跨界基因沉默菌株的快速構建及對靶標基因干擾能力的檢測
2017-11-28 06:59:58胡琳敏楊霄旭王金向雙林魏晨曦
湖南師范大學學報·自然科學版 2017年5期
胡琳敏+楊霄旭+王金+向雙林+魏晨曦

摘 要 “跨界基因沉默”是一種利用大腸桿菌在哺乳動物細胞中傳遞小干擾RNA并引起基因沉默的新穎技術,具有實用、穩定和低成本等優點.該技術通過構建一種能轉錄出短發夾RNA (shRNA) 的大腸桿菌,而這種大腸桿菌產生的shRNA能靶向哺乳動物細胞中的特定基因,從而引發RNA干擾(RNAi).本文用RecA同源重組的方法將跨界基因沉默質粒的工作元件快速地整合到大腸桿菌的染色體DNA上,構建了跨界基因沉默菌株.通過吖啶橙染色實驗證明了跨界基因沉默菌株能夠入侵哺乳動物細胞,最后Western blot結果表明跨界基因沉默菌株可以對靶標基因產生干擾作用.該技術的完善將進一步促進工程菌的研發和利用.
關鍵詞 siRNA傳遞;跨界基因沉默菌株;RNA干擾;RecA同源重組
中圖分類號 Q812;Q9399 文獻標識碼 A 文章編號 1000-2537(2017)05-0029-07
Fast Construction of Trans-kingdom RNAi Bacteria and Detection of Their Ability to Silence Expression of Target Genes
HU Lin-min1#, YANG Xiao-xu1,2#, WANG Jin1, XIANG Shuang-lin1, WEI Chen-xi1*
(1. The National and Local Joint Engineering Laboratory of Animal Peptide Drug Development,
School of Life Sciences, Hunan Normal University, Changsha 410081, China;
2. The Medicine School, Hunan University of Chinese Medicine, Changsha 410208, China)
Abstract Trans-kingdom RNA interference (TkRNAi) is a useful, stable and low-cost approach for small interference RNA delivery, which employs the genetically engineered E.coli to produce short hairpin RNA and then deliver it into mammalian cells. The shRNA, transcribed by the genetically engineered E.coli, can target and silence the particular gene expression in mammalian cells. We used RecA homologous recombination to quickly integrate working components of TkRNAi into the chromosome DNA of engineered E.coli, thus yielding trans-kingdom interference bacteria. Acridine orange staining tests demonstrated that the trans-kingdom interference bacteria could invade into mammalian cells. Finally, western blot analysis results showed that the trans-kingdom interference bacteria could silence expression of target genes. The improvement of the TkRNAi technology will further promote the development and utilization of this kind of engineering bacteria.
Key words siRNA delivery; trans-kingdom RNAi bacteria; RNAi; RecA homologous recombination
RNA干擾(RNA interference, RNAi)是由小干擾RNA(small interference RNA, siRNA)引起的細胞內序列特異性基因沉默,這給惡性腫瘤等疾病的治療帶來了新的希望[1].然而,siRNA的低血清穩定性、易脫靶性和易引起免疫反應等缺點,也給臨床應用帶來了很大的挑戰[2].此外,鑒于siRNA自身的結構,導致其不能直接穿過細胞膜,因此找到一種穩定、安全和有效的siRNA傳遞方式十分重要[3].
“跨界基因沉默”技術利用工程菌攜帶短發夾RNA(short hairpin RNA, shRNA),將shRNA傳遞至哺乳動物細胞,引發哺乳動物細胞特定基因的沉默.這種新型siRNA傳遞技術的出現在某種程度上彌補了RNA干擾藥物的傳遞困境,具有成為臨床藥物載體的潛力[4-6].“跨界基因沉默”技術由向雙林等提出[7],其基本流程為:首先構建跨界干擾質粒 (Trans-kingdom RNAi plasmid, TRIP) pT7-RNAi-inv-hly.該質粒上的inv基因,其表達產物為Invasin蛋白,能與表皮細胞膜上表達的β1-整合蛋白 (β1-integrin) 結合,促使細胞內陷并吞噬附著于該區域的細菌;hly基因,其表達產物為分泌蛋白LLO,使細菌能夠從細胞內體中逃離[8-9].將TRIP轉化到大腸桿菌BL21(DE3)(以下簡稱為BL21(DE3))內,最后將轉化TRIP的BL21(DE3)侵染哺乳動物細胞,即可在哺乳動物細胞中發揮RNAi作用[7, 10].然而,質粒轉入BL21(DE3)后游離于細菌的胞質中,在無抗性的條件下,質粒容易因環境壓力而丟失,不利于質粒穩定存在并表達shRNA.若能使跨界基因沉默技術更加穩定方便,將有利于拓展跨界基因沉默技術的使用范圍[11].本研究對跨界基因沉默技術進行改進,將TRIP的工作元件整合到BL21(DE3)的基因組中,改進后的整合型菌株克服了質粒易丟失的缺點,且菌株的保存與使用比質粒更方便.endprint
在原核生物中,由RecA介導的同源重組技術已經很成熟[12-14].RecA蛋白在細菌中廣泛存在且高度保守,在細菌的同源重組途徑中起著重要的作用.RecA同源重組系統可介導1 000 bp長同源序列的同源重組,利用大腸桿菌依賴的RecA蛋白重組系統,可以對大腸桿菌基因組進行修飾和基因替換等改造[15-16].
本研究以BL21(DE3) 基因組中的lac Z為模板,克隆出一段長2 027 bp的序列,將其作為lac Z的同源序列插入到TRIP中,構建出環狀整合型跨界基因沉默質粒(Cyclic Integrated Trans-kingdom RNAi plasmid, CI-TRIP).CI-TRIP無須線性化即可快速將TRIP上的工作元件整合入BL21(DE3) 基因組中,實現對BL21(DE3) 基因組的改造.隨后利用CI-TRIP構建了跨界基因沉默菌株,并檢測該菌株侵染哺乳動物細胞SW480的能力以及對SW480細胞中特定基因的干擾能力.
1 材料與方法
1.1 實驗材料
E.coli BL21(DE3),E.coli,GBpir菌,質粒TRIP,質粒pR6k及質粒pCre等為本實驗室保存;人結直腸癌細胞株 (SW480) 為本實驗室保存的細胞株;質粒小量提取試劑盒、膠回收試劑盒購于東盛生物公司;限制性核酸內切酶Pci Ⅰ,限制性核酸內切酶NgoM Ⅳ,限制性核酸內切酶BamH Ⅰ,限制性核酸內切酶Sal Ⅰ和T4 DNA連接酶購自NEB;Anti-LLO antibody,Anti-GAPDH antibody和Anti-β-catenin antibody購于武漢博士德公司;HRP標記的羊抗鼠二抗購于Santa Cruz公司;氯霉素、氨芐青霉素鈉、4%多聚甲醛、DNA分子量標準、氯化鈉、硼酸、瓊脂糖、酵母提取物等購于生工生物工程(上海)股份有限公司;吖啶橙購于Sigma公司;ECL化學發光試劑購于Merck Millipore公司.培養細胞所用的DMEM培養基和胎牛血清購自美國Gibco公司.
1.2 實驗方法
1.2.1 環狀整合型跨界基因沉默質粒的構建 以質粒pR6k為模板,用引物R6k F1和R6k R1擴增出oriR6k-cm(cm代表氯霉素抗性基因)片段,在氯霉素抗性基因兩側設計同方向的LoxP72位點;以BL21(DE3) 基因組為模板,用引物lac F1和N-lac R1擴增出lac Z的重組片段.兩者的PCR產物分別用膠回收試劑盒純化,測定濃度后于-20 ℃保存備用.將oriR6k-cm片段和lac Z重組片段等量混合,以此混合物為模板,以R6k F1,N-lac R1為引物擴增出oriR6k-cm-lac Z基因片段.引物序列如表1所示.
將TRIP中的基因片段hly-inv用Pci Ⅰ和NgoM Ⅳ雙酶切,回收hly-inv基因片段;再用相同的酶剪切oriR6k-cm-lac Z,然后將這兩部分連接.將連接產物用熱轉化法轉入GBpir菌,用含氯霉素抗性(終濃度為15 mg/L)的固體培養基篩選,置于37 ℃培養箱中培養.第二天挑單克隆擴增,用菌落PCR初篩,之后提取陽性菌落中的質粒,雙酶切驗證,最后將驗證為陽性的質粒送上海生工測序.將測序正確的質粒命名為CI-TRIP,即帶有hly-inv基因的環狀整合型跨界基因沉默質粒.
1.2.2 CI-TRIP整合到跨界基因沉默菌株基因組 取CI-TRIP加到BL21(DE3) 中,用熱轉化法處理,置于37 ℃細菌培養箱過夜培養.第二天從固體培養基中挑取單克隆重組子,接種于1.5 mL離心管中,離心管先用無菌針頭在管蓋上戳一個小孔,并加1.0 mL帶氯霉素抗性的LB液體培養基.最后將離心管放在恒溫搖床上,37 ℃,1 000 r/min培養12~16 h.將重組子菌液做模板,用PCR初篩,inv的引物如表1所示,將陽性重組子菌液復壯,第二次PCR篩檢用相同的引物,以細菌基因組DNA為模板.PCR條件為:95 ℃預變性5 min;95 ℃變性30 s,60 ℃退火1 min,72 ℃延伸5 min,循環30次;72 ℃后延伸7 min.將陽性菌落的菌液加入甘油管中,于-80 ℃保種.
1.2.3 細菌基因組中抗性基因的敲除 構建CI-TRIP時,筆者已在氯霉素抗性基因兩側設計了同方向的LoxP72位點,因此可用Cre-LoxP重組酶系統來刪除抗性基因.取1 mL細菌復壯后,加1 μL pCre質粒到管中,混勻后轉移到2 mm電擊杯中.電壓為2.5 kV,5.0 ms電擊一次.電擊轉化后,37 ℃,250 r/min復蘇1 h.將菌液沉淀濃縮至100 μL,涂到氯霉素/氨芐青霉素雙固體培養基上,30 ℃過夜培養.長出單克隆后,挑取接種到含氨芐青霉素液體培養基中,30 ℃,250 r/min培養2 h,37 ℃繼續培養2 h.將菌液分別接種到無抗性的LB固體培養基和氯霉素固體培養基,進行雙固體培養基對照檢測.選取無抗性的菌落,以inv c1f/inv c1r為引物做菌落PCR.以菌落PCR為陽性結果的細菌基因組DNA做模板,用引物inv c1f/inv c1r和cm f/cm r(引物序列如表1所示)進行第二次PCR驗證,inv c1f/inv c1r擴增為陽性而cm f/cm r擴增為陰性的菌落即為抗性敲除成功的跨界基因沉默菌株.將此菌液復壯,當OD600達到0.4~0.6時,按500 μL/管分裝于甘油管內,-80 ℃保種待用.
1.2.4 Western blot檢測跨界沉默菌株中LLO蛋白的表達 從固體培養基上挑單克隆接種到LB培養液中,過夜培養.將菌液離心后收集上清到新管中.用TCA-丙酮沉淀法提取LLO分泌蛋白,根據沉淀量加入SDS裂解液,-20 ℃保存備用.用Western blot檢測LLO的表達情況,取50 μg蛋白質進行SDS-聚丙烯酰胺凝膠電泳,膠濃度為10%,電壓為100 V;轉PVDF膜,恒電流400 mA,時間為80 min.用5%脫脂奶粉封閉,孵育LLO蛋白質的一抗(1∶ 2 000).洗去一抗,孵育HRP標記的羊抗鼠二抗(1∶ 2 000),最后用ECL化學發光試劑顯影,并用Tanon-2500全自動數碼凝膠圖像分析系統(上海天能科技有限公司)記錄實驗結果.endprint
1.2.5 吖啶橙染色檢測細菌侵染SW480細胞的能力 在6孔板中放入玻璃蓋玻片,將細胞以每孔1×105個接種于孔板內,用于細胞爬片.之后置于37 ℃,5%CO2,90%相對濕度的細胞培養箱中培養.從固體培養基上復蘇細菌進行侵染實驗.經IPTG誘導2 h后,當細菌培養到OD600=1.0時終止培養,取出菌株用于侵染SW480細胞.用無血清、無抗生素的細胞培養基洗滌細菌2次,最后用細胞培養基重懸細菌,完全混勻.根據實驗中需要的MOI值(細胞與細菌的比值) (1∶ 500/1∶ 1 000/1∶ 1 500) 稀釋細菌.準備進行細菌侵染試驗前0.5 h,先將細胞的培養基更換成無血清、無抗生素的培養基,0.5 h后,加入含有細菌的培養液,輕搖培養板使細菌均勻分布.1.5~4 h后去除含細菌的培養基,加入含慶大霉素的培養基,用以殺死細胞外的細菌.將細胞用4%多聚甲醛固定液固定處理,PBS洗滌后在搖床上輕搖.用通透液(V丙酮∶ V甲醇=1∶ 1)處理30 s,再用PBS洗滌.用0.01%吖啶橙工作液以1 mL/孔加入孔板中,染色45 s,立即用PBS沖洗.用濾紙吸去多余的水,然后用封閉膠水將玻璃片封閉固定,最后用激光掃描共聚焦顯微鏡(德國Zeiss LSM710)觀察拍照,保存實驗結果.
1.2.6 跨界基因沉默菌株BL21(DE3)-TRIP-CTNNB1的構建 利用1.2.1的質粒CI-TRIP快速構建跨界基因沉默菌株BL21(DE3)-TRIP-CTNNB1.目標基因CTNNB1的發夾結構干擾序列為,sense: CACCGGATGTGGATACCTCCCAAGTCGAAACTTGGGAGGTATCCACATCC; antisense: AAAAGGATGTGGATACCTCCCAAGTTT-CGACTTGGGAGGTATCCACATCC.將這兩段干擾序列合成后,95℃1 min,37℃1 h退火,將CI-TRIP質粒用BamH Ⅰ和Sal Ⅰ雙酶切,最后將干擾序列與CI-TRIP質粒連接.經測序,連接正確的質粒就是BL21(DE3)-TRIP-CTNNB1.取1 μL CI-TRIP-CTNNB1加入BL21(DE3)感受態中,獲得重組菌.重組菌的鑒定方法同1.2.3.用Western blot檢測跨界基因沉默菌株BL21(DE3)-TRIP-CTNNB1的干擾能力,ImageJ軟件分析目的條帶的光密度值.采用t-檢驗判斷實驗組和對照組之間均值的差異顯著性,顯著水平定為P<0.05.
2 實驗結果
2.1 同源臂擴增及環形跨界整合質粒的構建與鑒定
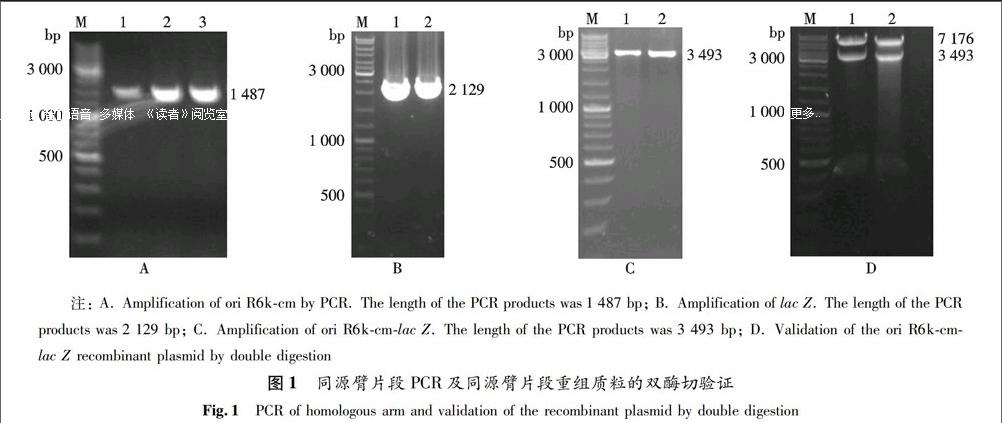
首先,擴增出同源臂片段,如圖1所示.以質粒pR6k為模板,用引物擴增出oriR6k-cm片段,其長度為1 487 bp(圖1A);以BL21(DE3) 基因組為模板,用引物擴增出lac Z的重組片段,其長度為2 129 bp(圖1B).將兩者的PCR產物分別用膠回收試劑盒回收,然后將兩種產物等量混合,以此混合物為模板,PCR擴增出oriR6k-cm-lac Z基因片段,其長度為3 493 bp(圖1C).再將CI-TRIP和oriR6k-cm-lac Z基因片段分別用Pci Ⅰ和NgoM Ⅳ雙酶切,將兩者的連接產物轉入GBpir菌.然后提取陽性菌落中的質粒,并進行雙酶切驗證(圖1D).實驗結果表明,環形跨界基因沉默整合質粒被成功構建,并被命名為CI-TRIP.
2.2 利用CI-TRIP構建跨界基因沉默菌株
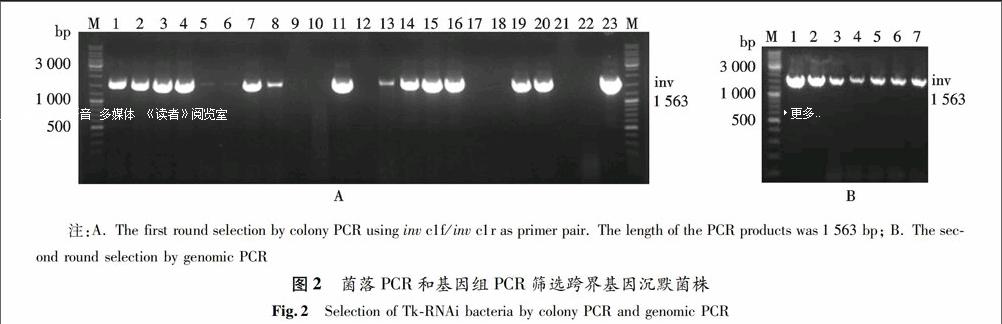
將CI-TRIP轉進BL21(DE3),用引物inv c1f/inv c1r進行菌落PCR初篩.如圖2A中的1,2,3,4,7,8,11,13,14,15,16,19,20,23號菌液擴增出1 563 bp的鑒定條帶,因為inv是跨界基因沉默質粒元件中的一段序列,對于改造后的BL21(DE3) 而言,這是一個外源的基因,能用菌落PCR擴增出inv的目的條帶,說明這些菌經CI-TRIP已將其他元件也整合到其染色體上了.為了確認這些菌是否確實為陽性,筆者將上述部分陽性菌液作為候選菌株繼續培養,并提取基因組DNA,然后用相同引物進行第二輪PCR.如圖2B所示,7個泳道都有目的條帶,且條帶的大小和位置正確.通過兩輪PCR驗證,跨界基因沉默菌株已構建成功.
2.3 Cre重組酶刪除抗性基因及驗證
雖然跨界基因沉默菌株已經構建成功,但是菌株的基因組上還含有質粒上的抗性基因,因此要刪除整合到染色體上的抗性基因,使其成為安全的siRNA傳遞載體.通過Cre-LoxP重組酶系統刪除抗性基因后,跨界基因沉默菌株有可能同時丟失已整合的inv-hly工作元件.將只在無抗性的LB固體培養基上生長的菌落進行菌落PCR,如圖3A中泳道5,10所示,工作元件已丟失;而泳道1,2,3,4,6,7,8,9為刪除抗性基因后,工作元件仍然存在,為陽性菌.取部分陽性菌進一步做PCR驗證,提取細菌的基因組DNA,并用引物inv c1f/inv c1r和cm f/cm r進行PCR檢測,若在這一輪PCR篩選中引物inv c1f/inv c1r擴增產物為陽性而cm f/cm r擴增產物為陰性,則說明跨界基因沉默作用元件已穩定整合到該菌株的基因組染色體上,而抗性基因cm被成功刪除.如圖3B所示,泳道1,2,3,4都有目的條帶,說明工作元件未丟失.如圖3C所示,泳道1,2,3,4都無目的條帶,說明抗性基因刪除成功,泳道p為陽性對照,圖3B和3C中編號相同的泳道其模板來源相同.實驗結果表明,抗性基因cm已經成功被刪除.
2.4 檢測跨界基因沉默菌株中LLO蛋白的表達
跨界基因沉默菌株的基因組DNA整合了TRIP中的hly和inv基因,為了檢測上述整合進細菌基因組的跨界干擾元件能否正常工作,筆者通過Western blot和侵染實驗來進行驗證.endprint
首先,通過Western blot檢測跨界基因沉默菌株是否能夠穩定表達hly基因的編碼產物LLO蛋白.如圖4所示,BL21(DE3) 為空載對照,TRIP為含TRIP質粒的BL21(DE3),B-CI-TRIP為已將跨界基因沉默質粒整合到基因組且刪除抗性基因的BL21(DE3).結果表明跨界基因沉默菌株能正常表達LLO蛋白.
2.5 跨界基因沉默菌株侵染細胞的能力
其次,用侵染實驗來檢驗跨界基因沉默菌株是否能穩定表達inv基因,從而有入侵細胞的能力.吖啶橙能透過細胞膜,并與細胞核中的核酸結合,在藍光的激發下發出綠色熒光.如圖5(彩圖見封三)所示,SW480細胞內最大的綠色熒光團為細胞核,細胞質處有許多綠色小亮斑即為入侵的細菌.用空載細菌檢測侵染細胞的能力,可見SW480細胞質中沒有入侵的細菌,由此可知,空載菌不能侵染哺乳動物細胞(圖5A).在相同處理條件下,用攜帶了跨界基因沉默整合元件的菌株去侵染細胞,SW480細胞質中有入侵的細菌(圖5B).以上結果表明,筆者構建的跨界基因沉默菌株具有入侵哺乳動物細胞的能力.
2.6 跨界基因沉默菌株干擾靶基因的能力
用1.2.6中的方法,構建干擾CTNNB1基因的跨界基因沉默菌株,將其命名為BL21(DE3)-TRIP-CTNNB1.將構建好的跨界基因沉默菌株侵染SW480細胞,然后用Western blot來檢測此跨界基因沉默菌株對細胞中靶基因的干擾能力.結果如圖6所示,跨界基因沉默菌株BL21(DE3)-TRIP-CTNNB1能在48 h和72 h顯著下調SW480細胞中β-catenin蛋白的表達,將β-catenin的表達量分別降至NC組(不用細菌處理細胞)的76.3%(P<0.01)和61.3%(P<0.05).這表明跨界基因沉默菌株可攜帶shRNA,并能抑制哺乳細胞中目的基因的表達.
3 討論
RNAi是一種由雙鏈RNA引起的基因沉默現象,能抑制靶基因的表達,具有特異性和高效等優點.對哺乳動物細胞來說,要使外源的siRNA起基因沉默作用,siRNA的傳遞成為限制RNAi技術發展的瓶頸之一,更限制了特異性干擾作用應用于臨床藥物開發.向雙林等在2006年提出了“跨界基因沉默”技術,利用基因工程改造后的工程細菌攜帶shRNA侵染細胞,并在細胞內釋放shRNA的特點,使RNAi的運用不再因傳遞的問題而被限制.但是TRIP轉入大腸桿菌后容易因環境壓力而丟失,從而喪失跨界基因沉默的功能.如果能使質粒整合到跨界基因沉默菌株的基因組內,那么跨界基因沉默系統會變得更加穩定方便,這也將有利于拓展跨界基因沉默技術的使用范圍.將來可以把益生菌改造成“跨界基因沉默”菌株,當人們飲用這種益生菌到腸道后,它不僅對身體有益,還能發揮RNAi作用,這種一舉兩得的方式將使得臨床治療變得更為便捷.
基于以上考慮,筆者對跨界基因沉默技術進行完善.首先運用傳統分子克隆的方法構建環形跨界基因沉默整合質粒 (CI-TRIP),然后通過RecA介導的同源重組將CI-TRIP上的跨界基因沉默元件成功地整合到大腸桿菌 (BL21-DE3) 的基因組中,并通過Cre-LoxP重組酶系統刪除跨界基因沉默菌株基因組中的抗性基因.隨后利用Western blot和細菌侵染實驗來檢測跨界基因沉默菌株中基因組的跨界干擾元件是否能正常工作.結果表明,抗性刪除的基因沉默菌株不僅能穩定表達LLO蛋白,而且具有細胞侵染的能力.
為了驗證構建出的基因沉默菌株是否能在細胞中發揮其基因沉默作用,筆者對跨界基因沉默菌株進行了功能檢測:以CTNNB1基因做為靶標,用環形跨界基因沉默整合質粒構建了BL21(DE3)-TRIP-CTNNB1跨界基因沉默菌株,此菌株帶有能引起CTNNB1基因沉默的shRNA序列.將干擾菌株侵染細胞后,用Western blot來檢測菌株對靶標基因的干擾作用.結果表明,跨界基因沉默菌株可通過傳遞shRNA抑制細胞中目的基因的表達.
在本研究中,改良后的跨界基因沉默整合菌株解決了載體細菌中質粒容易丟失的缺點,提高了跨界沉默菌株的穩定性,同時表現出無抗性的特點,而且還能有效地沉默細胞中靶基因,這使跨界基因沉默技術的使用變得更方便.在今后的研究中,筆者還將對跨界基因沉默技術進一步改造,使該菌株真正成為既能治療疾病又對人體無害的工程菌.
參考文獻:
[1] LARES M R, ROSSI J J, OUELLET D L. RNAi and small interfering RNAs in human disease therapeutic applications[J]. Trends Biotechnol, 2010,28(11):570-579.
[2] KIM S H. Local and systemic delivery of VEGF siRNA using polyelectrolyte complex micelles for effective treatment of cancer[J]. J Control Rel, 2008,129(2):107-116.
[3] HONG C A, NAM Y S. Functional nanostructures for effective delivery of small interfering RNA therapeutics[J]. Theranostics, 2014,4(12):1211-1232.
[4] AIGNER A. Transkingdom RNA interference (tkRNAi) as a new delivery tool for therapeutic RNA[J]. Expert Opin Biol Ther, 2009,9(12):1533-1542.endprint
[5] NGUYEN T A, FRUEHAUF J H. Transkingdom RNA interference (tkRNAi): a novel method to induce therapeutic gene silencing[J]. Methods Mol Biol, 2009,514:27-34.
[6] GRILLOT C C. Functional gene transfer from intracellular bacteria to mammalian cells[J]. Nat Biotechnol, 1998,16(9):862-866.
[7] XIANG S L, FRUEHAUF J, LI C J. Short hairpin RNA-expressing bacteria elicit RNA interference in mammals[J]. Nat Biotechnol, 2006,24(6):697-702.
[8] MATHEW E. Cytosolic delivery of antisense oligonucleotides by listeriolysin O-containing liposomes[J]. Gene Ther, 2003,10(13):1105-1115.
[9] HEUCK A P. Mechanism of membrane insertion of a multimeric beta-barrel protein: perfringolysin O creates a pore using ordered and coupled conformational changes[J]. Mol Cell, 2000,6(5):1233-1242.
[10] XIANG S L, KEATES A C, FRUEHAUF J, et al.. In vitro and in vivo gene silencing by Transkingdom RNAi (tkRNAi)[J]. Methods Mol Biol, 2009,487:147-160.
[11] LAGE H, KRUHN A. Bacterial delivery of RNAi effectors: transkingdom RNAi[J]. J Vis Exp, 2010(42):e2099.DOI:10.3791/2099.
[12] YADAV T. Roles of bacillus subtilis DprA and SsbA in RecA-mediated genetic recombination[J]. J Biol Chem, 2014,289(40):27640-27652.
[13] CHEN Z, YANG H, PAVLETICH N P. Mechanism of homologous recombination from the RecA-ssDNA/dsDNA structures[J]. Nature, 2008,453(7194):489-494.
[14] LIN Z. Origins and evolution of the recA/RAD51 gene family: Evidence for ancient gene duplication and endosymbiotic gene transfer[J]. Proc Natl Acad Sci USA, 2006,103(27):10328-10333.
[15] 戴 倩,田 云,盧向陽. 同源重組介導的克隆策略研究進展[J]. 農產品加工(學刊), 2011(6):7-9,13.
[16] 楊獻偉,楊瑞馥,崔玉軍. 細菌基因組同源重組:量化與鑒定[J]. 遺傳, 2016(2):137-143.endprint
