寬皮柑橘種質資源表型多樣性分析及綜合評價
2017-11-28 07:31:49孫珍珠李秋月王小柯趙婉彤薛楊馮錦英劉小豐劉夢雨江東
中國農業科學 2017年22期
孫珍珠,李秋月,王小柯,趙婉彤,2,薛楊,2,馮錦英,2,劉小豐,2,劉夢雨,江東,2
?
寬皮柑橘種質資源表型多樣性分析及綜合評價
孫珍珠1,李秋月1,王小柯1,趙婉彤1,2,薛楊1,2,馮錦英1,2,劉小豐1,2,劉夢雨1,江東1,2
(1西南大學柑桔研究所,重慶400712;2中國農業科學院柑桔研究所,重慶400712)
分析寬皮柑橘的表型多樣性和遺傳變異規律,探討柑橘種質資源的綜合評價方法。本研究利用變異系數、Shannon-Weaver多樣性指數及方差分析對239份寬皮柑橘的18種表型性狀進行多樣性分析和性狀差異分析,用SPSS進行聚類分析,通過主成分、相關性以及回歸分析對寬皮柑橘種質資源進行綜合評價和評價指標篩選。對239份寬皮柑橘表型多樣性分析,變異系數結果表明種子數、固酸比、可滴定酸含量及單果重的性狀變異比較豐富,果形指數、囊瓣數及可溶性固形物含量的遺傳特性相對穩定。遺傳多樣性指數結果表明,單果重、橫徑、縱徑、囊瓣數、葉柄長、葉長、葉寬、還原糖含量及轉化糖含量在每一級分布比較均勻,果皮光滑度、種子數及固酸比分級較少,且在每個表現型上的分布不均勻。對5個不同地理來源的寬皮柑橘資源進行方差分析,結果顯示來源于美國的寬皮柑橘品種可溶性固形物和轉化糖的含量平均值高于其他地理來源;來源于日本的寬皮柑橘單果重、橫徑和縱徑大于其他地理來源,種子數顯著少于其他地理來源;來源于長江流域的寬皮柑橘品種的果型、可溶性固形物、轉化糖及還原糖含量大于珠江流域,但固酸比顯著低于珠江流域。對野生品種、地方品種、選育品種3種不同類型的寬皮柑橘性狀作方差分析,選育品種呈現果型大、果皮光滑、種子少、酸度低、糖度高等特點。系統進化樹和主成分分析圖均表明不同地理來源、不同類型的寬皮柑橘在遺傳水平上存在明顯差異,中國小果資源可作為一個獨立類群,青皮蜜橘是一份獨特的柑橘資源。主成分分析發現,前9個主成分的累計貢獻率達81.94%,表明這9個主成分包含了寬皮柑橘表型性狀的大部分信息。表型性狀的綜合評價分析表明,239份寬皮柑橘綜合評價值均值為0.480,來源于日本的愛媛21號的值最高(0.664),來源于日本的扁橘值最低(0.211)。18個表型性狀與綜合值的相關性分析結果表明,除了果皮光滑度外的17個表型性狀數據均與值呈極顯著相關。采用逐步回歸分析方法,篩選出9個表型性狀:縱徑、果形指數、果皮光滑度、囊瓣數、葉柄長、葉寬、可溶性固形物含量、可滴定酸含量以及還原糖含量。寬皮柑橘種質資源具有較高的表型多樣性,不同地理來源、不同類型的寬皮柑橘性狀存在差異,篩選出的9個表型性狀可作為寬皮柑橘的綜合評價指標。
寬皮柑橘;表型多樣性;不同地理來源;綜合評價
0 引言
【研究意義】由于寬皮柑橘與柑橘屬其他種易發生種間雜交,且在長期栽培和選育過程中產生了許多變異類型和實生后代,加上從其他國家引進了大量品種,大大豐富了中國的寬皮柑橘資源[1]。因此,寬皮柑橘的遺傳多樣性較為豐富,對寬皮柑橘遺傳多樣性進行綜合評價,明晰寬皮柑橘資源的表型多樣性及其表型性狀的綜合評價指標顯得非常重要,不僅可以發掘出具有重要經濟價值的特異資源,對寬皮柑橘種質資源的創新研究也具有積極促進作用,同時也便于加強對寬皮柑資源的高效利用和基因資源的挖掘[2-3]。【前人研究進展】王小柯等[4]利用GBS技術對240份寬皮柑橘進行分類,與用植物形態學分類的結果相吻合,表明寬皮柑橘按照傳統的植物學特征進行分類劃分具有一定的合理性。開展柑橘遺傳多樣性的方法有很多,除了形態學鑒定[5]、孢粉學[6]、細胞學[7]、同工酶[8]、數值分類學[9]等常規方法外,還有已廣泛采用的分子標記方法[10-19]等。CAMPOS等[20]利用形態鑒定和AFLP分子標記的方法,對不同地理來源的63份寬皮柑橘種質進行多樣性研究,兩種方法的結果均表現為顯著的遺傳多樣性;劉勇等[9]對柚類33個品種65個形態學性狀進行了品種數值分類研究,利用主成分分析選出影響力較大的18個性狀。李映志[21]采用SSR、RAPD等分子標記對33份寬皮柑橘地方品種和19份野生柑橘品種的遺傳多樣性進行研究,表明寬皮柑橘野生資源的遺傳多樣性都要高于地方品種。【本研究切入點】利用分子標記技術可以更準確地了解植物遺傳多樣性,但最終仍要跟表型性狀結合起來[22-23],形態學方法是植物遺傳多樣性研究最直觀、最基礎的方法,通過形態學性狀遺傳多樣性研究,能從整體了解資源的豐富程度[23-24],為全基因組關聯分析(GWAS)[25-27]提供可靠的表型數據,有利于挖掘重要的農藝性狀基因。目前關于種質資源多樣性和綜合評價研究的報道很多[28-30],但對大樣本寬皮柑橘的表型多樣性研究和綜合評價國內尚未報道。【擬解決的關鍵問題】本研究對239份不同地理來源的寬皮柑橘地方品種、野生資源及國內外選育品種表型性狀進行多樣性分析,以揭示寬皮柑橘資源表型變異規律并篩選相對合理的表型評價指標,為寬皮柑橘遺傳育種、發掘有利基因提供相應的參考和依據。
1 材料與方法
1.1 試驗材料
所有試材均來自于國家果樹種質重慶柑橘資源圃,該圃位于重慶市北碚區歇馬鎮,屬于中亞熱帶濕潤季風氣候區,年平均溫度18℃,年降水量1 100—1 300 mm。試驗選取的239份寬皮柑橘來自17個國家、13個省(市)的239個寬皮柑橘品種為材料(具體材料名單見電子附表1),其中包括起源于中國南方地區的野生寬皮柑橘15份,地方品種77份,選育品種147份。為進行不同地理起源材料的多態性分析,根據239份寬皮柑橘資源的地理起源將其劃分為5個生態區域類型:珠江流域(22份)、長江流域(116份)、美國(14份)、日本(61份)以及其他國家(26份)。
1.2 數據收集
在2015、2016年連續兩年對239份寬皮柑橘的果實和葉片進行數據采集,選取樹勢中等、樹體健康的植株進行采樣。葉片采樣定在春梢停止生長期,隨機選取樹冠外圍春梢中部具有代表性的老熟葉片10片,果實從樹體各個方位隨機采取10個成熟果用于分析,對果實的單果重、橫徑、縱徑、果形指數、果皮光滑度、果皮厚度、囊瓣數、種子數、可溶性固形物、可滴定酸、抗壞血酸含量、固酸比、還原糖及轉化糖含量,對葉片采集葉柄長、葉長、葉寬及葉形指數共18個農藝性狀(反映果實、葉片表型及果實營養成分指標)進行測定。數據采集和整理方法參照《柑橘種質資源描述規范和數據標準》[31]。
1.3 數據分析
1.3.1 多樣性分析 根據各性狀的平均觀測值(`x)與標準差()進行性狀等級的劃分,按照`x±(其中=0、0.5、1、1.5、2)將各個性狀(除果皮光滑度外)的觀測值劃分為1—10級,果皮光滑度分成6級,分別用1、2、3、4、5、6予以附值。
分布頻率:p=n/n,式中,p表示第個性狀第個變異的分布頻率,n表示第個性狀處于第個變異的材料個數,表示材料的總數。
利用變異系數(coefficient of variation,)表示某一性狀的離散程度。公式:=/(為標準差,為平均值)
利用Shannon-Weaver遺傳多樣性指數(index of genetic diversity,)表示任一組中某一性狀的多樣性程度。公式:
用SPSS19.0進行方差分析,并采用平方歐式距離的方法進行聚類分析。
1.3.2 根據品質性狀對寬皮柑橘進行綜合評價 用模糊隸屬函數計算出各個表型性狀的隸屬函數值,將各個性狀定義在[0,1]閉區間。模糊數學的隸屬函數值計算公式為:U=(X-X)/(X-X)。其中,U表示第個品種第個變異的隸屬函數值;X表示第個品種第個變異的測定值;X和X分別表示各品種第個變異的最大和最小值。
綜合評價方法:(1)將標準化表型性狀數據代入每個主成分中,計算各個主成分的得分,根據各個主成分因子的重要程度,確定權重系數[32],最后計算各份材料的的綜合得分(值),進而篩選出優異的種質資源,再結合逐步回歸分析篩選綜合評價指標。
以上數據的統計與分析使用Excel和SPSS19.0軟件進行。
2 結果
2.1 寬皮柑橘表型性狀遺傳多樣性
2.1.1 239份寬皮柑橘表型性狀的變化及分布特征 對239份寬皮柑橘的18個表型性狀進行描述性分析,由表1可知,18個性狀的變異系數值介于9.51%—72.01%,平均值為30.59%,其中變異系數較大的為種子數、固酸比、可滴定酸含量和單果重,表明以上4個性狀變異幅度大,而變異系數較小的為果形指數、囊瓣數、可溶性固形物含量,表明這3個性狀變異幅度小,遺傳特性較為穩定。遺傳多樣性指數值在1.40—2.06變化,均值為1.88,單果重、橫徑、縱徑、囊瓣數、葉柄長、葉長、葉寬、還原糖及轉化糖含量的遺傳多樣性指數較大,表明這些性狀在每一級中的分布比較均勻;果皮光滑度、種子數及固酸比遺傳多樣性指數較小,表明這幾個性狀表現型較少,且在每個表現型上分布不均勻。
2.1.2 不同地理來源的寬皮柑橘的表型性狀差異 根據239份寬皮柑橘的地理起源,將其劃分為5個不同的生態區域類型,其中源于珠江流域占9.21%、長江流域占48.54%、美國占5.86%、日本占25.52%,其他國家占10.88%,采用方差分析對不同區域寬皮柑橘的表型性狀進行比較。
由表2可以看出,除果形指數、果皮厚度、可滴定酸和抗壞血酸含量性狀差異不顯著外,其余14個性狀均存在顯著差異。其中源于日本的寬皮柑橘種質,在單果重、橫徑、縱徑、葉柄長、葉長和葉寬上顯著大于其他地理來源的相應性狀,而種子數顯著低于其他地理來源的種質;來源于美國的寬皮柑橘種質,可溶性固形物和轉化糖的平均含量高于其他地理來源的種質。對比來源于珠江流域和長江流域的寬皮柑橘可以看出,來源于長江流域的寬皮柑橘單果重、橫經、縱徑、可溶性固形物、轉化糖、還原糖含量均大于珠江流域的種質,而固酸比顯著低于珠江流域的種質。

表1 239份寬皮柑橘種質資源18個性狀變化及分布特征

表2 不同地理來源的寬皮柑橘表型性狀的比較
同性狀間的不同字母表示在<0.05水平下差異顯著。下同
Different letters indicate significant difference at<0.05. The same as below
2.1.3 不同類型寬皮柑橘種質的表型性狀比較 239份寬皮柑橘根據資源類型被分為野生資源(15份)、地方品種(77份)、選育品種(147份)3種類型。3種不同類型的寬皮柑橘的方差分析結果表明(表3),除果皮厚度、囊瓣數及葉長差異不顯著外,其余性狀均存在顯著差異。單果重、橫徑、縱徑等幾個與果型大小相關的性狀不同類型之間存在顯著差異,且按野生品種、地方品種、選育品種的順序依次增大;野生資源的果皮光滑度、果形指數比其他兩種類型要小,說明野生資源的果皮比較粗糙,果形一般為扁圓型,而地方品種、選育品種果皮相對比較光滑,果形一般為高扁圓型;野生資源的葉形指數較大,葉片呈長橢圓形,而地方品種、選育品種一般為橢圓形;選育品種的種子數顯著低于野生資源和地方品種。在果實的內在品質性狀方面,野生品種的可滴定酸含量顯著高于地方品種和選育品種,選育品種的可溶性固形物、還原糖、轉化糖等幾個與含糖量相關的性狀均最大。
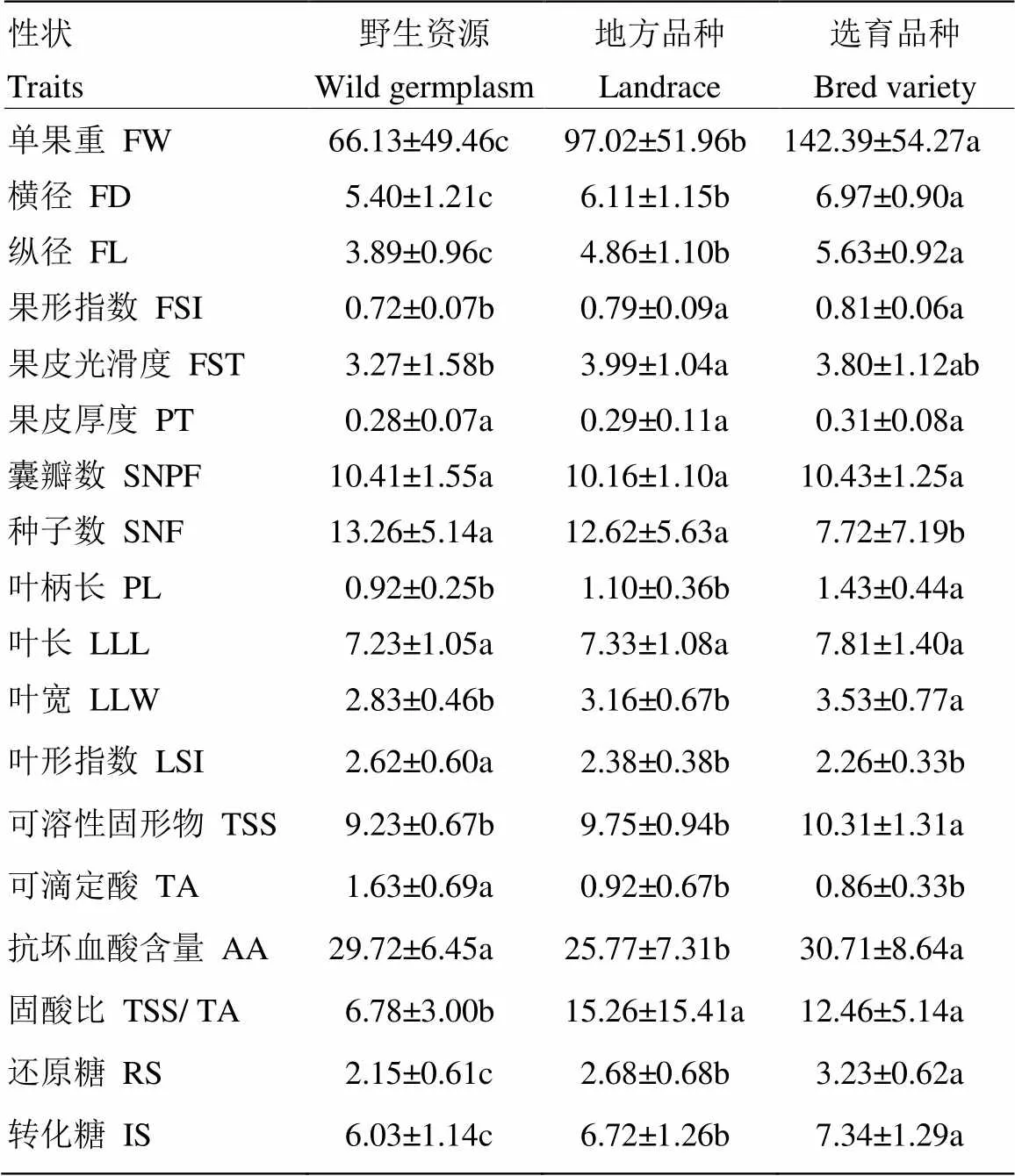
表3 不同類型的寬皮柑橘表型性狀的比較
2.1.4 基于239份寬皮柑橘表型數據的聚類分析 根據表型數據對239份寬皮柑橘進行聚類,結果見圖1,在歐式距離為10時,239份寬皮柑橘可以分為7個類群,類群Ⅰ(僅含有一份資源材料為青皮蜜橘,該種質可滴定酸含量極低);類群Ⅱ(大部分為可溶性固形物較高的品種,包括湘慈43號、費爾柴爾德及一些優異的雜交新品種);類群Ⅲ是最大的類群(主要是橘橙類,還有少量的栽培種和溫州蜜柑,包括默科特橘、艾倫達爾、克里曼丁、砂糖橘及大浦溫州蜜柑等);群體Ⅳ(包含大部分的溫州蜜柑以及少量的橘橙類品種,如久能溫州、杉山溫州、米澤溫州,明尼奧拉等);群體Ⅴ(主要是大果型品種,包括不知火、春見、瀨戶見等);類群Ⅵ(小果酸橘類型,包括印度酸橘、岑溪酸橘及廣西紅皮酸橘等);類群Ⅶ(主要為小果類型的栽培種和野生種,包括本地早橘、南豐蜜橘、莽山野橘及道縣野橘等)。
2.2 寬皮柑橘表型性狀的綜合評價
2.2.1 表型性狀間的相關性分析 對239份寬皮柑橘的18性狀進行相關分析,結果見表4。18個表型性狀間存在不同程度的相關性。其中單果重這一性狀,除了與種子數、葉形指數、可溶性固形物、抗壞血酸及轉化糖含量之間相關性不顯著外,與其余性狀均有顯著或極顯著的相關性;葉柄長與果形指數、葉形指數、可溶性固形物含量、固酸比及轉化糖含量之間的相關性不顯著,與其余性狀均存在一定的顯著相關性;果實的橫徑、縱徑及葉柄長與單果重存在極顯著的正相關,相關系數分別為0.585、0.555和0.517,葉長與葉寬存在極顯著正相關,相關系數為0.656,葉長、葉寬與葉柄長存在極顯著的正相關,相關系數分別為0.515、0.494;轉化糖、還原糖與可溶性固形物含量極顯著正相關,相關系數分別為0.646和0.536。
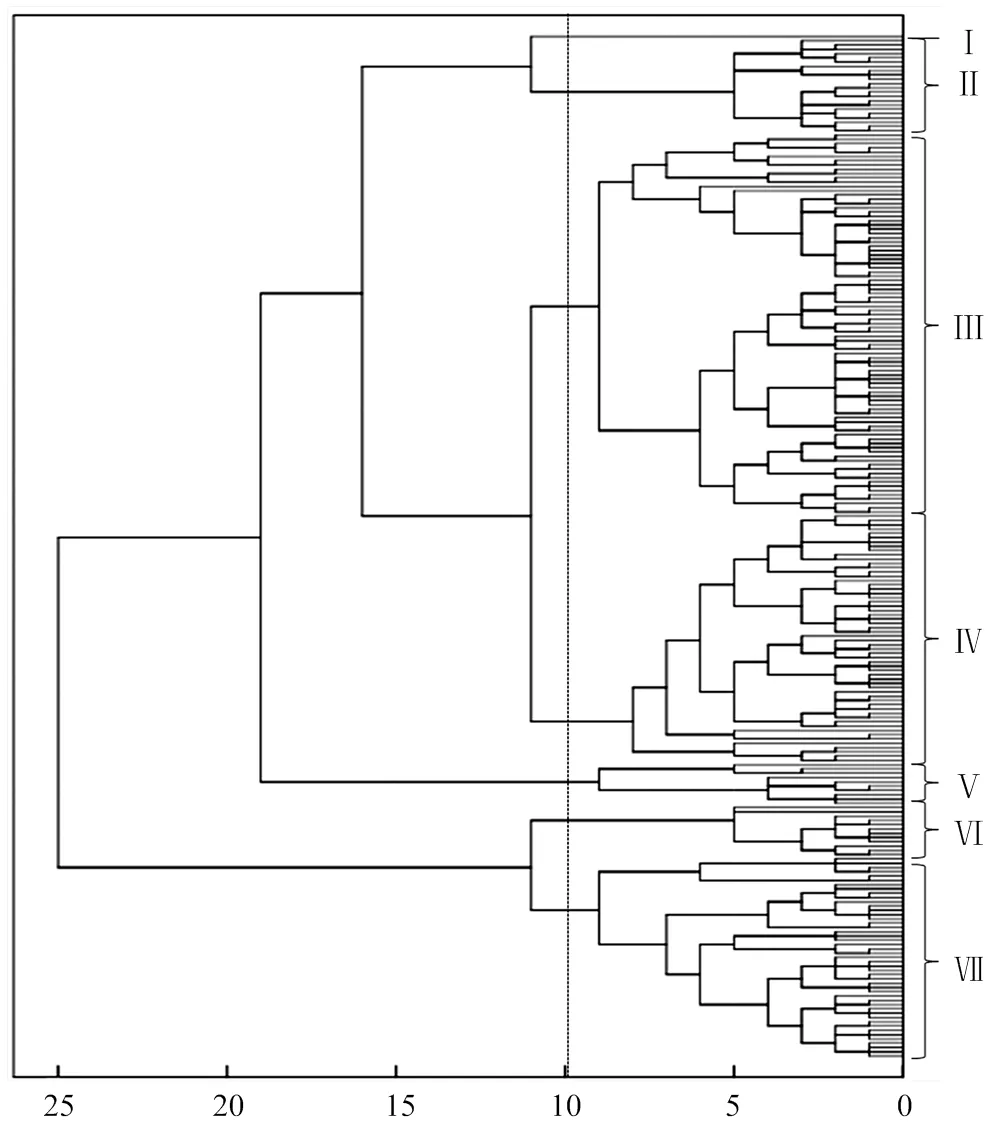
圖1 239份寬皮柑橘基于18個性狀的聚類圖
附表1 85 份谷子材料及其來源
Supplementary Table 1 The names and origins of 85 foxtail millet accessions
2.2.2 表型性狀的主成分分析 對18個表型性狀進行主成分分析(表5),結果顯示前9個主成分的累計貢獻率達81.94%,表明前9個主成分代表了寬皮柑橘表型性狀的大部分信息。將每個主成分向量特征值除以其對應成分的初始特征值的平方根,得到各個成分每個指標的特征向量值,不同成分各個性狀特征向量值、特征值、累計貢獻率如表5所示。
第一主成分貢獻率最大,為18.49%,特征向量值較大的為縱徑、橫徑及單果重,說明第一主成分主要是與果實大小相關的性狀;第二主成分的貢獻率為12.43%,特征向量較大的為可溶性固形物、轉化糖及還原糖,說明第二主成分主要是與含糖量相關的性狀;第三主成分貢獻率為10.12%,其中抗壞血酸、固酸比及可滴定酸的特征向量值大于其他性狀,說明在第三主成分中上述3種性狀是主要性狀;第四主成分貢獻率為8.32%,葉寬的特征向量值最高;第五主成分貢獻率為8.23%,囊瓣數的特征向量值最高,其次是葉形指數,表明在第五主成分中囊瓣數、葉形指數為主要的性狀決定因子;第六主成分的貢獻率為6.81%,特征向量值最大的為葉長;第七主成分貢獻率為6.15%,果形指數、種子數的特征向量值較大;第八主成分的貢獻率為5.78%,果皮光滑度的特征向量值最大;第九主成分的貢獻率5.66%,囊瓣數的特征向量值最大,表明第九主成分中的主要決定因子為囊瓣數。
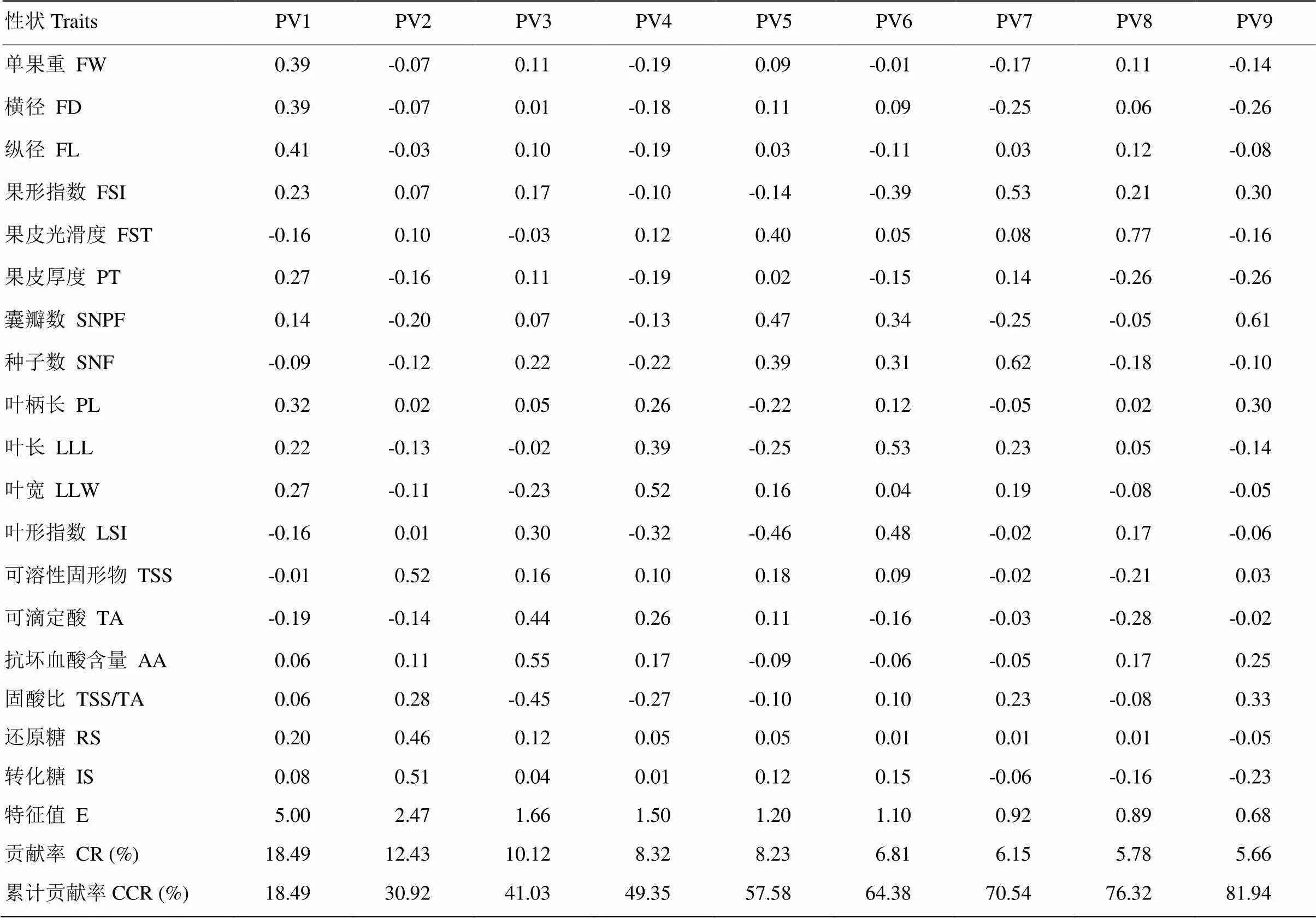
表5 18個表型性狀前9個主成分的特征向量、特征值、貢獻率及累計貢獻率
以第一主成分(果型大小)為橫坐標、第二主成分(含糖量)為縱坐標作主成分散點圖(圖2),將239份寬皮柑橘劃分為4個類群,類群Ⅰ(小果類型的酸橘和野生資源,包括廣西紅皮酸橘、粗皮狗屎柑、江津酸桔及聶都野桔等);類群Ⅱ(其余寬皮柑橘及其雜種品種,包括椪柑、橘橙、溫州蜜柑、清見雜種后代等);類群Ⅲ(只有一份材料為青皮蜜橘,屬于酸度極低的品種);類群Ⅳ(包括1-2、1-14,兩份資源均為愛媛的雜種后代,是含糖量較高的資源)。
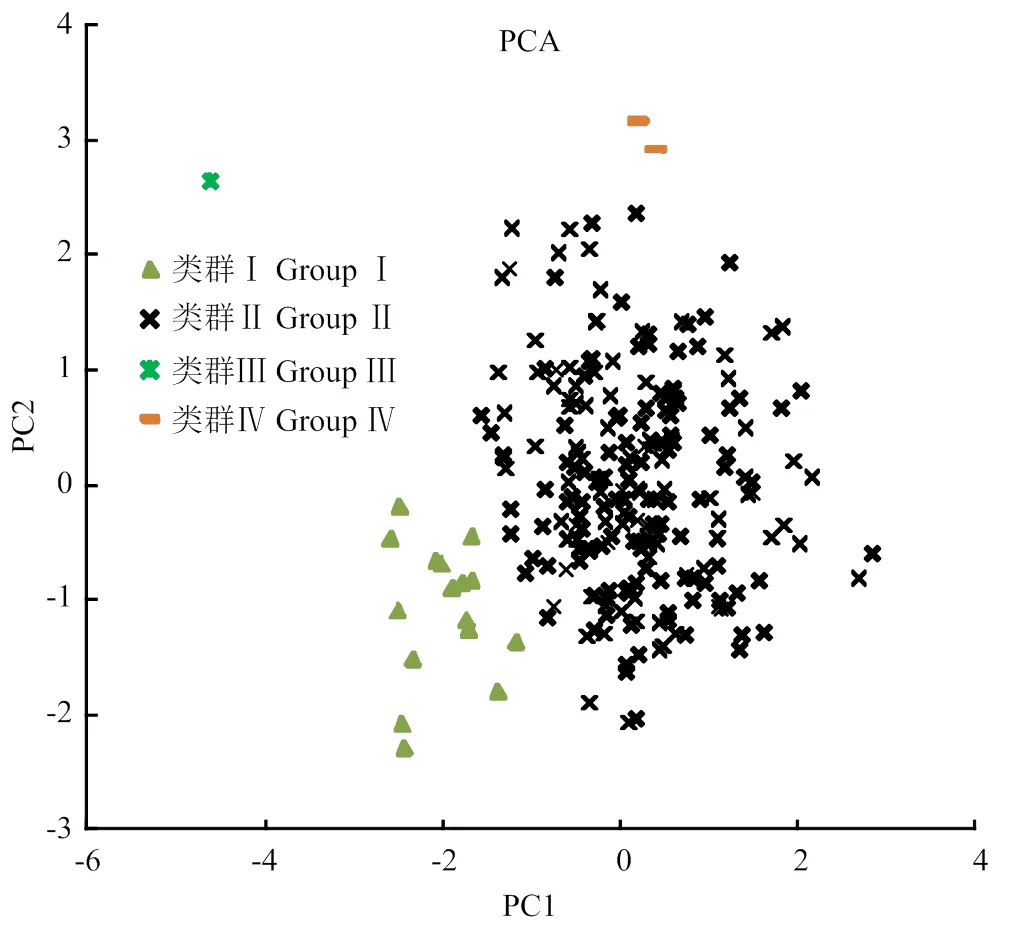
圖2 239份寬皮柑橘的主成分分析圖
2.2.3 寬皮柑橘表型性狀的綜合評價 為對寬皮柑橘進行性狀的綜合評價,根據特征向量矩陣及標準化的表型數據得到主成分得分計算公式,第一主成分得分線性方程如下所示:
F=0.391+0.3920.4130.2340.1650.276+0.147-0.098+0.329+0.2210+0.2711-0.1612-0.0113-0.1914+0.0615+0.0616+0.2017+0.0818
將9個主成分得分用隸屬函數規一化處理,并根據各個主成分在該評價中的重要程度,人為確定各個主成分權重系數(0.35、0.25、0.02、0.05、0.10、0.05、0.03、0.05、0.10),最后得出每份種質材料的綜合得分值(電子附表1),值越高表型綜合性越好,進而對239份寬皮柑橘進行綜合評價。結果表明,239份寬皮柑橘平均值為0.480,其中來自日本的雜柑—愛媛21號,綜合得分值(值)最高(0.664),表明愛媛21號的性狀綜合表現最好;來源于日本的酸橘品種—扁柑,綜合得分(值)最低(0.211),表明扁柑的綜合表現最差。
將18個表型性狀與綜合值做相關性分析(表6),結果表明,除了果皮光滑度外,17個表型性狀數據均與值呈極顯著相關,其中種子數、葉形指數和可滴定酸與值呈極顯著負相關,其余13個性狀呈極顯著正相關。

表6 18個表型性狀與表型綜合值(F)間的相關系數
2.2.4 寬皮柑橘表型性狀綜合指標的篩選 用得到的綜合值與表型性狀構建回歸方程,進一步篩選寬皮柑橘綜合評價指標,構建最優回歸方程為:
=0.109+0.1163+0.0564+0.0555+0.1147+0.0899+0.09011+0.1113-0.04414+0.11617
其中3、4、5、7、9、11、13、14、17等分別代表縱徑、果形指數、果皮光滑度、囊瓣數、葉柄長、葉寬、可溶性固形物、可滴定酸和還原糖含量9個性狀,其直接通徑系數分別為0.280、0.144、0.159、0.255、0.233、0.170、0.263、-0.072、0.218,方程相關系數和決定系數2分別為0.985**和0.971,表明9個表型性狀可決定總變異的97.1%。值為852.335,回歸方程達到極顯著水平,表明以上9個表型性狀可作為寬皮柑橘綜合評價指標。
3 討論
3.1 寬皮柑橘的表型多樣性分析
本研究采用的239份試驗材料涵蓋了寬皮柑橘的大部分類型,包含的種質資源信息十分豐富,目前國內還沒有對大樣本寬皮柑橘進行表型多樣性分析的相關報道。關于寬皮柑橘多樣性分析的報道,高恒錦等[33]利用SSR、InDel及來源于lncRNA的分子標記對中國不同地理區域的45份野生、半野生及地方寬皮柑橘資源進行遺傳多樣性分析,表明寬皮柑橘中的一些育成品種及雜柑資源具有較豐富的遺傳多樣性;KOEHLERSANTOS[16]利用表型鑒定和分子標記結合的方法對來源于巴西南部的37份寬皮柑橘進行多樣性研究,表型聚類和分子標記聚類結果存在差異;DORJI等[34]對39份寬皮柑橘進行表型多樣性分析,結果表明不同品種間的葉片和果實存在顯著差異,這和本研究的結果是一致的,本研究中主成分分析結果表明果實重量、果實大小和葉寬、葉長性狀對不同寬皮柑橘資源表型差異具有較大貢獻。
不同地理來源的寬皮柑橘品種有其獨有的特性,來源于美國的品種,可溶性固形物和轉化糖高于其他地理來源品種,這與美國地區常用甜橙作親本進行雜交育種有關,并且這類品種常具有晚熟的特性[4]。來源于日本的柑橘種子數顯著低于其他地理來源的品種,因為日本育種工作者偏好用溫州蜜柑、清見等少核品種作為親本,雜種后代也一般表現出少核的性狀。對不同類型的寬皮柑橘進行比較可以看出,與野生品種、地方品種相比,選育品種呈現果型大、果皮光滑、種子少、酸度低、糖度高等特點,這與人們育種的目標和方向是一致的。
系統進化樹和主成分分析圖可以看出青皮蜜橘和中國的小果寬皮柑橘是較為獨特的兩個類群,其中青皮蜜橘是一份獨特的寬皮柑橘品種,其可滴定酸含量較低,而中國小果寬皮柑橘包含了較多的野生寬皮柑橘和酸橘類型資源,該結果與王小柯等[4]采用GBS進行的分析結果是吻合的。同時也可以看出不同地理來源、不同類型的寬皮柑橘在遺傳水平上存在明顯差異,說明地理隔離是增加寬皮柑橘多樣性的一個重要因素,另外育種工作者在選擇雜交親本材料時,既要考慮材料的地理來源也要考慮類群的歸屬[35]。主成分分析圖篩選出了兩份含糖量較高的資源(愛媛的雜種后代1-2、1-14),一份酸度極低的資源(青皮蜜橘),這3份特異資源可以作為雜交育種的親本材料。
3.2 基于表型性狀對寬皮柑橘的綜合評價
本研究基于寬皮柑橘的表型性狀,采用隸屬函數、相關性和主成分分析結合的方法,對寬皮柑橘進行綜合評價,此評價方法已在植物表型綜合評價[28-30]、耐酸堿砧木的綜合評價[36]等方面得到應用。根據各個主成分在該評價體系的重要程度,人為給出各個主成分相應的權重系數,對比按貢獻率確定權重系數的方法而言,此方法確定權重得到的結果更為可靠。通過比較綜合值值的大小篩選出的優異種質資源,可以做為雜交育種親本材料或優異種質在育種和生產中利用。
由于寬皮柑橘表型性狀較多,且彼此間存在一定相關性,對評價和利用寬皮柑橘資源產生一定的影響。本研究通過主成分分析、確定權重系數和逐步回歸分析的方法,篩選出9個性狀。上述性狀可作為寬皮柑橘的綜合評價指標,有利于種質資源評價工作的開展以及種質資源的高效利用,因此,在今后的柑橘評價工作中,這些表型性狀應得到特別關注。
4 結論
利用表型鑒定對239份寬皮柑橘的18個表型性狀統計分析,結果表明寬皮柑橘有較高的表型多樣性,不同地理來源的寬皮柑橘種質有其獨有的特性。日本地區的種質表現為果型大、種子少的特點;沿長江流域傳播的寬皮柑橘果型、可溶性固形物、轉化糖、還原糖含量均大于珠江流域,但固酸比顯著低于珠江流域。和野生品種、地方品種相比,選育品種的綜合表現和人們的育種目標是一致的。群體進化樹和主成分分析圖表明青皮蜜橘是一份獨特的柑橘資源,中國小果資源可作為一個獨立的類群。篩選出縱徑、果形指數、果皮光滑度、囊瓣數、葉柄長、葉寬、可溶性固形物含量、可滴定酸含量及還原糖含量等9個性狀可作為鑒定寬皮柑橘的重要指標。
[1] 鄧秀新. 中國柑橘品種. 北京: 中國農業出版社, 2008.
DENG X X.. Beijing: China Agriculture Press, 2008. (in Chinese)
[2] 王洪新, 胡志昂. 植物的繁育系統、遺傳結構和遺傳多樣性保護. 生物多樣性, 1996, 4(2): 92-96.
WANG H X, HU Z A. Plant breeding system, genetic structure and conservation of genetic diversity., 1996, 4(2): 92-96. (in Chinese)
[3] 王永康, 吳國良, 趙愛玲. 棗種質資源的表型遺傳多樣性. 林業科學, 2014, 50(10): 33-41.
WANG Y K, WU G L, ZHAO A L. Phenotypic genetic diversity of jujube germplasm resources., 2014, 50(10): 33-41. (in Chinese)
[4] 王小柯, 江東, 孫珍珠. 利用 GBS 技術研究 240 份寬皮柑橘的系統演化. 中國農業科學, 2017, 50(9): 1666-1673.
WANG X K, JIANG D, SUN Z Z. Study on phylogeny of 240 mandarin accessions with genotyping-by-sequencing technology., 2017, 50(9): 1666-1673. (in Chinese)
[5] 曾勉. 對柑橘分類的認識體會和整理的意見. 中國果樹, 1962(2): 31-37.
ZENG M. The understanding and opinion of the citrus classification., 1962(2): 31-37. (in Chinese)
[6] 張映南, 李潤唐, 蔣開軍. 幾種柑橘屬植物花粉外壁超微結構比較研究. 果樹學報, 2003, 20(3): 165-168.
ZHANG Y N, LI R T, JIANG K J. A comparative study on pollen exine ultrastructure of several citrIls., 2003, 20(3): 165-168. (in Chinese)
[7] 粱國魯. 柑桔類的細胞分類學研究. 武漢植物學研究, 1990, 8(1): l-7.
LIANG G L. Studies on the cytotaxonomy of citrus., 1990, 8(1): l-7. (in Chinese)
[8] 方德秋, 章文才, 蕭順元. 溫州蜜柑起源新探—同工酶證據. 果樹學報, 1995, 12(1): 16-20.
FANG D Q, ZHANG W C, XIAO S Y. Satsuma mandarin new origin of evidence – isoenzyme., 1995, 12(1): 16-20. (in Chinese)
[9] 劉勇, 孫中海, 劉德春, 吳波, 江東. 部分柚類品種數值分類研究. 果樹學報, 2006, 23(1): 35-40.
LIU Y, SUN Z H, LIU D C, WU B, JIANG D. Study on numerical taxonomy of pomelo cultivars., 2006, 23(1): 35-40. (in Chinese)
[10] LI Y Z, CHENG Y J, YI H L, DENG X X. Genetic diversity in mandarin landraces and wild mandarins from China based on nuclear and chloroplast simple sequence repeat markers., 2006, 81(3): 371-378.
[11] KACAR Y, UZUN A, POLAT I, YESILOGLU T, YILMAZ B, GULSEN O, TUZCU O, KAMILOGLU M, KURT S, SEDAY U. Molecular characterization and genetic diversity analysis of mandarin genotypes by SSR and SRAP markers., 2013, 11(11): 516-521.
[12] BARKLEY N A, ROOSE M L, KRUEGER R R, FEDERICI C T. Assessing genetic diversity and population structure in a citrus germplasm collection utilizing simple sequence repeat markers (SSRs)., 2006, 112(8): 1519-1531.
[13] GARCIA-LOR A, LURO F, OLLITRAULT P. Genetic diversity and population structure analysis of mandarin germplasm by nuclear, chloroplastic and mitochondrial markers., 2015, 11(6): 1-15.
[14] FILHO H D C, MACHADO M A, TARGON M L P N, MOREIRA M C P Q D G. Analysis of the genetic diversity among mandarins (spp.) using RAPD markers., 1998, 102(1): 133-139.
[15] GULSEN O, ROOSE M L. Lemons: diversity and relationships with selected citrus genotypes as measured with nuclear genome markers., 2001, 126(3): 309-317.
[16] KOEHLERSANTOS, PATR CIADORNELLES, CUNHAFREITAS A L, DE L B O. Characterization of mandarin citrus germplasm from Southern Brazil by morphological and molecular analyses., 2003, 38(7): 797-806.
[17] ABKENAR A A, ISSHIKI S. Molecular characterization and genetic diversity among Japanese acid citrus (spp.) based on RAPD markers., 2003, 78(1): 108-112.
[18] PANG X M, HU C G, DENG X X. Phylogenetic relationships within Citrus and its related genera as inferred from AFLP markers., 2007, 54(2): 429-436.
[19] SIRAGUSA M, DE PASQUALE F, ABBATE L, TUSA N. Identification of sour orange accessions and evaluation of their genetic variability by molecular marker analyses., 2006, 41(1): 84-89.
[20] CAMPOS E T, WARBURTON M L, VARELA A S, MONTER A V. Characterization of madarin (spp.) using morphological and aflp markers., 2005, 206(5): 923-926.
[21] 李映志. 寬皮柑橘(Blanco)的遺傳多樣性及系統發育研究[D]. 武漢: 華中農業大學, 2006.
LI Y Z. Research on the genetic diversity and phylogenetic relationship of loose skin mandarins (Blanco) [D]. Wuhan: Huazhong Agricultural University, 2006. (in Chinese)
[22] 陳巍, 王力榮, 朱更瑞. 基于SSR標記和生物學性狀進行桃遺傳多樣性的比較分析. 植物遺傳資源學報, 2009, 10(1): 86-90.
CHEN W, WANG L R, ZHU G R. Comparison of genetic diversity among peach cultivars based on biological traits and SSR markers., 2009, 10(1): 86-90. (in Chinese)
[23] 潘存祥, 許勇, 紀海波. 西瓜種質資源表型多樣性及聚類分析. 植物遺傳資源學報, 2015, 16(1): 59-63.
PAN C X, XU Y, JI H B. Phenotypic diversity and clustering analysis of watermelon germplasm., 2015, 16(1): 59-63. (in Chinese)
[24] 田稼, 鄭殿升. 中國作物遺傳資源. 北京: 中國農業出版社, 1994.
TIAN J, ZHENG D S.. Beijing: China Agriculture Press, 1994. (in Chinese)
[25] SONAH H, O'DONOUGHUE L, COBER E, RAJCAN I, BELZILE F. Identification of loci governing eight agronomic traits using a GBS-GWAS approach and validation by QTL mapping in soya bean., 2014, 13(2): 1-11.
[26] TIEMAN D, ZHU G, JR R M, LIN T, NGUYEN C, BIES D, RAMBLA J L, BELTRAN K S, TAYLOR M, ZHANG B. A chemical genetic roadmap to improved tomato flavor., 2017, 355(6323): 391-394.
[27] MORRIS G P, RAMU P, DESHPANDE S P, HASH C T, SHAH T, UPADHYAYA H D, RIERA-LIZARAZU O, BROWN P J, ACHARYA C B, MITCHELL S E. Population genomic and genome-wide association studies of agroclimatic traits in sorghum., 2013, 110(2): 453-458.
[28] 胡標林, 萬勇, 李霞, 雷建國, 羅向東, 嚴文貴, 謝建坤. 水稻核心種質表型性狀遺傳多樣性分析及綜合評價. 作物學報, 2012, 38(5): 829-839.
HU B L, WAN Y, LI X, LEI J G, LUO X D, YAN W G, XIE J K. Analysis on genetic diversity of phenotypic traits in rice () core collection and its comprehensive assessment., 2012, 38(5): 829-839. (in Chinese)
[29] 王海崗, 賈冠清, 智慧. 谷子核心種質表型遺傳多樣性分析及綜合評價. 作物學報, 2016, 42(1): 19-30.
WANG H G, JIA G Q, ZHI H. Phenotypic diversity evaluations of foxtail millet core collections., 2016, 42(1): 19-30. (in Chinese)
[30] 代攀虹, 孫君靈, 何守樸. 陸地棉核心種質表型性狀遺傳多樣性分析及綜合評價. 中國農業科學, 2016, 49(19): 3694-3708.
DAI P H, SUN J L, HE S P. Comprehensive evaluation and genetic diversity analysis of phenotypic traits of core collection in upland cotton., 2016, 49(19): 3694-3708. (in Chinese)
[31] 江東. 柑橘種質資源描述規范和數據標準. 北京: 中國農業出版社, 2006.
JIANG D.(spp). Beijing: China Agriculture Press, 2006. (in Chinese)
[32] 張海英, 韓濤, 王有年, 李麗萍. 桃果實品質評價因子的選擇. 農業工程學報, 2006, 22(8): 235-239.
ZHANG H Y, HAN T , WANG Y N , LI L P. Selection of factors for evaluating peach () fruit quality., 2006, 22(8): 235-239. (in Chinese)
[33] 高恒錦, 儲春榮, 王小柯, 陳紹彬, 江東. 45份寬皮柑桔野生和地方資源遺傳多樣性分析. 中國南方果樹, 2016, 45(2): 1-9.
GAO H J, CHU C R, WANG X K, CHEN S B, JIANG D. The genetic diversity of landrace mandarin germplasms., 2016, 45(2): 1-9. (in Chinese)
[34] DORJI K, YAPWATTANAPHUN C. Assessment of morphological diversity for local mandarin (Blanco.) accessions in Bhutan., 2011, 7(2): 485-495.
[35] 萬述偉, 宋鳳景, 郝俊杰. 271份豌豆種質資源農藝性狀遺傳多樣性分析. 植物遺傳資源學報, 2017,18(1): 10-18.
WAN S W, SONG F J, HAO J J. Genetic diversity of agronomic traits in 271 pea germplasm resources., 2017, 18(1): 10-18. (in Chinese)
[36] 王海崗, 陳凌, 王君杰, 曹曉寧, 董俊麗, 王綸, 楊天育, 喬治軍. 20份山西糜子種質資源抗旱性綜合評價. 中國農學通報, 2014, 16(36): 1916-1920.
WANG H G, CHEN L, WANG J J, CAO X N, DONG J L, WANG L, YANG T Y, QIAO Z J. Comprehensive assessment of drought resistance of 20 proso millet germplasm resources in whole growth period., 2014, 16(36): 1916-1920. (in Chinese)
(責任編輯 趙伶俐)
Comprehensive Evaluation and Phenotypic Diversity Analysis of Germplasm Resources in Mandarin
SUN ZhenZhu1, LI QiuYue1, WANG XiaoKe1, Zhao WanTong1,2, XUE Yang1,2, FENG JinYing1,2, LIU XiaoFeng1,2, LIU MengYu1, JIANG Dong1,2
(1Citrus Research Institute, Southwest University, Chongqing 400712;2Citrus Research Institute of Chinese Academy of Agricultural Sciences, Chongqing 400712)
The objective of this study is to evaluate some major phenotypic characteristics diversity of mandarin germplasms and explore the comprehensive method for screening important mandarin germplasm resources.The diversity evaluation of 18 phenotypic traits in 239 mandarin accessions were carried out. Variation coefficients, Shannon-Weaver Information index cluster analysis, principal component analysis, correlation and regression analysis were comprehensively used to evaluate the relationship of characteristics and select important characteristics.Based on the variation coefficient of 239 mandarins, large variations were found in the traits of seed numbers per fruit (SNF), the ratio of soluble solids to titratable acidity (TSS/TA), the contents of titratable acid (TA), fruit weight (FW), while genetic characteristics of fruit shape index(FSI), segment numbers per fruit (SNPF), the contents of total soluble solids (TSS) kept relatively stable. Shannon-Weaver Information index showed that the diversity level of FW, fruit diameter (FD), fruit length (FL), SNPF, petiole length (PL), leaf lamina length (LLL), leaf lamina width (LLW), the contents of reducing sugar (RS) and invert sugar (IS) were high, while fruit surface texture(FST), SNF, TSS/TA had low diversity level. The variance analysis was conducted to evaluate the trait difference of 239 mandarin accessions, which divided into five different geographical origins. The result showed that mandarin accessions from America have higher average contents of TSS and IS; the mandarin accessions from Japan have larger FW, FD and FL and lower SNF. Mandarin accessions from the Yangtze River Basin were larger fruit size, higher contents of the TSS, RS and IS than the Pearl River Basin, but lower TSS/TA. Variance analysis for wild germplasms, landraces and bred varieties showed that bred accessions have larger fruits, smoother peels, fewer seeds, lower contents of TA and higher contents of sugar content. Both the cluster analysis and PCA suggested genetic variations were presented in different geographical origins. Small fruit type accession from China should be thought as an unique subgroup, Green Peel Mi Ju is an unique citrus accession. Principal component analysis showed that the variation cumulative contribution rate of the first nine principal components was accounted for 81.94%. The averagevalue from the comprehensive evaluation of phenotypic traits of 239 mandarins was 0.480. The highestvalue (0.664) was given to Ehime Kashi No.21 from Japan. The lowestvalue (0.211) was Shikuwasha. The correlation analysis between the phenotypic traits andvalue showed that the phenotypic traits except FST were significantly correlated with thevalue. Nine phenotypic traits, including fruit length (FL), FSI, FST, SNPF, PL, LLW, the contents of TSS, TA and RS, were selected out as evaluation indexes by stepwise regression analysis.Mandarin accessions presented large phenotypic diversity. The traits difference of the accessions from different geographical origins or germplasm types are great. Nine phenotypic traits could be used as important evaluation charaterisitics for screening unique mandarin.
mandarin; phenotypic diversity; different geographical origins; comprehensive evaluation
2017-05-27;
國家“十二五”科技支撐計劃(2013BAD01B04)、重慶市科委重點項目(cstc2016shms-ztzx80004)
接受日期:2017-08-24
聯系方式:孫珍珠,Tel:18375735360;E-mail:sunzhenzhu417@163.com。通信作者江東,Tel:13983194771;E-mail:jiangdong@cric.cn
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
石油瀝青(2021年4期)2021-10-14 08:50:44
世界科學技術-中醫藥現代化(2021年10期)2021-03-02 05:52:06
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
資源再生(2017年3期)2017-06-01 12:20:59
中國教育技術裝備(2015年19期)2015-03-01 02:43:07
中國工程咨詢(2015年2期)2015-02-14 02:59:26
俄羅斯問題研究(2012年1期)2012-03-25 09:54:51
