水稻OsZFP互作蛋白的篩選與鑒定
2017-11-27 07:21:38鄭秀文石嘉偉楊靜靜陳敏敏劉宏波
浙江農林大學學報 2017年6期
鄭秀文, 崔 鵬, 石嘉偉, 楊靜靜, 陳敏敏, 鄭 瑤, 許 玲, 劉宏波
(1.浙江農林大學 農業與食品科學學院,浙江 杭州 311300; 2.浙江理工大學 生命科學學院,浙江杭州 310018)
水稻OsZFP互作蛋白的篩選與鑒定
鄭秀文1, 崔 鵬1, 石嘉偉1, 楊靜靜1, 陳敏敏2, 鄭 瑤2, 許 玲2, 劉宏波1
(1.浙江農林大學 農業與食品科學學院,浙江 杭州 311300; 2.浙江理工大學 生命科學學院,浙江杭州 310018)
CCHC型鋅指結構蛋白OsZFP參與調控水稻Oryza sativa側根的生長發育,但其相關互作蛋白及調控機制未知。以水稻 ‘日本晴’ ‘Nipponbare’為試驗材料,克隆了OsZFP基因,利用EcoRI和SalI酶切位點構建酵母雙雜交釣餌表達載體pGBKT7+OsZFP,驗證該表達載體對酵母菌株Y2H無毒性及報告基因自激活現象;采用酵母雙雜交技術,從已構建的水稻cDNA文庫中篩選到1個陽性互作蛋白,經美國國家生物技術信息中心數據庫(NCBI)同源性比對,鑒定為含有T-complex polypeptide 1(TCP-1)結構域的分子伴侶蛋白第7個亞基(Os06g0687700),命名為chaperonin containing TCP-1 eta subunit(CCT-eta),進而通過酵母一對一回復驗證該互作蛋白。基于CCT-eta亞基與OsZFP蛋白互作,推測分子伴侶蛋白亞基CCT-eta可能參與調控水稻側根的生長發育。圖4參19
植物學;水稻;調控蛋白;載體構建;酵母雙雜交;蛋白互作
水稻Oryza sativa是世界上重要的禾本科Gramineae糧食作物之一。據估計,到2030年,禾本科作物的產量至少需要提高40%才能滿足人口增長所帶來的糧食需求[1]。生長發育良好的水稻根系不僅使植株適應不利的生長環境,而且還能提高植株對水分和營養的利用率[2],從而保證結實率和產量。水稻的種子根只在植株發育初期起作用,隨后的整個生育期主要通過發育更為完善和發達的側根和冠根維持生長發育。因此,發掘和鑒定調控水稻側根和根冠生長發育的蛋白對水稻根系遺傳發育及分子調控機制研究顯得尤為重要。目前,在水稻根系發育研究中已獲得了一些突變體表型及其調控側根和冠根發育的關鍵基因,這些基因的功能缺失突變能特異地影響根系的生長發育和形態建成[3-5]。水稻側根通過中柱鞘和內皮層細胞不斷分裂而形成[6]。已有研究表明:真核生物中含鋅指結構域蛋白能特異結合DNA,RNA和蛋白質,從而參與調控真核生物細胞的發育和凋亡過程及相關信號傳導途徑[7-8]。擬南芥Arabidopsis thaliana C1-1i亞族中GIS,GIS2,ZFP8和ZFP5蛋白受赤霉素和細胞分裂素誘導,參與調控表皮毛的生長發育及相關信號傳導途徑[9-12];而C1-2i亞族中的Zat6被證實參與調控種子根的生長發育和磷酸鹽的吸收[13]。最近,水稻中已鑒定出189個含C2H2型鋅指結構蛋白,其中至少26個蛋白響應不同的生物脅迫和非生物脅迫[14]。這些研究結果預示著含鋅指結構域的蛋白在水稻生長發育過程中起著重要的調節作用。在前期研究中,我們已鑒定到1個水稻調控側根生長發育的C2HC型鋅指結構蛋白。本研究擬利用酵母雙雜交技術篩選鑒定與該蛋白互作的調控蛋白,以期為水稻根系生長發育分子機制的解析奠定工作基礎。
1 材料與方法
1.1 釣餌表達載體構建
根據酵母表達載體pGBKT7多克隆位點及水稻鋅指結構蛋白基因(OsZFP)表達序列設計引物,正向: 5′-CGGAATTCATGGCAAGGGAACCAAGTCC-3′(下劃線為 EcoRI酶切位點), 反向: 5′-ACGCGTCGACCTAAAATGCCTGGCTCCTAA-3′(下劃線為SalI酶切位點)。以本實驗室保存的水稻品種 ‘日本晴’‘Nipponbare’幼苗為試驗材料,對幼苗cDNA進行聚合酶鏈式反應(PCR)擴增,反應體系和擴增程序參照文獻[15]稍作修改。目的基因經雙酶切回收后連接至酵母表達載體pGBKT7并轉化大腸埃希菌Escherichia coli DH5α,重組酵母表達載體pGBKT7+OsZFP雙酶切驗證后進行單克隆測序。
1.2 釣餌載體毒性和自激活分析
按YeastmakerTMYeast Transformation System 2小量法轉化酵母表達載體pGBKT7和酵母釣餌表達載體pGBKT7+OsZFP至50 μL酵母菌株Y2H感受態細胞,分別稀釋轉化菌液至1/10和1/100,取100 μL酵母表達載體pGBKT7轉化液涂布于SD/-Trp;同時,取100 μL酵母釣餌表達載體pGBKT7+OsZFP轉化液涂布于 SD/-Trp(SDO), SD/-Trp/X-α-Gal(SDO/X)和 SD/-Trp/X-α-Gal/AbA(SDO/X/A)固體培養基, 30℃培養3~5 d,對比pGBKT7和pGBKT7+OsCYP2兩者轉化后酵母單克隆大小,觀察統計結果,檢測釣餌載體是否對酵母菌株有毒性及是否對報告基因存在自激活現象。
1.3 cDNA文庫篩選
按YeastmakerTMYeast Transformation System 2共轉化水稻幼苗cDNA文庫和酵母釣餌表達載體pGBKT7+OsZFP至600 μL酵母菌株Y2H感受態細胞,所有轉化菌液全部涂布于SD/-Leu/-Trp/X-α-Gal/AbA(DDO/X/A)固體培養基,30℃培養3~5 d,挑選藍色單克隆接種至SD/-Leu/-Trp/-His/Ade/X-α-Gal/AbA(QDO/X/A)固體培養基,30℃培養3~5 d,將藍色克隆菌株劃線至SD/-Leu/-Trp//X-α-Gal(DDO/X)固體培養基,重復2~3次,分離陽性互作蛋白。
1.4 互作蛋白的分離和回復驗證
挑取上述DDO/X固體培養基中分離出來的藍色單克隆,用酵母質粒提取試劑盒(easy yeast plasmid isolation kit)提取釣餌與互作蛋白表達質粒,轉化大腸桿菌DH5α感受態細胞,涂布于含有氨芐青霉素的LB固體培養基,37℃過夜培養,挑選單克隆提取質粒,送生物公司測序。將分離的互作蛋白表達質粒,與pGBKT7或pGBKT7+OsZFP分別共轉化酵母菌株Y2H,取100 μL轉化菌液涂布于DDO/X和QDO/X/A固體培養基中,30℃培養3 d,回復驗證陽性互作蛋白。
2 結果與分析
2.1 含OsZFP基因的釣餌載體獲得
PCR擴增OsZFP基因獲得大小為828 bp的目的片段(圖1A),與預期條帶相符;利用EcoRI和SalI酶切位點酶切酵母表達載體pGBKT7及目的基因OsZFP(圖1B),T4連接酶連接后轉化大腸埃希菌DH5α,提取重組表達載體后進行雙酶切驗證(圖1C),陽性克隆送生物公司測序驗證。
2.2 釣餌表達載體毒性及自激活分析
酵母表達載體pGBKT7和釣餌載體pGBKT7+OsZFP轉化酵母菌株Y2H感受態細胞,分別涂布于SDO,SDO/X和SDO/X/A固體培養基,30℃培養3~5 d。結果顯示:SDO培養基中,含有pGBKT7與pGBKT7+OsZFP載體的酵母菌株單克隆大小相近(圖2A和2B),表明含有OsZFP基因的釣餌表達載體對酵母菌株的生長沒有毒性作用;同時,含有pGBKT7+OsZFP載體的酵母菌株在SDO/X固體培養基中呈白色(圖2C),且在SDO/X/A固體培養基中不能生長(圖2D),表明pGBKT7+OsZFP載體在酵母菌株Y2H中無自激活現象,適合互作蛋白的文庫篩選。
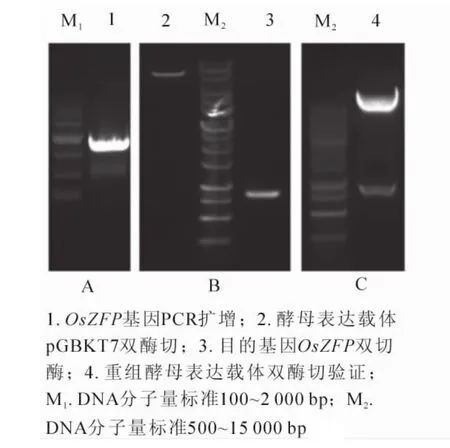
圖1 含OsZFP基因的酵母表達載體構建Figure 1 Construction of bait vector which containing OsZFP

圖2 OsZFP基因對酵母菌株Y2H的毒性及自激活驗證Figure 2 Testing bait vector which contained OsZFP for toxicity and autoactivation
2.3 互作蛋白的篩選與分離
將pGBKT7+OsZFP載體與水稻cDNA文庫共轉化酵母菌株Y2H感受態細胞,涂布于DDO/X/A固體培養基進行篩選;挑選候選陽性單克隆進一步接種至QDO/X/A固體培養基進行篩選。將候選陽性單克隆(83號和87號)轉接至DDO/X固體培養基2~3次,篩選分離出藍色(箭頭所示)陽性單克隆(圖3)。
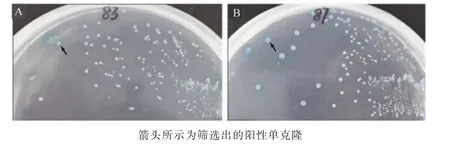
圖3 候選互作蛋白(83號和87號)在DDO/X固體培養基中的篩選Figure 3 Segregation of candidate positive interactors (blue) from non-interactors (white)
2.4 互作蛋白的分離和回復驗證
利用酵母質粒提取試劑盒提取DDO/X固體培養基中藍色單克隆表達質粒。將分離互作蛋白表達載體 pGADT7+83(AD-83)和 pGADT7+87(AD-87)分別與 pGBKT7(BD)或 pGBKT7+OsZFP(BD-ZFP)共轉化酵母菌株Y2H,涂布于DDO/X和QDO/X/A固體培養基進行篩選。結果顯示,BD-ZFP/AD-83和87以及BD/AD-83和87共轉化后,酵母Y2H均能在DDO/X固體培養基中生長;但是只有BD-ZFP/AD-87共轉化后酵母Y2H能在QDO/X/A固體培養基中生長且呈藍色(圖4),表明87號篩選蛋白可能為陽性互作蛋白。進一步對87號互作蛋白的文庫表達載體進行測序和同源性比對,鑒定為含有T-complex polypeptide 1(TCP-1)結構域的分子伴侶蛋白第7個亞基(LOC4341893),命名為chaperonin containing TCP-1 eta subunit(CCT-eta);該基因mRNA全長為2 123 bp,編碼560個氨基酸。

圖4 酵母雙雜交回復驗證Figure 4 Confirmation of positive interaction by yesat two-hybrid
3 討論
鋅指結構蛋白是一類含有指狀結構域的基因表達調控蛋白,依據蛋白結構中半胱氨酸和組氨酸殘基的數目和順序可以將鋅指結構分為C2H2,C2HC和C2C2等類型。在真核生物中,C2H2型鋅指結構蛋白的類型最多,研究也較為深入。水稻基因組中存在189個編碼含有QALGGH保守結構域的C2H2型鋅指結構轉錄因子,其中有18個基因是調控水稻穗發育,12個是種子特異表達基因[16];另外,含有C2H2型鋅指結構的水稻蛋白ZFP36被證實參與脫落酸誘導的抗氧化防御機制[17]。這些結果都表明:鋅指結構蛋白廣泛參與調控水稻的生長發育及生理生化途徑。
本研究以調控水稻側根生長發育的CCHC型鋅指結構蛋白為釣餌,通過酵母雙雜交技術從水稻cDNA文庫中篩選獲得2個互作蛋白;酵母雙雜交一對一回復驗證顯示BD/AD-87共轉化酵母在QDO培養基中不能生長,但BD-ZFP/AD-87能在QDO培養基中生長且呈藍色(圖4),表明87號蛋白為陽性互作蛋白。另外,BD/AD-83在QDO培養基中呈白色,但BD-ZFP/AD-83共轉化酵母在QDO培養基上不能生長,因此,83號篩選蛋白為假陽性。
含有TCP-1結構域的分子伴侶蛋白已被證實在真核生物細胞中參與蛋白的折疊與復性[18]。該蛋白至少由8個不同功能亞基組成,各個亞基同源性僅為30%;同時,各個蛋白亞基也含有相對保守的結構域,可能與ATP酶活性相關[19]。本研究篩選到的編碼CCT蛋白的第7個亞基與C2HC型鋅指結構蛋白存在互作,推測它可能參與鋅指結構蛋白的折疊與形成,從而調控水稻根系的生長發育,其相關生物學功能需要通過反向遺傳學進一步研究。
[1] KHUSH G S.What it will take to feed 5.0 billion rice consumers in 2030 [J].Plant Mol Biol,2005,59(1):1-6.
[2] KING J,GAY A,SYLVESTER-BRADLEY R,et al.Modelling cereal root systems for water and nitrogen capture:towards an economic optimum [J].Annu Bot,2003,91(3):383-390.
[3] ZHAO Yu,HU Yongfeng,DAI Mingqiu,et al.The WUSCHEL-related homeobox gene WOX11 is required to activate shoot-borne crown root development in rice [J].Plant Cell,2009,21(3):736-748.
[4] ZHU Zhenxing,LIU Yu,LIU Shaojun,et al.A gain-of-function mutation in OsIAA11 affects lateral root development in rice [J].Mol Plant,2012,5(1):154-161.
[5] KANG Bo,ZHANG Zhongchen,WANG Lingling,et al.OsCYP2,a chaperone involved in degradation of auxin-responsive proteins,plays crucial roles in rice lateral root initiation [J].Plant J,2013,74(1):86-97.
[6] COUDERT Y,PéRIN C,COURTOIS B,et al.Genetic control of root development in rice,the model cereal [J].Trends Plant Sci,2010,15(4):219-226.
[7] LUCHI S.Three classes of C2H2zinc finger proteins [J].Cell Mol Life Sci,2001,58(4):625-635.
[8] CIFTCIYILMAZ S,MILLTER R.The zinc finger network of plants [J].Cell Mol Life Sci,2008,65(7/8):1150-1160.
[9] GAN Yinbo,KUMIMOYO R,LIU Chang,et al.GLABROUS INFLORESCENCE STEMS modulates the regulation by gibberellins of epidermal differentiation and shoot maturation in Arabidopsis [J].Plant Cell,2006,18(6):1383-1395.
[10] GAN Yinbo,LIU Chang,YU Hao,et al.Integration of cytokinin and gibberellin signalling by Arabidopsis transcription factors GIS,ZFP8 and GIS2 in the regulation of epidermal cell fate [J].Development,2007,134(11):2073-2081.
[11] ZHOU Zhongjing,AN Lijun,SUN Lili,et al.Zinc finger protein 5 is required for the control of trichome initiation by acting upstream of ainc finger protein 8 in Arabidopsis [J].Plant Physiol,2011,157(2):673-682.
[12] AN Lijun,ZHOU Zhongjing,SUN Lili,et al.A zinc finger protein gene ZFP5 integrates phytohormone signaling to control root hair development in Arabidopsis [J].Plant J,2012,72(3):474-490.
[13] DEVAIAH B N,NAGARAJAN V K,RAGHOTHAMA K G.Phosphate homeostasis and root development in Arabidopsis are synchronized by the zinc finger transcription factor ZAT6 [J].Plant Physiol,2007,145(1):147-159.
[14] AGARWAL P,ARORA R,RAY S,et al.Genome-wide identification of C2H2zinc-finger gene family in rice and their phylogeny and expression analysis [J].Plant Mol Biol,2007,65(4):467-485.
[15] LIU Hongbo,GUO Xiang,NAEEM M S,et al.Transgenic Brassica napus L.lines carrying a two gene construct demonstrate enhanced resistance against Plutella xylostella and Sclerotinia sclerotiorum [J].Plant Cell Tiss Org Cult,2011,106(1):143-151.
[16] AGARWAL P,ARORA R,RAY S,et al.Genome-wide identification of C2H2zinc-finger gene family in rice and their phylogeny and expression analysis [J].Plant Mol Biol,2007,65(4):467-485.
[17] ZHANG Hong,LIU Yanpei,WEN Feng,et al.A novel rice C2H2-type zinc finger protein,ZFP36,is a key player involved in abscisic acid-induced antioxidant defence and oxidative stress tolerance in rice [J].J Exp Bot,2014,65(20):5795-5809.
[18] ZEBOL J R,HEWITT N M,MORETTI P A,et al.The CCT/TriC chaperonin is required for maturation of sphingosine kinase 1 [J].Int J Biochem Cell Biol,2009,41(4):822-827.
[19] KUBOTA B H,HYNES G,WILLISON K.The chaperonin containing t-complex polypeptide 1 (TCP-1) multisubunit machinery assisting in protein folding and assembly in the eukaryotic cytosol[J].Eur J Biochem,1995,230(1):3-16.
Screening and identification of a protein for controlling lateral root development in Oryza sativa
ZHENG Xiuwen1,CUI Peng1,SHI Jiawei1,YANG Jingjing1,CHEN Minmin2,ZHENG Yao2,XU Ling2,LIU Hongbo1
(1.School of Agricultural and Food Sciences,Zhejiang A&F University,Hangzhou 311300,Zhejiang,China;2.School of Life Science,Zhejiang Sci-Tech University,Hangzhou 310018,Zhejiang,China)
OsZFP,a CCHC-type zinc finger protein,has reportedly participated in lateral root development in rice.To determine the molecular mechanism and its interacting proteins in a regulated network,the OsZFP gene from Oryza sativa. ‘Nipponbare’ was cloned,and then a yeast bait vector,pGBKT7+OsZFP,was constructed by EcoRI and SalI for cDNA library screening of rice.Results showed no toxicity or autoactivation with transformation of the Y2H strain containing the pGBKT7+OsZFP bait vector.In a yeast two-hybrid assay,a positive interacted protein was obtained and confirmed from the cDNA library.Sequencing and homologous alignment from the National Center for Biotechnology Information (NCBI) showed that the interacting protein(named chaperonin containing a T-complex polypeptide 1 (TCP-1) eta subunit,CCT-eta)was the eta subunit of the chaperone which contained a TCP-1 domain.Based on the interaction between OsZFP and the CCT-eta subunit,it was speculated that the CCT-eta subunit of the chaperone protein may be involved in controlling lateral root development in rice. [Ch,4 fig.19 ref.]
botany;Oryza sativa;regulated protein;vector construction;yeast two-hybrid;protein interaction
S511;Q943.2
A
2095-0756(2017)06-1024-05
10.11833/j.issn.2095-0756.2017.06.008
2016-10-30;
2016-12-12
國家自然科學基金青年基金資助項目(31301272);浙江農林大學人才啟動基金資助項目(2013FR022);國家級大學生創新創業訓練計劃項目(201510341001);浙江省高校生物學重中之重(一級)學科開放基金資助項目(2015D19)
鄭秀文,從事水稻根系發育基因功能研究。E-mail:1327158979@qq.com。通信作者:劉宏波,副教授,博士,從事作物水稻根系發育與逆境相關基因功能研究。E-mail:hbliu@zafu.edu.cn
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
