三級結構并沒有太大影響。
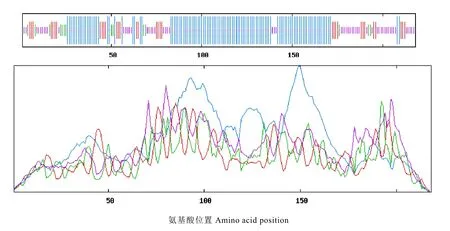
圖6 茶樹CsMADS1的二級結構預測Fig. 6 The secondary structure of CsMADS1 from C. sinensis

圖7 茶樹CsMADS1的三維結構預測Fig. 7 The tertiary structure of CsMADS1 from C. sinensis
2.8 茶樹CsMADS1基因在不同非生物脅迫下的表達分析
通過實時熒光定量 PCR方法,檢測了龍井43茶樹中CsMADS1基因在高溫(38℃)、低溫(4℃)、干旱(20% PEG)和高鹽(200 mmol/L NaCl)4種非生物脅迫條件下的表達情況。由圖8可知,茶樹中CsMADS1基因的表達在不同脅迫時間和不同脅迫處理間存在差異。在高溫、干旱和高鹽脅迫處理下,CsMADS1表達量均表現出先降低后增加再降低的趨勢,其中高溫和干旱脅迫下的表達量均在8 h時達到了最大值,且較對照差異顯著,分別為對照的1.34和2.22倍。在高鹽脅迫下的表達量較對照均表現出一定程度下調,1 h時表達量最低,隨處理時間增加,到8 h時表達量有一定的回升,此時與對照無顯著差異。低溫脅迫處理下,CsMADS1表達量先增加后降低再增加,在1 h和4 h時的CsMADS1表達量顯著高于對照,分別為對照的2.29和1.88倍,而在8 h和24 h時CsMADS1較對照均無顯著差異。
3 討論
植物生長發育過程中常受到高低溫、干旱、高鹽等非生物脅迫的傷害,嚴重時會導致植株死亡。茶樹是極易受到惡劣環境影響的經濟作物之一。目前,關于 MADS-box轉錄因子對茶樹在生長發育和逆境調控中的作用機制等研究尚未見報導。因此本文開展茶樹中MADS-box轉錄因子的克隆、序列分析、結構特征、功能及特性等研究,及其對茶樹逆境脅迫的響應,具有較為重要的意義。
本文通過RT-PCR方法從茶樹龍井43的cDNA中克隆得到1個MADS-box基因,命名為CsMADS1,該基因編碼218個氨基酸,通過Blast分析表明,它具有MADS-box轉錄因子家族典型的 MIKC結構域[26]。序列比對和系統進化樹分析結果表明,茶樹CsMADS1轉錄因子屬于MADS-box家族的STMADS11亞族,與多種植物有較高的同源性。亞細胞定位預測顯示,CsMADS1定位于細胞核中。其無序化比例為65.60%,可能與其親水性氨基酸比例較高有關[20]。茶樹CsMADS1二級結構主要表現為α-螺旋結構。通過三級結構建模顯示,茶樹CsMADS1雖與人MEF2蛋白有19個差異位點,但對空間結構影響較小[27]。
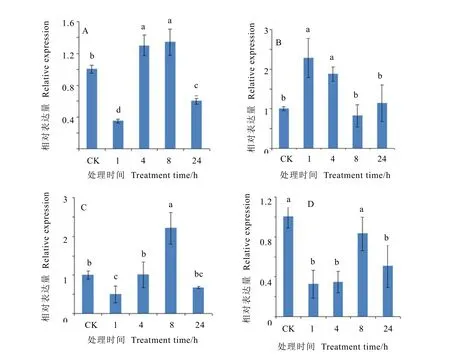
圖8 茶樹CsMADS1在不同處理條件下的表達Fig. 8 Expression patterns of CsMADS1 under different stress treatments
實時熒光定量 PCR分析表明,高低溫、干旱、高鹽等非生物脅迫明顯影響著茶樹CsMADS1轉錄因子的表達量,但不同逆境之間響應存在一定的差異。已有的研究證明,在二穗短柄草中,多個MADS-box基因在低溫、干旱和鹽脅迫處理后出現不同的響應[5];銀杏葉在鹽、干旱和冷脅迫后,MADS-box轉錄因子基因的表達量顯著高于對照[28];Guo等[29]發現,番茄SlMBP11基因在鹽和受傷處理下的表達較為相似,都是先上升再下降,表達量分別在4 h和8 h時達到最大值,在干旱處理時,是一個逐漸上升的趨勢,直到 36 h時到達最大值;水稻[2,30]和大白菜[31]在經過鹽、干旱、低溫、GA3和BAP等處理后,MADS-box轉錄因子基因的表達量也出現不同程度的上調和下調。本試驗中,茶樹CsMADS1基因受低溫脅迫后能夠快速誘導表達,在鹽脅迫下主要表現為被抑制,在高溫和干旱脅迫下,其表達量先降低,隨后逐漸上升后又下降。以上結果顯示,MADS-box轉錄因子在不同物種間的響應有明顯的差異性。
MADS-box轉錄因子擁有范圍廣泛的靶目標基因,它的家族成員均是通過 C區的保守序列與其他因子形成復合體而行使功能的。對于高等植物而言,任一逆境環境的改變不僅僅導致植株簡單的生理變化,而是通過影響其他因子的響應來影響 MADS蛋白復合體的功能[13]。Hill等[32]證明與 MADS-box亞家族成員 AGL15作用的 SIN3/HDAC1復合體成員SAP18在低溫、干旱和高鹽脅迫下,其表達量將會增加。由于 MADS-box轉錄因子自身擁有靶目標的廣泛性和復合體多樣性等特點,同時不同物種間在生理途徑和響應逆境具有差異性,CsMADS1基因在響應茶樹逆境脅迫時也表現出特有的表達模式。CsMADS1基因在茶樹中可能作為調控基因直接參與抗逆相關生理途徑,或者通過調控抗逆相關調控因子間接影響茶樹的生長發育。由于植物體內逆境調控的復雜性,以上基于生物信息學分析的預測結果有待更多的試驗研究加以驗證,以便進一步了解茶樹 CsMADS1轉錄因子的調節途徑和作用機理。
[1]Parenicová L, De F S, Kieffer M, et al. Molecular and phylogenetic analyses of the complete MADS-box transcription factor family in Arabidopsis: new openings to the MADS world [J]. Plant Cell, 2003, 15(7): 1538-1551.
[2]Arora R, Agarwal P, Ray S, et al. MADS-box gene family in rice: genome-wide identification, organization and expression profiling during reproductive development and stress [J]. BMC Genomics, 2007, 8(1): 242-263.
[3]Díazriquelme J, Lijavetzky D, Martínezzapater J M, et al.Genome-Wide analysis of MIKCC-type mads box genes in grapevine [J]. Plant Physiology, 2009, 149(1): 354-369.
[4]Leseberg C H, Li A, Kang H, et al. Genome-wide analysis of the MADS-box gene family inPopulus trichocarpa[J].Gene, 2006, 378(1): 84-94.
[5]Wei B, Zhang R Z, Guo J J, et al. Genome-wide analysis of the MADS-Box gene family inBrachypodium distachyon[J].Plos One, 2014, 9(1): e84781.
[6]Rameneni J J, Dhandapani V, Paul P, et al. Genome-wide identification, characterization, and comparative phylogeny analysis of MADS-box transcription factors inBrassica rapa[J]. Genes amp; Genomics, 2014, 36(4): 509-525.
[7]趙興富, 朱永平, 肖靖譯, 等. 植物 MADS-box基因多樣性及進化研究進展[J]. 北方園藝, 2015(11): 180-186.
[8]Yu C, Liu Y, Zhang A, et al. MADS-box Transcription factor OsMADS25 regulates root development through affection of nitrate accumulation in rice [J]. Plos One, 2015,10(8): e0135196.
[9]Yamane H, Tao R, Ooka T. Comparative analyses of dormancy-associated MADS-box genes, PpDAM5 and PpDAM6, in low- and high-chill peaches (PrunuspersicaL.)[J]. Journal of the Japanese Society for Horticultural Science,2011, 80(3): 276-283.
[10]Shima Y, Kitagawa M, Fujisawa M, et al. Tomato FRUITFULL homologues act in fruit ripening via forming MADS-box transcription factor complexes with RIN [J].Plant Molecular Biology, 2013, 82(4): 427-438.
[11]Xie Q, Hu Z, Zhu Z, et al. Overexpression of a novel MADS-box geneSlFYFLdelays senescence, fruit ripening and abscission in tomato [J]. Scientific Reports, 2014,4(7491): 4367. DOI: 10.1038/srep04367.
[12]Cooper B, Clarke J D, Budworth P, et al. A network of rice genes associated with stress response and seed development[J]. Proceedings of the National Academy of Sciences, 2003,100(8): 4945-4950.
[13]陳璟, 李名揚, 閆明旭, 等. 矮牽牛 PMADS9基因的結構特征和 mRNA的表達分析[J]. 園藝學報, 2011, 38(1):108-116.
[14]Yu L H, Miao Z Q, Qi G F, et al. MADS-box transcription factor AGL21 regulates lateral root development and responds to multiple external and physiological signals [J].Molecular Plant, 2014, 7(11): 1653-1669.
[15]童啟慶. 茶樹栽培學[M]. 北京: 中國農業出版社, 2007:4-33.
[16]Wu Z J, Li X H, Liu Z W, et al. De novo assembly and transcriptome characterization: Novel insights into catechins biosynthesis inCamelliasinensis[J]. BMC Plant Biology,2014, 14(1): 277. DOI: 10.1186/s12870-014-0277-4.
[17]Kumar S, Stecher G, Tamura K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular Biology amp; Evolution, 2016, 33(7):1870-1874.
[18]Goldberg T, Hecht M, Hamp T, et al. LocTree3 prediction of localization [J]. Nucleic Acids Research, 2014, 42(Web Server issue): W350-W355.
[19]李慧峰, 賈厚振, 董慶龍, 等. ‘魯星’桃中 10個MADS-box基因克隆和表達分析[J]. 中國農業科學, 2016(23):4593-4605.
[20]Prilusky J, Felder C E, Zeevbenmordehai T, et al. Foldindex:a simple tool to predict whether a given protein sequence is intrinsically unfolded [J]. Bioinformatics, 2005, 21(16):3435-3438.
[21]Combet C, Blanchet C, Geourjon C, et al. NPS@: network protein sequence analysis [J]. Trends in Biochemical Sciences, 2000, 25(3): 147-150.
[22]Schwede T, Kopp J, Guex N, et al. SWISS-MODEL: an automated protein homology-modeling server [J]. Nucleic Acids Research, 2003, 31(13): 3381-3385.
[23]Wu Z J, Tian C, Jiang Q, et al. Selection of suitable reference genes for qRT-PCR normalization during leaf development and hormonal stimuli in tea plant (Camellia sinensis) [J]. Scientific Reports, 2016, 6: 19748. DOI:10.1038/srep19748.
[24]Pfaffl M W. A new mathematical model for relative quantification in real-time RT-PCR [J]. Nucleic Acids Research, 2001, 29(9): e45.
[25]Becker A, Theissen G. The major clades of MADS-box genes and their role in the development and evolution of flowering plants [J]. Molecular Phylogenetics amp; Evolution,2004, 29(3): 464-489.
[26]Ren Z Y, Yu D Q, Yang Z E, et al. Genome-wide identification of the MIKC-type MADS-box gene family inGossypiumhirsutumL. unravels their roles in flowering [J].Frontiers in Plant Science, 2017, 8: 384.https://doi.org/10.3389/fpls.2017.00384.
[27]Han A, Pan F, Stroud J C, et al. Sequence-specific recruitment of transcriptional co-repressor cabin1 by myocyte enhancer factor-2 [J]. Nature, 2003, 422(6933):730-734.
[28]Yang F, Xu F, Wang X, et al. Characterization and functional analysis of a MADS-box transcription factor gene(GbMADS9) fromGinkgobiloba[J]. Scientia Horticulturae,2016, 212: 104-114.
[29]Guo X, Chen G, Cui B, et al.Solanum lycopersicum,agamous-like MADS-box protein AGL15-like gene,SlMBP11,confers salt stress tolerance [J]. Molecular Breeding, 2016,36(9): 125. DOI: 10.1007/s11032-016-0544-1.
[30]Puig J, Meynard D, Khong G N, et al. Analysis of the expression of the AGL17 - like, clade of MADS-box transcription factors in rice [J]. Gene Expression Patterns,2013, 13(5): 160-170.
[31]Duan W, Song X, Liu T, et al. Genome-wide analysis of the MADS-box gene family inBrassicarapa(Chinese cabbage)[J]. Molecular Genetics and Genomics, 2015, 290(1):239-255.
[32]Hill K, Wang H, Perry S. A transcriptional repression motif in the MADS factor AGL15 is involved in recruitment of histone deacetylase complex components [J]. Plant Journal for Cell amp; Molecular Biology, 2008, 53(1): 172-185.
Cloning of a MADS-box Transcription Factor Gene from Camellia sinensis and its Response to Abiotic Stresses
SHEN Wei, TENG Ruimin, LI Hui, LIU Zhiwei, WANG Yongxin, WANG Wenli, ZHUANG Jing*
Tea Science Research Institute, College of Horticulture, Nanjing Agricultural University, Nanjing 210095, China
In this study, based on the transcriptome database of tea plant, theCsMADS1gene was cloned from cDNA of ‘Longjing43’ by RT-PCR method. The length of open reading frame ofCsMADS1gene was 657 bp, encoding 218 amino acids, which was a typical transcription factor of MADS-box family. Multiple alignments of CsMADS1 with related species showed that the identity of them was 65.65%, with a highly conserved MADS domain and a semi-conserved K domain. The physicochemical properties, hydrophilicity and hydrophobicity, subcellular localization, disordered feature, secondary and tertiary structure were also analyzed. CsMADS1 transcription factor is a hydrophilic protein, may be located in nucleus. The disordered feature of CsMADS1 protein was obvious, which was mainly composed of alpha helix structure, and had similar tertiary structure with MEF2 of human. Quantitative real-time PCR was used to analyze the expression profiles of theCsMADS1gene under abiotic stress treatments of tea cultivar ‘Longjing43’. The results showed that theCsMADS1gene responded to high and low temperatures, drought and high salinity. Different expression patterns ofCsMADS1gene were observed under those abiotic stress treatments.
Camellia sinensis, MADS-box, transcription factor, phylogenetic analysis, abiotic stress, expression analysis
TS272.5+4;Q946.88
A
1000-369X(2017)06-575-11
2017-04-28
2017-07-03
國家自然科學基金面上項目(31570691)
沈威,男,碩士研究生,主要從事茶樹分子生物學研究。*通訊作者:zhuangjing@njau.edu.cn
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
現代企業(2015年9期)2015-02-28 18:56:50
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
