mIDH1真核表達載體的構建及其對U87細胞增殖的影響
2017-11-01 05:56:56段景怡王舉波
山西醫科大學學報 2017年9期
段景怡,王舉波
(1寶雞市中醫醫院腦病二科,寶雞 721001;2西安交通大學第二附屬醫院神經外科;*通訊作者,E-mail:5195861@qq.com)
mIDH1真核表達載體的構建及其對U87細胞增殖的影響
段景怡1,王舉波2*
(1寶雞市中醫醫院腦病二科,寶雞 721001;2西安交通大學第二附屬醫院神經外科;*通訊作者,E-mail:5195861@qq.com)
目的 構建人mIDH1基因的真核表達載體及U87穩轉細胞系,觀察mIDH1對膠質瘤U87細胞體內外增殖的影響。 方法 以IDH1-cDNA為模板,采用PCR方法擴增IDH1片段,IDH1片段經雙酶切、連接重組至真核表達載體p-EGFP-C1,構建IDH1野生型(wIDH1)重組質粒,利用點突變技術通過IDH1真核表達載體構建突變型IDH1(mIDH1)真核表達載體,并通過雙酶切產物明膠電泳及DNA測序鑒定該重組質粒。Lip2000脂質體介導突變型IDH1轉染U87膠質瘤細胞,利用G418篩選構建穩定轉染p-EGFP-mIDH1真核表達載體的單克隆細胞株,構建穩轉細胞系,CCK-8法檢測U87細胞體外增殖能力,裸鼠成瘤實驗檢測mIDH1細胞系與對照U87細胞系的體內增殖。 結果 mIDH1重組質粒經雙酶切及測序鑒定證明mIDH1重組質粒構建成功,順利轉染U87膠質瘤細胞并高效表達,體外可抑制膠質瘤細胞的增殖,體內可減緩裸鼠皮下成瘤速度。 結論 p-EGFP-mIDH1真核表達載體可在U87膠質瘤細胞系中穩定表達,并可抑制膠質瘤細胞的增殖。
真核表達載體; 膠質瘤; U87細胞; IDH1基因; 穩轉細胞系; 細胞增殖
膠質瘤是神經系統最常見、最具有侵襲性的惡性腫瘤,約占所有腦腫瘤的45%[1]。腫瘤已經成為危害世界人民健康的公眾難題,而膠質瘤以其高發病率、髙致殘率、高死亡率、高復發率而躋身于人類有史以來最具危害性的腫瘤之一,盡管不斷有新的治療方法在嘗試[2-4],但其總體中位生存期不足15個月。長期以來,人們都在尋求治愈腫瘤的方法,在手術、放化療基礎上的各種綜合治療方法頻頻報道[5,6]。即使如此,膠質瘤治療的療效目前仍然不容樂觀。
2008年,Parsons等[7]在一項基因組研究計劃中發現異檸檬酸脫氫酶(isocitrate dehydrogenase,IDH)在膠質瘤患者中有著較高的突變率,其中以IDH1的突變率最為常見[8]。任曉輝等[9]的研究發現IDH1在膠質瘤中的突變率竟達到67%,而其中R132H的突變率竟然達到98.2%。臨床研究顯示突變型的IDH1(mutatant isocitrate dehydrogenase 1,mIDH1)患者相對于野生型的IDH1(wild isocitrate dehydrogenase 1,wIDH1)患者有著相對較好的預后,而腫瘤的增殖是影響患者生存的一大重要因素,突變型的IDH1膠質瘤患者相對良好的預后是否來源于其增殖速度的改變是本研究的出發點。因而深入研究IDH1及其突變體對膠質瘤細胞增殖影響的差異有著重要的價值。本研究以IDH1-cDNA為模板,利用PCR技術及DNA重組技術構建mIDH1真核表達載體,同時利用載體報告基因對其進行監測,構建穩定表達mIDH1蛋白的U87穩轉細胞系,研究mIDH1對膠質瘤細胞增殖與成瘤能力的影響,以期為深入研究mIDH1的功能及其在膠質瘤發生發展中的作用與機制提供基礎參考。
1 材料與方法
1.1 細胞株及主要試劑
U87細胞購自中國科學院上海細胞庫;p-EGFP-C1載體及DH5a菌株由西安交通大學中心實驗室饋贈。DMEM高糖培養基、胎牛血清等購自GIBIC公司;lipofectamine2000購自Invitrogen公司;DNA聚合酶、dNTP Mixture、限制性內切酶、T4 DNA ligase等購自TAKARA公司;去內毒素質粒小摘試劑盒、PCR產物純化試劑盒、膠回收試劑盒均購自美國OMEGA公司。
1.2 目的基因的獲取及擴增
用Primer Premier 5.0引物設計軟件設計特異性引物(Pcmv-IDH1 sequence:225-1466),并在目的基因的兩端加入限制性內切酶NcoⅠ(CCATGG)和EcoRⅤ(GATATC)的酶切位點(加粗部分)及相應的保護堿基(下劃線部分)。并在其兩端加入SacⅠ(GAGCTC)和BamHⅠ(GGATCC)的酶切位點和保護堿基,軟件設計引物如下:p1(翻譯引物):5′-ATCGAGCTCAGGAACTGGGGTGATAAGA-3′;p2(反向引物):5′-CGCGGATCCTTCACAAAGGTGGCAATAAC-3′;模板:Pcmv-IDH1 sequence 225-1466為IDH1 ORF。
以模板1.0 μl+p1 1.0 μl+p2 1.0 μl+5×Taq PCR Mix 4.0 μl+去離子水13.0 μl為反應體系,反應條件:94 ℃ 2 min,94 ℃變性30 s,51-61 ℃梯度退火30 s,72 ℃延伸30 s,共30循環,72 ℃延伸10 min,4 ℃ 10 min。
1.3 重組真核表達載體的構建
根據實驗前設計的BamHⅠ和SacⅠ酶切位點,利用限制性內切酶BamHⅠ和SacⅠ對IDH1的PCR擴增產物及p-EGFP-C1在31 ℃水浴條件下進行酶切4 h。酶切產物加入上樣緩沖液并通過瓊脂糖電泳分別純化回收。進而在T4 DNA ligase作用下于16 ℃連接過夜。次日,取5 μl連接產物轉化感受態細胞DH5a并將其涂布于含卡那霉素的LB選擇性瓊脂平板上進行篩選,挑取陽性單克隆,置于含卡那霉素的LB液體培養基中,37 ℃振搖培養14 h,去內毒素質粒抽提試劑盒抽提質粒,獲得野生型IDH1真核表達載體,即wIDH1。
根據突變試劑盒說明對目的基因進行定點突變改造,將反應產物進一步利用DNA明膠電泳法進行提純,細菌轉化,獲得單克隆細菌菌團,搖菌擴增,并對擴增提純質粒進行瓊脂糖電泳鑒定,同時將質粒送上海奧克公司測序確認。
1.4 U87細胞的培養及轉染
U87細胞接種于6孔板,細胞密度確保第2天每孔能匯合70%-80%,DMEM+10%FBS培養,CO2細胞培養箱(5%CO2,37 ℃)孵育細胞。將對數生長期的U87細胞按適當濃度接種于6孔板,常規培養24 h,取無抗生素、無血清培養基250 μl兩管,按1 ∶2.5的質粒、脂質體質量濃度比分別加入重組質粒與脂質體,室溫孵育5 min,將脂質體與質粒溶液混合,室溫孵育20 min,將上述混合液逐滴加入6孔板中(具體可參照Lipofectamine 2000說明書),饑餓培養4 h,吸棄去上述混合液,加入無抗生素的DMEM血清培養基繼續培養24 h。次日于熒光顯微鏡下觀察轉染效率。
1.5 mIDH1穩轉細胞系的構建
取轉染細胞用于篩選,以G418濃度500 μg/ml(預實驗確定的在10 d內使所有U87細胞死亡的最佳篩選濃度)的無抗生素DMEM血清培養液培養篩選細胞。每隔2-3 d常規更換培養液1次,經6周可見單克隆細胞團出現,熒光顯微鏡觀察可見單克隆細胞團發綠色熒光,局部標記后用消毒牙簽挑取單克隆細胞團,將其轉入96孔板繼續培養,并利用胰酶消化法逐步擴增傳代,連續傳代10代見細胞仍能全部發出綠色熒光,確認為穩轉細胞系構建成功。
1.6 膠質瘤細胞體外增殖能力檢測
取生長良好的對數生長期的mIDH1過表達U87膠質瘤細胞,以新配PBS漂洗2次,0.25%胰酶常規消化細胞,D-Hanks液終止消化,800 r/min離心5 min,棄上清,完全培養基培養重懸,將細胞濃度調整至1×105個/ml,按照2000細胞/孔分別接種于培養板內(96孔),每塊培養板設10個復孔,并做空白對照調零孔,共鋪設6塊培養板,常規培養,采用CCK-8法每天檢測1塊培養板,連續檢測6 d。檢測時于超凈臺內用微量移液器向每孔內加入CCK-8溶液10 μl,加入CCK-8溶液的培養板置入37 ℃細胞培養箱內繼續孵育4 h,終止培養,培養板輕輕振蕩1 min除去培養液中的氣泡,用酶聯免疫檢測分析儀測定A450值;以不加細胞并只加培養液的空白孔調零,于自動酶標讀數儀上測定各孔吸光度值(OD)值,計算各組的平均值;記錄結果。
1.7 膠質瘤細胞體內成瘤能力的檢測
4-5周齡雄性BALB/c裸小鼠12只,體質量20-21 g,由西安交通大學醫學院動物研究中心代購于上海中科院動物研究所,SPF級實驗室飼養。將12只裸鼠隨機分為2組,分別接種mIDH1過表達細胞及空載體對照細胞,每組6只。mIDH1過表達細胞及空載體對照細胞分別培養,常規消化、離心、收集細胞,細胞計數,重懸、離心收集細胞,適量無血清DMEM高糖培養基制成細胞懸液,4 ℃暫存,裸鼠皮膚接種處消毒,用注射針吸取細胞懸液,將注射針斜行刺入皮下,接種腫瘤細胞懸液,每只接種細胞1×107,mIDH1過表達細胞及空載體對照細胞按隨機分組各接種6只裸鼠。裸鼠接種腫瘤細胞后,每日觀察裸鼠的一般情況,包括精神狀態,飲食、活動情況、皮膚、體重等;每周測量腫瘤生長大小。測量時,以游標卡尺計算腫瘤大小,測量腫瘤的體表長徑與寬徑,計算瘤體積大小(V) ∶V=π(L2·W)/6。實驗第21天終止實驗,處死裸鼠,剝離腫瘤組織,記錄分析實驗數據,將腫瘤組織分塊標記。
2 結果
2.1 mIDH1重組真核表達載體構建成功
經過質粒酶切,與對應Marker比對提示,mIDH1質粒經限制性內切酶,切出約4 700,1 380 bp的片段(見圖1),分別代表質粒p-EGFP-C1和插入目的片段IDH1,表明IDH1真核表達載體構建成功。而構建載體經雙向測序也證實測序結果正確,目的基因序列與GENEBANK上公布的CDS編碼區完全相符。

圖1 mIDH1真核表達載體雙酶切鑒定Figure 1 Identification of p-EGFP-mIDH1 by double enzyme digestion
2.2 mIDH1真核表達載體在U87膠質瘤細胞中穩定表達
U87細胞轉染p-EGFP-mIDH1后轉染效率在60%-80%,可以用于進一步的穩轉細胞系篩選,經6周的篩選時間,最終篩選出穩定表達mIDH1的單克隆細胞(見圖2A)。最終經過10代以上的傳代后,所有細胞仍經熒光顯微鏡下(見圖2B)與復合光下(見圖2C)對比能穩定發出綠色熒光,提示轉染基因與膠質瘤細胞基因穩定整合,因mIDH1與GFP共用同一啟動子,故而mIDH1也得到穩定表達。
2.3 mIDH1真核表達載體抑制U87膠質瘤細胞的體外增殖與裸鼠皮下成瘤
細胞增殖體外實驗結果顯示,在接種后第1,2天,不同穩轉細胞的增殖速度并無明顯異常,而在第3,4,5天mIDH1過表達細胞增殖速度明顯落后于對照細胞,吸光度值差距明顯(P<0.01,見表1);而隨著時間逐漸延長,這種差異不顯著,考慮與細胞之間受到接觸抑制的影響,細胞增殖速度變慢,因而細胞之間的增殖速度差異逐漸縮小。而如果進一步將接種細胞數減少,延長細胞培養時間,在細胞鋪滿培養孔底壁之前,細胞增殖速率之間的差異持續存在。
而在體內裸鼠成瘤實驗中,單純裸鼠成瘤結果顯示:mIDH1及空載體對照細胞均可用于裸鼠成瘤,而且成瘤率高,受試裸鼠全部成瘤。經過3周的觀察,小鼠飲食、行為無明顯改變。實驗結束后,根據V=π(L·W2)/6公式計算每只裸鼠腫瘤的體積。使用SPSS16.0統計軟件進行數據分析,細胞成瘤體積大小以均數±標準差表示,mIDH1組腫瘤體積較小,為135.43±15.62;與對照組(193.57±18.53)比較,差異有統計學意義(P<0.05)。這一結果與體外試驗中檢測到的腫瘤細胞間的增殖差異基本相符。

A.單細胞克隆 B.熒光下的穩轉細胞 C.復合光下的穩轉細胞圖2 穩轉細胞系中mIDH1與熒光共表達圖(×200)Figure 2 Co-expression of mIDH1 and fluorescence in stable cell lines (×200)
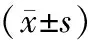


組別1d2d3d4d5d6d mIDH1組0113±00080179±00090330±0015?0779±0039?1131±0066?1743±0109 對照組0136±00050245±00140608±00171334±01161551±00881805±0128
與對照組比較,*P<0.05
3 討論
IDH是三羧酸循環中的重要限速酶,對細胞體的物質代謝與能量傳遞起著至關重要的作用。近期研究發現IDH1基因突變改變了傳統的三羧酸循環途徑,導致a-酮戊二酸生成2-羥基戊二酮[10]。因而有人認為,酮代謝異常導致了膠質瘤的發生,然而卻未能找到充分的證據,即使是在先天性苯丙酮尿癥患者中也未見有腫瘤高發率的報道。而最近也有研究顯示膠質瘤的發生與否以及腫瘤體積的大小與a-酮戊二酸并無直接關系。相反IDH1突變可能對膠質瘤患者是一種保護因素,正常的IDH1功能細胞可以提供細胞代謝所需能量,產生大量的NADPH對抗體內產生的氧自由基。而突變型的IDH1不僅不能產生NADPH,反而進一步消耗NADPH至更低的水平[10]。這在一定程度上降低了細胞對抗氧化代謝的能力及自身修復的能力,使得細胞更容易受到氧化損傷,這或許能解釋為什么突變型的IDH1膠質瘤患者有著相對較好的預后,究其原因可能是突變型的IDH1膠質瘤患者對放化療有著更好的敏感性。
腫瘤細胞的增殖速度往往與患者預后有較大的關系。實驗研究顯示,mIDH1過表達細胞在第2-5天增殖速度明顯快于對照組過表達細胞,差異有統計學意義,但從第6天開始,增殖速度差異縮小,可能與快速增殖的細胞增殖空間受限或接觸抑制有關。上述研究表明mIDH1膠質瘤細胞增殖速度較野生型慢,這也與臨床觀察到的mIDH1患者具有相對較好的預后相吻合。
細胞增殖是與細胞周期密切相關的一個生理過程。人體正常的組織細胞內,除了大部分的非分裂細胞構成的組織外,多數成熟的細胞都含有保持分裂能力的靜止期細胞,當受傷或受到外界刺激時可再次進入細胞周期。根據再生能力強弱可將人體組織細胞分為不穩定細胞、穩定細胞和永久性細胞;而永久性細胞是指不能分裂增生的組織細胞,如神經節細胞、心肌細胞等。相比之下,永生性細胞是指細胞的增殖周期調控出現差錯,凋亡增殖比例嚴重失調,細胞持續處于增殖時,表現出無限增殖的特性,這就是我們常說的腫瘤。但是,即使是在腫瘤細胞內,無論是否符合程序,它仍然存在一定的增殖與周期調控機制,不同的腫瘤細胞之間其增殖差異往往較大。膠質瘤中IDH1功能酶的突變可能導致了細胞代謝產物的改變[10],其代謝產物可能對細胞增殖起到了抑制作用,使得細胞不能在G1前期合成細胞分裂所需要的特殊RNA或代謝調節酶,或者使得該過程減速,大量細胞阻滯于G1期,致使細胞周期不能啟動。而另一個更為可能的原因是,突變的IDH1影響三羧酸循環正常進行,導致NADPH不足與磷酸化供能不足,從而影響組蛋白甲基化,影響細胞的分化增殖[11]導致細胞增殖周期的差異,最終導致了細胞增殖的差異,而其細胞周期的差異已在前面的周期檢測中得到證實。這與Seltzer等[12]的研究有著一致性。
此外,在G1期內存在一個校正點或阻止點(簡稱R點),主要控制G1期時間的長短。通過了此點,細胞就能以正常速度不受外界條件的影響而完成細胞周期的其他時期,因而也有人認為細胞周期的終點在于R點,即為止于R點的細胞將退出細胞周期,進入G0期。IDH1的基因突變也可能導致細胞代謝微環境改變,使得細胞不能正常跨過R點,因而導致G1期阻滯。S期即DNA合成期,在此期,主要合成DNA和組蛋白。DNA復制所需要的酶都在這一時期合成,因而,mIDH1可能通過改變G1/G2期變化影響細胞周期,又因mIDH1為非致死性基因變化,考慮可能通過G1期的細胞都能進行DNA合成,從而延續細胞的周期分裂,維持細胞生存,而DNA及組蛋白合成緩慢,缺乏物質合成所必需的能量等導致S期向G2期過渡轉化緩慢所致。G2期是有絲分裂的準備期,在這一時期,DNA合成終止,大量合成RNA及蛋白質,包括微管蛋白和促成熟因子等。而正常生理狀態條件下,靜止細胞處于G0期。不同種類的細胞其增殖的周期長短不同,在相同時間內進入增殖周期的細胞數也不同,因而具有不同的增生能力。
深入研究mIDH1對膠質瘤細胞細胞生物學行為的影響,可以為臨床病理亞型的確立提供更多的理論依據,為尋找膠質瘤治療的新方法提供更多的理論基礎,野生型的IDH1或突變型IDH1有望作為膠質瘤治療的一個新型靶點進一步深入研究[13],或許在不久的將來,更多的膠質瘤患者可以從中獲益。
[1] 王忠誠.王忠誠神經外科學[M].武漢:湖北科技出版社,2005:512-519.
[2] Gao C, Liang C, Nie Z,etal. Alkannin inhibits growth and invasion of glioma cells C6 through IQGAP/mTOR signal pathway[J]. Int J Clin Exp Med, 2015, 8(4):5287-5294.
[3] Li XT, Ju RJ, Li XY,etal. Multifunctional targeting daunorubicin plus quinacrine liposomes, modified by wheat germ agglutinin and tamoxifen, for treating brain glioma and glioma stem cells[J]. Oncotarget, 2014, 5(15):6497-6511.
[4] Hottinger AF, Pacheco P, Stupp R. Tumor treating fields: a novel treatment modality and its use in brain tumors[J]. Neuro Oncol, 2016, 18(10):1338-1349.
[5] Li J, Cai J, Zhao S,etal. GANT61, a GLI inhibitor, sensitizes glioma cells to the temozolomide treatment[J]. J Exp Clin Can-cer Res, 2016, 35(1):184-197.
[6] Yang QY, Guo CC, Chen ZP. Profile of nimotuzumab in the treatment of high-grade glioma[J]. Onco Targets Ther,2015,8:819-825.
[7] Parsons DW, Jones S, Zhang X,etal. An integrated genomic analysis of human glioblastoma multiforme[J]. Science, 2008, 321(5897):1807-1821.
[8] Bleeker FE, Lamba S, Leenstra S,etal. IDHl mutations at residue p.R132 (IDHI(R132)) occur frequently in high-grade gliomas but not in other solid tumors[J]. Hum Mutat, 2009, 30(1):7-11.
[9] 任曉輝,崔向麗,尚志梅,等.少枝膠質腫瘤異檸檬酸脫氫酶1基因突變特點分析[J].中華醫學雜志,2012,92(11):724-726.
[10] Dang L, White DW, Gross S,etal. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate[J].Nature,2009,462(7274):739-744.
[11] Lu C, Ward PS, Kapoor GS,etal. IDH mutation impairs histone demethylation and results in a block to cell differentiation[J]. Nature, 2012, 483(7390):474-478.
[12] Seltzer MJ, Bennett BD, Joshi AD,etal. Inhibition of glutaminase preferentially slows growth of glioma cells with mutant IDH1[J]. Cancer Res, 2010, 70(22):8981-8987.
[13] Zhang C, Moore LM, Li X,etal. IDH1/2 mutations target a key hallmak of cancer by deregulating cellular metabolism in glioma[J]. Neuro Oncol, 2012, 15(9): 1114-1126.
ConstructionofmIDH1eukaryoticexpressionvectoranditseffectonproliferationofU87cells
DUAN Jingyi1,WANG Jubo2*
(1DepartmentofNeurology,BaojiHospitalofTraditionalChineseMedicine,Baoji721001,China;2DepartmentofNeurosurgery,SecondAffiliatedHospitalofXi’anJiaotongUniversity;*Correspondingauthor,E-mail:5195861@qq.com)
ObjectiveTo construct the eukaryotic expression vector of human mIDH1 gene and its stable expression cell line of U87 for investigating its influence on cell proliferationinvivoandinvitro.MethodsWith cDNA fragment of IDH1 as template, IDH1 fragment was amplified by PCR. Then, IDH1 gene was cloned into the p-EGFP-Cl vector after double digestion and ligation to get wild IDH1(wIDH1) vector.The wIDH1 vector was used to construct the mutant IDH2(mIDH1) vector using point mutation technology, and the mIDH1 vector was identified by enzyme digestion and DNA sequence analysis. The recombinant plasmids were transfected into U87 cells by lipofectamine 2000. The cell clone that stably expressing mIDH1 was selected out by G418 method and further to get the stable cell line. CCK-8 method was used to analyze the proliferation ability of U87 cell lineinvitro, and the tumor formation experiment was used to detect the proliferation of mIDH1 cell line and control U87 cell lineinvivo.ResultsAgarose electrophoresis analysis of plasmid resection and sequencing proved that the recombinant plasmid mIDH1 was constructed successfully. The mIDH1 was transfected successfully and stably expressed in U87 cell line. The mIDH1 inhibited the proliferation of U87 cell lineinvitroand slowed down the tumor formation speedinvivo.ConclusionThe eukaryotic expression plasmid mIDH1 can be stably expressed in U87 cell line and inhibit the proliferation of U87 cell lineinvivoandinvitro.
eukaryotic expression vector; glioma; U87 cells; IDH1 gene; stable expression cell line; proliferation
R739.4
A
1007-6611(2017)09-0913-05
10.13753/j.issn.1007-6611.2017.09.010
段景怡,女,1982-12生,碩士,主治醫師,E-mail:121436425@qq.com
2017-04-27
