土壤干旱對2個沙棘品種葉片水勢和光合特性的影響
2017-11-27 07:30:28高國日劉娟娟陳道國董禮隆何彩云
浙江農林大學學報 2017年6期
高國日,劉娟娟,陳道國,董禮隆,何彩云
(1.中國林業科學研究院 林業研究所 國家林業局林木培育重點實驗室,北京 100091;2.中國林業科學研究院 沙漠林業實驗中心,內蒙古 磴口015200)
土壤干旱對2個沙棘品種葉片水勢和光合特性的影響
高國日1,劉娟娟1,陳道國1,董禮隆2,何彩云1
(1.中國林業科學研究院 林業研究所 國家林業局林木培育重點實驗室,北京 100091;2.中國林業科學研究院 沙漠林業實驗中心,內蒙古 磴口015200)
為了研究干旱脅迫對沙棘Hippophae rhamnoides生長的影響,通過模擬土壤自然干旱和復水過程,研究不同程度土壤干旱下中國沙棘H.rhamnoides subsp.sinensis品種 ‘豐寧’ ‘Fengning’和蒙古沙棘H.rhamnoides subsp.mongolia品種 ‘向陽’ ‘Xiangyang’2個品種葉片水勢和光合特性。結果表明:蒙古沙棘 ‘向陽’在控水第22天(19.40%)左右出現葉片干縮,而中國沙棘 ‘豐寧’在第33天(11.66%)左右出現,說明 ‘豐寧’的抗旱性要高于‘向陽’。隨著土壤體積含水量的降低,2個沙棘品種的葉片水勢下降,增加了對水分的吸收能力,并且 ‘豐寧’下降幅度要大于 ‘向陽’。與光合生理相關的最大凈光合速率、蒸騰速率、氣孔導度、光飽和點、表觀量子效率均隨著干旱程度加深而下降,而葉綠素相對含量和光補償點出現上升,水分利用效率先上升后下降, ‘豐寧’試驗組的暗呼吸速率出現先下降后恢復趨勢,而 ‘向陽’試驗組先不變后上升。復水后各生理指標出現不同程度的恢復。因此,在人工林的培育過程中出現極度干旱,沙棘葉片干縮,要及時灌溉,以免對沙棘生長造成不可恢復的傷害。‘豐寧’比 ‘向陽’更適合作為選育良種的親本。圖3表3參37
植物學;沙棘;干旱脅迫;水分關系;光合生理
沙棘Hippophae rhamnoides為胡頹子科Elaeagnaceae沙棘屬Hippophae植物,是一種廣泛分布于亞洲、歐洲和北美洲的落葉樹種。在中國,沙棘天然分布于華北、西北、西南等半干旱地區,后經人工引種至東北地區[1]。沙棘不但適應性好,固氮能力強[2],具有良好的改良土壤和水土保持功能,而且沙棘果實及葉子中富含的多種營養成分和生物活性物質,具有醫藥及日化等工業前景[3-5],是一種生態效益、社會效益與經濟效益均十分顯著的優良樹種[6]。雖然沙棘在長期的進化中已經具備多種抗旱的結構特性和生理機制,但是水分仍然是沙棘生長和發揮功能的主要限制因子。所以,培育生態價值和經濟價值高的沙棘品種依然是林業生態建設的焦點。沙棘品種的抗旱性對于其價值的實現具有重要作用。一些學者不僅對沙棘的抗旱表型進行了研究,還探討了干旱脅迫下沙棘光合作用和生理生化等特性的變化[7-9],并以這些指標作為抗旱性強弱的評價依據,對沙棘品種間的抗旱性進行了比較[10-14]。但其試驗方案多采用人為控制土壤含水量不變的方法,不能保證補充散失水分過程中土壤水分均衡和模擬自然環境中動態的土壤干旱過程,同時缺少干旱脅迫后復水過程。本研究減少人為對水分的干預,模擬自然環境中土壤干旱及復水過程,觀察中國沙棘 ‘豐寧’H.rhamnoides subsp.sinensis‘Fengning’和引進品種 ‘向陽’H.rhamnoides subsp.mongolia‘Xiangyang’在不同土壤含水量的表型變化,以及對不同干旱程度的植株水分關系和光合生理特性,通過數據分析,比較2個品種對水分變化響應的差異性和共性,從而為沙棘抗旱樹種的篩選和育林的補水時間提供科學依據。
1 材料與方法
1.1 試驗材料
本試驗使用的材料采自中國林業科學研究院沙漠林業實驗中心,為2年生沙棘根蘗苗,分別是鄉土品種中國沙棘 ‘豐寧’和引進蒙古品種大果沙棘 ‘向陽’。2016年4月將苗木栽入高22.5 cm,內徑25.5 cm 的花盆中, 1 株·盆-1, 盆中裝土 5.5 kg[m(沙):m(草炭):m(珍珠巖):m(黏土)=5:3:1:1], 土壤最大含水量為62%,體積含水量44%。放置于中國林業科學研究院科研溫室大棚,生長條件為自然光照,晝/夜溫度為20~30℃/10~15℃。相對濕度為60%~70%[15],使植株正常生長。3個月后選取健康,生長均一的植株分組進行干旱處理試驗。
1.2 試驗方法
本試驗共分為2個組,分別為對照組(C)和試驗組(T)。2016年7月30日前所有供試苗木保持正常水分,第2天測定供試苗木正常水分條件下的土壤體積含水量,此后試驗組停止澆水進行自然干旱脅迫。對照組為正常澆水組,水分控制在(35.00±5.00)%土壤體積含水量。土壤含水量采用WET土壤三參數速測儀Moisture Meter-HH2(Delta-T Devices Ltd,英國)[16]測定,隔1 d測量1次,及時補充對照組散失的水分。隔5 d進行各項生理指標測定[17]。達到沙棘最低耐受程度后,對試驗組復水至對照組水平,第3天和第5天測定生理指標。
1.3 測定指標及方法
在試驗期間,用LI-6400便攜式光合儀(LI-COR,美國)分別測定對照組與試驗組2個沙棘品種的凈光合速率(Pn, μmol·m-2·s-1), 蒸騰速率(Tr, mmol·m-2·s-1), 氣孔導度(Cond, mol·m-2·s-1)和光響應曲線。測定條件:紅藍固定光源,固定光強為1 000 μmol·m-2·s-1,溫度28℃,相對濕度為70%,二氧化碳濃度為溫室中大氣濃度。光響應曲線使用自動測量程序,梯度為2 000,1 800,1 500,1 000,600,400, 200, 100, 50, 0(μmol·m-2·s-1), 同時用葉綠素含量測定儀(SPAD-502Plus, 日本)[18]測定相應位置的葉綠素相對含量,選擇15片葉片分成3組,測量記錄每組平均值。葉片水勢變化用WP4露點水勢儀(Decagon, 美國)[19]測定。
1.4 統計分析
利用Excel 2007統計軟件整理數據及制作圖表。在SPSS 17.0統計軟件中進行one-way ANOVA分析和0.05,0.01水平的最小顯著性差異(LSD)法多重分析。
2 結果分析

表1 2個沙棘品種不同控水天數葉片水勢的變化Table 1 Changes of leaf water potential of two Hippophae rhamnoides cultivars in different days
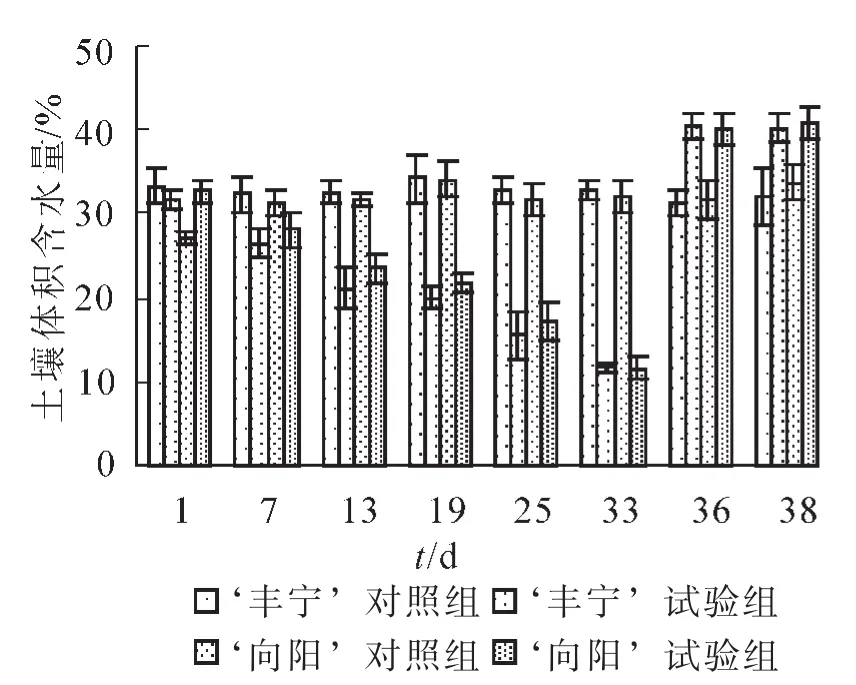
圖1 控水后土壤體積含水量變化Figure 1 Changes of soil volumetric water content after the water control

圖2 控水后葉綠素相對含量變化Figure 2 Changes of chlorophyll relative content after the water control
2.1 土壤體積含水量的變化
本研究將土壤體積含水量作為土壤干旱的分級標準[20],控水后的土壤體積含水量的變化如圖1所示。 ‘豐寧’對照組和 ‘向陽’對照組的體積含水量保持在最大體積含水量75%~80%的正常水分條件,而 ‘豐寧’試驗組和 ‘向陽’試驗組的體積含水量呈現下降趨勢并且趨勢相同。在自然干旱第7天,2個試驗組分別26.37%和28.09%,達到輕度干旱。干旱至第19天,體積含水量分別為19.90%和21.78%,為中度干旱。在第22天觀測到 ‘向陽’有葉片開始變黃,體積含水量約為19.40%。第33天開始對試驗組復水至澆透,復水前體積含水量為11.66%和11.63%,為重度干旱,達到 ‘豐寧’所能耐受的最低含水量。在該水分下植株葉片開始變黃、枯萎。
2.2 不同程度干旱脅迫對2個沙棘品種植株葉片水勢的影響
對 ‘豐寧’和 ‘向陽’不同控水天數葉片水勢的測定(表1)發現, ‘豐寧’試驗組與 ‘向陽’試驗組與對照組相比葉片水勢降低,其中 ‘豐寧’試驗組下降明顯,在第13天達到顯著差異水平(P<0.05),第19天達到極顯著水平(P<0.01)。 ‘向陽’試驗組下降趨勢緩慢,在第25天達到顯著差異,第33天達到極顯著水平(P<0.01)。在重度干旱情況下,雖然 ‘豐寧’差值(0.77)高于‘向陽’(0.45),但是 ‘豐寧’試驗組 (-2.36)和 ‘向陽’試驗組(-2.31)的大小差異不顯著。復水后沙棘葉片水勢快速上升,在第3天, ‘向陽’恢復正常水平,而 ‘豐寧’第5天才恢復。
2.3 干旱脅迫對光合生理的影響
2.3.1 不同程度干旱脅迫對2個沙棘品種光合特性的影響 從圖2可以看出: ‘豐寧’對照組和 ‘向陽’對照組葉綠素相對含量總體呈現穩定趨勢,但 ‘豐寧’對照組在后期有微量上升。兩者相比, ‘向陽’要高于 ‘豐寧’。 ‘豐寧’試驗組和 ‘向陽’試驗組葉綠素相對含量分別要高于對照組,在總體趨勢上隨著干旱程度增強上升,在第33天重度干旱下達到最大增長幅度,分別是10.33%和2.39%,干旱脅迫對 ‘豐寧’葉綠素相對含量的影響高于 ‘向陽’。復水后 ‘向陽’下降至對照組水平,而 ‘豐寧’仍高于對照組。
綠色植物主要的生命活動之一是光合作用,干旱脅迫對光合作用有重要影響。從表2可以看出:隨著控水時間延長,干旱程度增強,凈光合速率(Pn),蒸騰速率(Tr),氣孔導度(Cond)總體呈現出下降趨勢,復水后恢復或者超過正常水平。 ‘豐寧’試驗組和 ‘向陽’試驗組與對照組相比,在輕度干旱和中度干旱階段Pn下降趨勢不明顯,在控水后第33天達到重度干旱,都降到最低值,分別是對照組的32.2%和34.6%,具有顯著差異(P<0.05), ‘向陽’是 ‘豐寧’的1.4倍。復水第2天, ‘向陽’的Pn稍微上升,復水第3天,超過對照組。而 ‘豐寧’在復水第3天才出現上升,第5天與對照組沒有顯著差異; ‘豐寧’試驗組的Tr呈現下降趨勢,與對照組的差異逐漸增加,第25天土壤體積含水量為15.51%時,具有顯著差異(P<0.05),在第33天達到最低值,為對照組的16.38%。復水后開始上升,但沒有恢復正常水平。 ‘向陽’試驗組變化趨勢與 ‘豐寧’試驗組大體相同,但 ‘向陽’試驗組的下降趨勢更平緩,在第33天達到顯著差異水平 (P<0.05),為對照組的48.02%。復水后第3天超過正常水平; ‘豐寧’試驗組和 ‘向陽’試驗組的Cond變化趨勢是隨著干旱程度增強而下降,重度干旱程度具有顯著差異,分別是對照組的13.75%和36.89%,復水后上升,但是 ‘向陽’試驗組迅速恢復,而 ‘豐寧’試驗組則需要更長時間。Pn與Tr比值為植物水分利用效率(EWU),沙棘在不同的土壤水分條件和生長時期,EWU有很大差異。隨著干旱時間的延長,實驗組與對照相比先有較大上升,隨后出現下降,其中在中度干旱下沙棘的EWU最高, ‘豐寧’試驗組與 ‘豐寧’對照組相比增加35%,而 ‘向陽’試驗組與 ‘向陽’對照組相比增加31%。重度干旱下沙棘生長受抑制,物質積累減少,因此EWU降低。
2.3.2 不同程度干旱脅迫對2個沙棘品種光響應曲線及特征參數的影響 為了全面闡明不同干旱脅迫程度對植物光合作用的影響,除了測定了上述的光合生理特性,還測定了不同控水階段的光響應曲線。其結果如圖3所示。對光響應曲線擬合后,利用分段式函數[21]計算出凈光合最大速度(Pnmax),光飽和點(PLS),表觀量子效率(EAQ),光補償點(PLC),暗呼吸速率(Rd)數值,如表3所示。由圖3可以看出:所有的光響應曲線呈現二次方程的形式,光合有效輻射為0~400 μmol·m-2·s-1,Pn隨著光合有效輻射增加而增加,增幅較大。光照有效輻射進一步增加,Pn增幅變緩,增加到PLS后,Pn不再增加,符合光響應曲線的一般規律。從表3中看出在第13天輕度干旱階段, ‘豐寧’試驗組和 ‘向陽’試驗組與對照組相比,Pnmax,PLS,EAQ都出現下降趨勢,而PLC上升。 ‘豐寧’試驗組的Rd出現先下降后恢復趨勢,而‘向陽’試驗組出現先不變后上升。隨著控水時間延長,第33天重度干旱下降趨勢更加明顯,差異更大。復水第8天后, ‘豐寧’試驗組的Pnmax和PLS超過對照組,EAQ,PLC和Rd與對照組大體一致。而‘向陽’試驗組的Pnmax和PLC沒有恢復到正常水平,其他恢復正常。

表2 2個沙棘品種不同控水天數光合生理特性的變化Table 2 Changes of photosynthetic characteristic of two Hippophae rhamnoides cultivars in different days

圖3 控水第13天、第33天和復水第8天的光響應曲線Figure 3 Light response curve of 13,33 d after water control and 8 d after rehydration
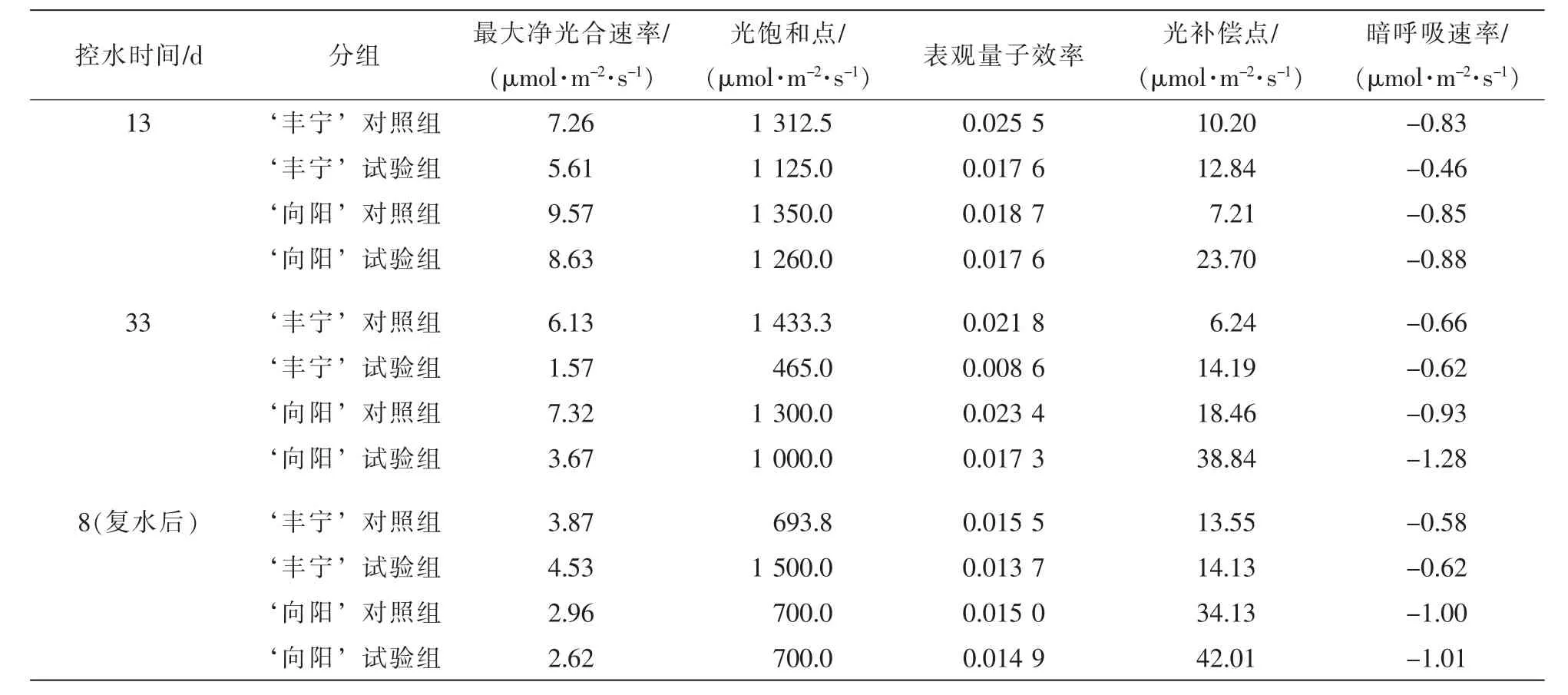
表3 2個沙棘品種不同控水天數光響應特性值的變化Table 3 Changes of response characteristics to light of two Hippophae rhamnoides cultivars in different days
3 討論
在沙棘的生長區,降水不足會導致沙棘生長量和生物量降低[22],水分成為制約沙棘正常生長的主導因素。本研究模擬自然環境中無降水的土壤水分變化,同時對土壤水分進行連續監測。由于溫室濕度大導致水分蒸發速率慢,同時沙棘對水分利用率高,對水分吸收較少,所以土壤水分下降緩慢,為植物適應不同強度的土壤干旱提供了時間。在該試驗土質和生長條件下, ‘向陽’在第22天出現干縮現象,體積含水量約為19.40%,而 ‘豐寧’在連續干旱33 d后出現同樣現象,體積含水量為11.66%,與吳林等[23]在田間觀察到的現象一致。
葉片水勢代表植物水分運輸的能力,是廣泛應用于衡量植物抗旱的生理指標,能夠反映植物吸收水分的能力和水分狀況好壞[24],影響葉水勢的因素很多,光照條件、水分供應情況都會對水勢大小產生影響,并且同一天不同時間段不同部位的葉片水勢也會不同[25]。隨著土壤水分降低,植物體內水分產生虧缺,植物為了滿足自身生命活動所需水分,降低葉片水勢,增加對土壤水分的吸收,抵御外界的干旱條件[26]。 ‘豐寧’比 ‘向陽’水勢低,差值大,說明2個沙棘品種對水分的吸收能力和利用效率有差異。
植物葉片中葉綠素相對含量高低、葉綠素a和葉綠素b的比值直接影響植物光合作用的強弱[27]。葉綠素的合成和降解與外界環境相關。干旱脅迫下,部分植物葉片中的葉綠素相對含量會增加,表現一定抗旱性[28]。沙棘葉綠素相對含量呈現出穩定或者微量上升的趨勢,與韓蕊蓮等[29]得到的結論相同。說明在遭受干旱脅迫時,2個沙棘品種為保持較高的生長速率,依舊維持較高的葉綠素含量, ‘豐寧’的作用要明顯于 ‘向陽’。
Pn能直接反映光合作用的強度,是衡量光合作用的重要指標,對植物自身來說,Pn大小受Tr和Cond等多因素影響,蒸騰作用可以顯示植物體內水分生理特征,Tr反應水分散失情況。Cond一定程度上反映植物內代謝水平,靈敏度可以作為抗旱特征[30]。它們共同保障植物光合作用的順利進行。Tr和Cond下降,同時Pn出現下降,說明植物水分缺失會影響Tr和Cond,阻礙光合作用的順利進行[31]。通過EWU的變化,更能直接表現出沙棘的有機物積累變化趨勢。EWU在中度干旱下最高,其原因是在中度水分虧缺下,耗水量大幅下降,但新枝生長量下降幅度小。而長時間的重度干旱使沙棘生長受抑制,EWU同樣很低。植物對于逆境環境具有抵抗性,在復水的短期內會出現超過正常水平現象,這是植物的防御機制所造成的。
通過測定光響應曲線,發現干旱會對Pnmax,PLS,EAQ,PLC影響顯著,PLC上升,EAQ下降說明干旱影響了植物對弱光的利用效率,而PLS下降說明干旱抑制了對光照的利用,在沒有達到對照組的光飽和點時水分變成了植物有機質積累的限制因子。干旱對Rd的影響表現出品種差異性, ‘豐寧’Rd下降,降低了有機物的消耗,有利于在較低的光合速率下保持生長。復水激發了植物的生理補償效應,植物表現出很強的恢復力,在很短的時間內恢復或超過正常水平[32],但如果干旱對植物造成不可恢復傷害,即使復水,植物也將恢復不到正常水平。在本試驗中,復水后, ‘豐寧’和 ‘向陽’的葉片水勢快速恢復正常。 ‘向陽’葉綠素相對含量降低至對照水平,而 ‘豐寧’仍高于對照,其他光合特性指標基本恢復正常。由于測定時間較長,跨度較大,出現季節性變化,導致不同時期的對照組指標也出現下降的趨勢。
對沙棘抗旱機制的研究不能只停留在生理水平,植物對干旱脅迫的反應主要受基因型和基因表達的影響,而表觀遺傳修飾在抗逆基因表達調控過程中起到不可忽視的作用[33]。許多研究表明,DNA甲基化程度高,基因表達受到抑制,組蛋白乙酰化水平高促進基因表達[34-35],兩者相互協調,從而實現對一些關鍵抗逆基因的準確調控。通過比較干旱脅迫前后DNA甲基化和組蛋白乙酰化水平的變化[36-37],為闡明DNA甲基化和組蛋白遺傳修飾在沙棘抗旱基因表達調控中的功能具有重要的意義。
4 結論
在干旱脅迫下, ‘豐寧’和 ‘向陽’沙棘形態特征、葉片水勢和光合生理都表現出明顯的變化,真實反映了沙棘在自然干旱環境中的變化。沙棘雖是強抗旱植物,但其抗旱能力有限,在人工林的培育過程中,在極度干旱情況下(土壤體積含水量為15%左右),沙棘葉片出現干縮,要及時灌溉,以免對沙棘生長造成不可恢復的傷害。通過葉片水勢和光合生理等比較, ‘豐寧’比 ‘向陽’具有更強的耐旱性,可以將其作為選育良種的親本。
[1] 安寶利,盧順光.沙棘種質資源保護和利用的現狀與展望[J].國際沙棘研究與開發,2004(2):12-15.AN Baoli,LU Shunguang.Conservation and utilization of germplasm resources of seabuckthorn (Hippophae rhamnoides L.) [J].Glob Seabuckthorn Res Dev,2004 (2):12-15.
[2] LIU Xiping,GONG Chunming,FAN Yangyang,et al.Response pattern of amino compounds in phloem and xylem of trees to soil drought depends on drought intensity and root symbiosis [J].Plant Biol,2013,15(suppl 1):101-108.
[3] ANDERSSON S C,OLSSON M E,JOHANSSON E,et al.Carotenoids in sea buckthorn (Hippophae rhamnoides L.)berries during ripening and use of pheophytin a as a maturity marker [J].J Agric Food Chem,2008,57 (1):250-258.
[4] LEE H I,KIM M S,LEE K M,et al.Anti-visceral obesity and antioxidant effects of powdered sea buckthorn (Hippophae rhamnoides L.) leaf tea in diet-induced obese mice [J].Food Chem Toxicol,2011,49(9):2370-2376.
[5] PATEL C A,DIVAKAR K,SANTSNI D,et al.Remedial prospective of Hippopha? rhamnoides Linn.(sea buckthorn)[J].ISRN Pharmacol,2012,doi:10.5402/2012/436857.
[6] 田魏龍,蔣志榮.不同沙棘品種對干旱脅迫的生理生化響應[J].中國沙漠,2011,31(5):1215-1220.TIAN Weilong,JIANG Zhirong.Physiological and biochemical response of six cultivars of Hippophae rhamnoides to drought stress [J].J Desert Res,2011, 31(5): 1215-1220.
[7] 阮成江,李代瓊.半干旱黃土丘陵區沙棘的光合特性及其影響因子[J].沙棘,2007,20(4):6-9.RUAN Chengjiang,LI Daiqiong.Photosynthetic characteristics of Hippophae rhamnoides L.and its influencce factors in semiarid loess lilly-region [J].Hippophae,2007,20(4): 6-9.
[8] 陳海玲,徐軍,劉芳,等.不同品種沙棘光合生理特性研究[J].沈陽農業大學學報,2015,46(1):110-113.CHEN Hailing,XU Jun,LIU Fang,et al.Photosynthetic and physiological characteristics of seabuckthorn cultivars[J].J Shenyang Agric Univ,2015, 46(1):110-113.
[9] 陳海玲,劉芳,羅紅梅,等.烏蘭布和沙區不同沙棘品種葉片及果實特性研究[J].林業資源管理,2014(3):122-125.CHEN Hailing,LIU Fang,LUO Hongmei,et al.Study on characteristics of leaf and berry of different cultivars of seabuckthorn in Ulan Uuh Desert[J].For Resour Manage,2014(3): 122-125.
[10] 張李華.俄羅斯大果沙棘與中國沙棘的干旱適應性比較研究[J].中國林副特產,2010(3):25-27.ZHANG Lihua.Study on drought adaptability between Hippophae rhamnoides ssp.Russia and Hippophae rhamnoides subsp.sinensis [J].For By-prod Spec China,2010(3): 25-27.
[11] 馬玉花,高英,向前勝,等.2種青海野生沙棘對干旱脅迫的響應及其抗旱性評價[J].湖北農業科學,2014, 53(16): 3827-3831.MA Yuhua,GAO Ying,XIANG Qiansheng,et al.The response to drought stress and drought-resistance evaluation of two species of wild seabuckthorn in Qinghai Provice [J].Hubei Agric Sci,2014, 53(16): 3827-3831.
[12] 何彩云,李夢穎,羅紅梅,等.不同沙棘品種抗旱性的比較[J].林業科學研究,2015,28(5):634-639.HE Caiyun,LI Mengying,LUO Hongmei,et al.Comprehensive evaluation on drought resistance of Hippophea rhamnoides [J].For Res,2015, 28(5): 634-639.
[13] 趙延霞,駱有慶,宗世祥,等.不同沙棘品種雌雄株葉片解剖結構及抗旱性比較[J].北京林業大學學報,2012, 34(6): 34-41.ZHAO Yanxia,LUO Youqing,ZONG Shixiang,et al.Comparison in leaf anatomical structure and drought resistance of different sex and varieties of sea buckthorn [J].J Beijing For Univ,2012, 34(6): 34-41.
[14] 王洪江,劉麗娜.3個優良沙棘品種抗旱性初步研究[J].遼寧林業科技,2013(1):23-24,61.WANG Hongjiang,LIU Lina.A preliminary study on the drought resistance of three advance Hippophea rhamnoides cultivars [J].J Liaoning For Sci Technol,2013(1): 23-24, 61.
[15] 徐潔,白坤棟,萬賢崇,等.歐美楊107楊抗硫無性系的變異及其抗氧化保護系統對SO2響應[J].林業科學,2011, 47(2): 66-71.XU Jie,BAI Kundong,WAN Xianchong,et al.Responses of poplar(Populus× euramericana ‘74/76’ )SO2-resistant clone to SO2fumigation and the variation in antioxidant systems [J].Sci Silv Sin,2011, 47(2): 66-71.
[16] 甘卓亭,劉文兆.HH2型土壤濕度計測定土壤含水量的標定研究[J].干旱地區農業研究,2005,23(4):69-72.GAN Zhuoting,LIU Wenzhao.Calibration on measurement of soil water content using SMS2 soil moisture probe and HH2 meter[J].Agric Res Arid Areas,2005, 23(4): 69-72.
[17] 李延生,王長平,張大偉,等.干旱脅迫對美國白蠟幼苗生理指標的影響[J].吉林林業科技,2016,45(2):24-26,62.LI Yansheng,WAGN Changping,ZHANG Dawei,et al.The effect of drought stress on physiological indexes of Frax-inus americana seedlings [J].J Jilin For Sci Technol,2016, 45(2):24-26, 62.
[18] 朱進.溫室不同黃瓜幼苗葉綠素熒光參數和葉綠素相對含量的變化[J].長江蔬菜,2012(2):36-38.ZHU Jin.Changes of fluorescence chlorophyll parameters and relative level of chlorophyll in different cucumber seedlings in greenhouse [J].J Changjiang Vegetab,2012(2): 36-38.
[19] 晏欣,龔吉蕊,張梓瑜,等.狼針草光合特性對放牧干擾的響應[J].植物生態學報,2013,37(6):530-541.YAN Xin,GONG Jirui,ZHANG Ziyu,et al.Responses of photosynthetic characteristics of Stipa baicalensis to grazing disturbance [J].Chin J Plant Ecol,2013, 37(6): 530-541.
[20] 劉珊,何茜,李吉躍,等.石漠化樹種圓葉烏桕對干旱脅迫的生理響應[J].華南農業大學學報,2016,37(2): 96-100.LIU Shan,HE Qian,LI Jiyue,et al.Physiological responses of the limestone endemic plant Triadica rotundifolia seedlings to drought stress [J].J South China Agric Univ,2016, 37(2): 96-100.
[21] 段愛國,張建國.光合作用光響應曲線模型選擇及低光強屬性界定[J].林業科學研究,2009,22(6):765-771.DUAN Aiguo,ZHANG Jianguo.Selection of models of photosynthesis in response to irradiance and definition of attribute of weak light[J].For Res,2009, 22(6):765-771.
[22] 代光輝,李根前,李甜江,等.水分條件對中國沙棘生長及種群穩定性的影響[J].西北林學院學報,2011,26(2):1-8.DAI Guanghui,LI Genqian,LI Tianjiang,et al.Effect s of water conditions on the growth and population stability of Hippophae rhamnoides subsp.sinensis [J].J Northwest For Univ,2011, 26(2):1-8.
[23] 吳林,李亞東,郝瑞,等.水分逆境對沙棘葉片脯氨酸、過氧化物酶、過氧化氫酶等的影響[J].沙棘,1996, 9(3):15-18.WU Lin,LI Yadong,HAO Rui,et al.The influence of proline,peroxidase and catalase in leaves of Hippophae rhamnoides under drought stress [J].Hippophae,1996, 9(3):15-18.
[24] 梁宗鎖,王俊峰.簡述沙棘抗旱性及其耗水特性研究現狀[J].沙棘,1997,10(1):28-31.LIANG Zongsuo,WANG Junfeng.The research status of drought resistance and water consumption characteristics of Hippophae rhamnoides [J].Hippophae,1997, 10(1): 28-31.
[25] 劉青柏,劉明國,孟函寧,等.半干旱地區5種沙棘幼苗水分生理特征研究[J].林業資源管理,2011(5):101-106.LIU Qingbo,LIU Mingguo,MENG Hanning,et al.Study on water physiological characteristics of the seedlings of five Hippophae rhamnoides varieties in semi-arid area [J].For Resour Manage,2011(5):101-106.
[26] 韓蕊蓮,梁宗鎖,侯慶春,等.黃土高原適生樹種苗木的耗水特性[J].應用生態學報,1994,5(2):210-213.HAN Ruilian,LIANG Zongsuo,HOU Qingchun,et al.Water consumption properties of adaptable nursery stocks on Loess Pleteau [J].Chin J Appl Ecol,1994, 5(2):210-213.
[27] 林陽,王世忠.4種油松混交灌木樹種的耐陰性研究[J].河北林果研究,2014,29(3):258-262.LIN Yang,WANG Shizhong.Research on the shade tolerance of 4 shrub tree species mixed with Chinese pine [J].Hebei J For Orchard Res,2014, 29(3):258-262.
[28] 馬玉花,高英,周至遠,等.干旱脅迫對肋果沙棘幼苗生理指標及膜特性的影響[J].青海大學學報(自然科學版), 2013, 31(2):1-5.MA Yuhua,GAO Ying,ZHOU Zhiyuan,et al.Effects of drought stress on physiological characteristics of Hipphae neu-rocpa [J].J Qinghai Univ Nat Sci Ed,2013, 31(2):1-5.
[29] 韓蕊蓮,梁宗鎖,鄒厚遠.4個樹種在干旱下的生理適應性研究[J].西北林學院學報,1991,6(4):23-28.HAN Ruilian,LIANG Zongsuo,ZOU Houyuan.Physiological adaptability to drought of four woody species [J].J Northest For Coll,1991, 6(4):23-28.
[30] 溫瑀,鞏如英,徐迎碧,等.沙棘抗旱生理生態特征研究進展[J].民營科技,2008(5):117-118.WEN Yu,GONG Ruying,XU Yingbi,et al.The review of physiological and ecological characteristics of Hippophae rhamnoides in drought-stress [J].Private Sci Technol,2008(5):117-118.
[31] 唐道鋒,賀康寧,朱艷艷,等.白榆沙棘光合生理參數與土壤含水量關系研究[J].水土保持研究,2007,14(1):230-233.TANG Daofeng,HE Kangning,ZHU Yanyan,et al.Study on the relation between the photosynthetic physiological properties of Ulmus pumila and Hippophae rhamnoides and soil water content[J].Res Soil Water Conserv,2007, 14(1):230-233.
[32] 張向娟,張亞黎,張天宇,等.棉花長期干旱后復水葉片光合特性及碳氮含量的變化[J].石河子大學學報(自然科學版), 2014, 32(2):138-142.ZHANG Xiangjun,ZHANG Yali,ZHANG Tianyu,et al.Change of rewatering after long-term drought stress on photosynthesis and content of carbon and nitrogen of cotton [J].J Shihezi Univ Nat Sci,2014, 32(2):138-142.
[33] CHINNUSAMY V,ZHU Jiankang.Epigenetic regulation of stress responses in plants [J].Curr Opin Plant Biol,2009,12(2):133-139.
[34] FUKUDA H,SANO N,MUTO S,et al.Simple histone acetylation plays a complex role in the regulation of gene expression [J].Brief Funct Gen Prot,2006,5(3):190-208.
[35] ZHOU Junli,WANG Xiangfeng,HE Kun,et al.Genome-wide profiling of histone H3 lysine 9 acetylation and dimethylation in Arabidopsis reveals correlation between multiple histone marks and gene expression [J].Plant Mol Biol,2010,72(6):585-595.
[36] KIM J M,TO T K,ISHIDA J,et al.Alterations of lysine modifications on the histone H3 N-tail under drought stress conditions in Arabidopsis thaliana [J].Plant Cell Physiol,2008,49(10):1580-1588.
[37] GARG R,CHEVALA V V S N,SHANKAR R,et al.Divergent DNA methylation patterns associated with gene expression in rice cultivars with contrasting drought and salinity stress response [J].Sci Rep,2015,5: 1-16.doi:10.1038/srep14922.
Leaf water potential and photosynthetic characteristics of two Hippophae rhamnoides cultivars with soil drought
GAO Guori1,LIU Juanjuan1,CHEN Daoguo1,DONG Lilong2,HE Caiyun1
(1.Key Laboratory of Tree Breeding and Cultivation of State Forestry Administration,Research Institute of Forestry,Chinese Academy of Forestry,Beijing 100091,China;2.Experimentat Center of Desert Forestry,Chinese Academy of Forestry,Dengkou 015200,Inner Mongolia,China)
To study the effects of drought stress on the growth of Hippophae rhamnoides in a natural environment,plant water relations and photosynthetic characteristics of two cultivars:H.rhamnoides subsp.sinensis‘Fengning’ and H.rhamnoides subsp.mongolia ‘Xiangyang’ with different degrees of drought were studied by simulating natural soil drought conditions and water recovery processes.Results showed that leaves,experiencing drought treatment,initially shrunk at 22nd (19.40%) for the exotic species ‘Xiangyang’ and 33rd(11.66%) for the native species ‘Fengning’.Also, ‘Fengning’ had stronger drought resistance compared to‘Xiangyang’.In addition,with increasing drought stress,the leaf water potential of both species decreased(0.77 and 0.45) and the moisture absorption ability increased.The maximum net photosynthetic rate,net photosynthetic rate,transpiration rate,stomatal conductance,light saturation point,and apparent quantum yield decreased with increasing drought stress;whereas,the relative content of chlorophyll and the light compensation point increased.Also,water efficiency rose after falling.The dark respiration rate of ‘Fengning’ treatment group decreased first and then recovered,but ‘Xiangyang’treatment group was unchanged and then in-creased.After rehydration,the physiological indexes appeared in different degrees of recovery.Therefore,when extreme drought occurred in the process of cultivating a plantation,the leaves shrunk and need timely irrigation to avoid unrecoverable damage with ‘Fengning’ being more suitable than ‘Xiangyang’ as a breeding parent.[Ch,3 fig.3 tab.37 ref.]
botany;Hippophae rhamnoides;drought stress;water relations;photosynthetic physiology
S718.4
A
2095-0756(2017)06-0999-09
10.11833/j.issn.2095-0756.2017.06.005
2016-12-05;
2017-02-20
國家林業局林業公益性行業科研專項(201504103)
高國日,從事樹木逆境生理及分子生物學研究。E-mail:ggrjiayou@126.com。通信作者:何彩云,副研究員,博士研究生,從事樹木逆境生理及分子生物學研究。E-mail:hecy@caf.ac.cn
