擬南芥表皮細胞發育關鍵調控因子GL2的原核表達及多克隆抗體制備
2017-10-27 07:28:55劉靜文張翔齊亞飛
江蘇農業科學 2017年16期
劉靜文 張翔 齊亞飛


摘要:擬南芥HD-ZIP家族轉錄因子GLABRA2(GL2)是控制植物表皮毛、根毛分化和發育的關鍵調控因子。為了深入研究GL2調控植物表皮細胞分化發育的分子機制,對GL2蛋白進行了生物信息學分析,選擇其保守性較低的C端區域(597~776 aa)作為抗原區,構建了pET28a/GL2原核表達系統。通過異丙基-β-D-硫代半乳糖苷誘導獲得了融合蛋白rGL2597~776,分子量約為196 ku。利用純化得到的融合蛋白免疫家兔,獲得了抗GL2的多克隆抗體,并利用抗原親和純化了GL2多克隆抗體。GL2多克隆抗體的制備及純化為進一步通過染色體免疫共沉淀等方法篩選和鑒定其下游作用靶基因奠定了基礎。
關鍵詞:表皮毛;根毛;GL2轉錄因子;抗體制備;原核表達
中圖分類號: Q786文獻標志碼:
文章編號:1002-1302(2017)16-0029-04
收稿日期:2016-04-08
基金項目:國家自然科學基金(編號:31470290);中央高校基本科研業務費(編號:2014YB036、Z109021537)。
作者簡介:劉靜文(1990—),女,新疆塔城人,碩士研究生,主要從事擬南芥表皮毛分化發育調控機理研究。E-mail:ljw0993@163com。
通訊作者:安麗君,博士,副教授,主要從事植物分子遺傳與發育生物學研究。E-mail:lijunan@nwsuafeducn。
植物表皮毛和根毛是由表皮細胞向外延伸形成的特殊結構,它們不僅是研究植物細胞分化發育的優良模式系統,而且在幫助植物抵御生物和非生物逆境脅迫中發揮著重要作用,因而具有重要的科研價值和實用意義[1-2]。植物表皮毛和根毛的發育受機體本身遺傳因素以及激素水平、水分、光照等環境因素的共同影響[3]。GL2是最早被發現的控制擬南芥表皮毛和根毛分化發育的關鍵調控因子之一,是表皮毛細胞正常發育所必需的調控因子之一[4-5]。GL2[編碼1個含有 HD-ZIP 結構域的轉錄因子,其T-DNA插入功能缺失突變體 gl2-3[WTBZ][STBZ]表現為表皮毛嚴重發育受阻,葉片表面光滑無毛,只在葉片的邊緣有極少的單根毛和毛狀突起,而根毛數量增多,根毛長度增加[4]。在葉片發育的早期,[GL2基因在整個葉片中表達,而隨著葉片的成熟,只在表皮毛細胞及其支持細胞中表達;在根中,在非根毛細胞中特異性表達,參與根毛和非根毛細胞命運的決定[6-7]。
目前,GL2[WTB調控植物表皮細胞分化發育的上游途徑已經比較明確,GL1-GL3/EGL3-TTG1轉錄復合體能夠激活基因的表達,并且以劑量依賴的方式調控細胞發育過程[8-9]。但是對于下游的調控路徑尚不清楚,GL2作為轉錄因子可能通過調控下游靶基因的表達來影響細胞的分化發育。GL2特異性的多克隆抗體能夠廣泛應用于檢測GL2蛋白表達量和染色體免疫共沉淀等試驗中,在基因組范圍內篩選受GL2調節的基因及GL2下游直接作用靶基因,為深入探討GL2參與的表皮細胞分化發育調控網絡奠定基礎。
1材料與方法
11試驗材料
111供試材料
擬南芥ColumbiaCol-0生態型野生型種子,由筆者所在實驗室保存;試驗用家兔,由西北農林科技大學動物飼養場提供,選取3只健康成年家兔用于試驗。
112載體、菌株和試劑
原核表達載體pET28a及大腸桿菌Top10和BL21()感受態細胞,由筆者所以實驗室保存;試驗中所用限制性內切酶、Taq DNA聚合酶、T4連接酶、牛小腸堿性磷酸酶、反轉錄酶等,均購自寶生物工程(大連)有限公司;弗氏完全佐劑與不完全佐劑、蛋白免疫印跡化學發光試劑盒及HRP標記的羊抗兔抗體IgG,購自Bio-Rad公司;His標簽蛋白純化柱填料Ni Sepharose 6 Fast Flow、硝酸纖維素膜(045 μm),均購自GE Healthcare Life Science;6×His抗體,購自Abcam公司。
12試驗方法
121表達序列的選取及載體構建
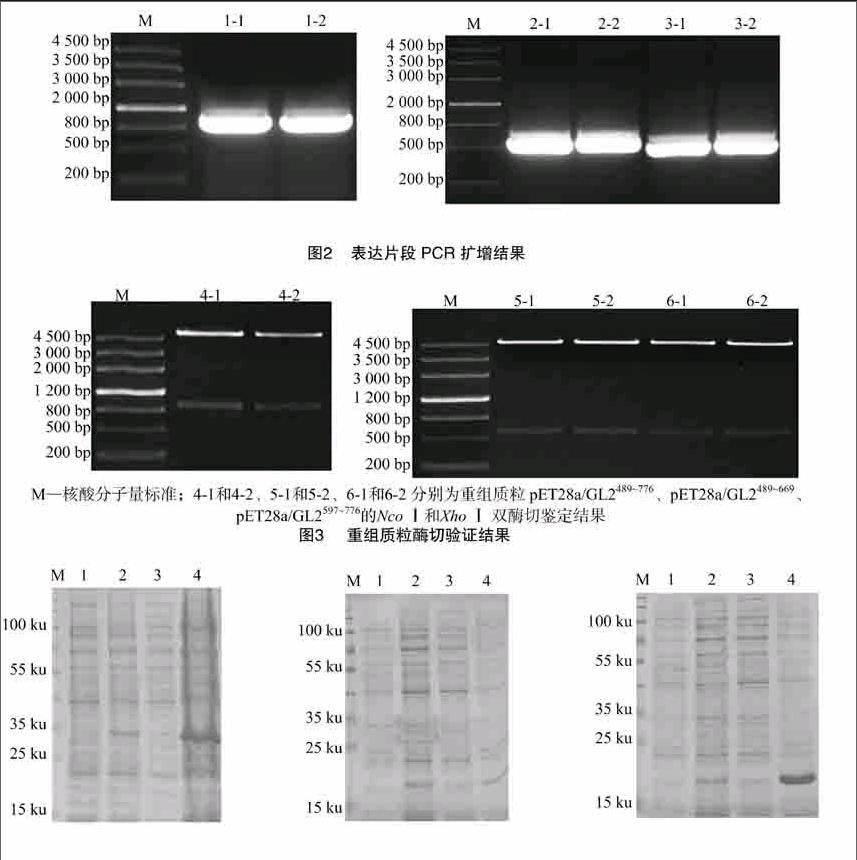
對基因及其推測的氨基酸序列進行分析,選取基因3′端長度為 865 bp 的片段進行PCR擴增,作為抗體制備的目的片段。取2周齡擬南芥野生型全部地上組織,提取總RNA,反轉錄獲得cDNA,以cDNA為模板分別用帶有NcoⅠ和XhoⅠ酶切位點的3對引物(GL2F1/R1、GL2F2/R2、GL2F3/R3)對選取的序列進行PCR擴增,得到的擴增片段與pET28a載體進行連接并轉化大腸桿菌Top10[JP3]感受態細胞,將鑒定獲得的陽性重組質粒進行測序。PCR擴增所用引物序列如表1所示。[JP]
122重組蛋白的表達與純化
將測序正確的重組質粒轉化大腸桿菌BL21(菌株,獲得單克隆后將其接種于含有卡那霉素的LB培養基中,37 ℃、220 r/min培養至對數生長期,加入誘導劑異丙基-β-D-硫代半乳糖苷(IPTG)至終濃度為10 mmol/mL,37 ℃誘導表達4 h;然后離心收集菌體并
進行超聲破碎,12 000 r/min離心30 min,利用十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(SDS-PAGE)檢測上清液及沉淀中蛋白的表達情況和存在形式,以誘導前的菌液為對照組。選擇表達量較高的重組蛋白進行免疫印跡試驗,對比免疫信號最終確定出最優重組蛋白作為免疫抗原進行進一步純化[10-11]。對確定的最優重組蛋白進行大量誘導表達,用Ni Sepharose 6 Fast Flow親和柱純化融合蛋白,最后以梯度濃度的牛血清白蛋白為對照,進行SDS-PAGE電泳,通過Image Lab軟件灰度分析對目的蛋白濃度進行定量[12]。
123多克隆抗體的制備endprint
以純化獲得的rGL2597~776重組蛋白免疫家兔,抗原量為200 μg/只,初次免疫將抗原與弗氏完全佐劑按體積比1 ∶[KG-3]1混合,背部脊柱兩側進行皮下多點注射。第一次免疫后每隔2周加強免疫1次,共免疫4次。加強免疫用弗氏不完全佐劑,抗原量為100 μg/只。最后一次免疫 7 d 后抽取家兔心頭血約40 mL,將血液于37 ℃放置1 h后取出再置于4 ℃過夜。收集析出的血清,1 000 r/min離心 10 min 收集上清液即為免疫獲得的抗血清[13-14]。
124多克隆抗體的特異性檢測
將作為抗原的重組蛋白按1 ∶[KG-3]10 000、1 ∶[KG-3]1 000、1 ∶[KG-3]100稀釋后進行SDS-PAGE電泳,將免疫獲得的血清按1 ∶[KG-3]1 000的比例作為一抗加入進行免疫印跡雜交,通過化學發光顯色系統檢測目標位置的雜交信號,根據信號強弱計算抗體效價[15]。
125GL2多克隆抗體的純化
為了獲得純化的Anti-GL2抗體,降低免疫印跡試驗中背景噪點的影響,利用抗原-抗體的特異性對得到的抗血清進行純化。取約1 mg的抗原進行SDS-PAGE并轉膜使其結合于硝酸纖維素膜上;將抗原結合區域的膜經100 mmol/mL酸性甘氨酸/鹽酸溶液(pH值為25)洗脫后封閉處理1 h;將處理后的膜裝入含有一抗(GL2抗血清)的離心管中,室溫孵育2~3 h后轉入4 ℃振蕩孵育過夜;洗膜后加入甘氨酸溶液,室溫孵育10 min,收集孵育體系中的甘氨酸溶液用1 mol/L Tris溶[JP3]液(pH值為80)調節pH值為70,獲得的甘氨酸溶液即為純化的 Anti-GL2抗體。[JP]
2結果與分析
21序列分析及表達序列的選取
利用擬南芥基因組數據庫(wwwarabidopsisorg)獲得了基因全長序列及推測的蛋白質序列。由圖1-a可知,基因cDNA全長為2 681 bp,開放閱讀框為2 331 bp,推測編碼1個含有776個氨基酸殘基的蛋白質(圖1-b)。為了使GL2抗體能有效地檢測到抗原,分別在SMART(http://smartembl-heidelbergde)和SWISS-MODEL(http://wwwexpasych/swissmod/SWISS-MODELhtml)上對GL2的結構域和三級結構進行了預測,由圖1-b可知,GL2蛋白包含有1個保守的同源區和1個起始翻譯區,分別覆蓋130~192、280~510 aa區域;在GL2蛋白的N端(1~129 aa)和C端(516~776 aa)保守性較低。由圖1-c可知,GL2蛋白的C端親水程度較高,經過折疊后暴露在外面,而其N端包埋在整個蛋白內部的疏水區域中,所以選取GL2蛋白C端489~776 aa作為原核表達和制備抗體目的蛋白序列。
由表2可知,該蛋白序列兩端的氨基酸組成存在明顯的差異,489~669 aa區域的氨基酸殘基主要由帶正電的氨基酸構成,而597~776 aa區域主要由帶負電的氨基酸殘基構成,所以導致該目的蛋白序列兩端的電性相反,N端等電點為1073,C端等電點為490。考慮到這些因素可能影響到后續的表達試驗,又進一步將此段蛋白序列分為2個片段連同之前選擇的C端序列一起進行表達,并將這些片段分別命名為GL2489~776、GL2489~669、GL2597~776。
22表達序列的PCR擴增及重組質粒pET28a/GL2的構建
提取2周齡[WTBX][STBX]Col-0[WTBZ][STBZ]背景野生型擬南芥全部地上組織的總RNA并反轉錄獲得cDNA,以cDNA為模板,分別以GL2489~776、GL2489~669、GL2597~776的特異引物GL2F1/R1、GL2F2/R2、GL2F3/R3進行擴增,結果(圖2)表明,分別得到長度為864、543、537 bp的表達片段。將PCR擴增獲得的DNA片段與原核表達載體pET28a同時進行NcoⅠ和XhoⅠ雙酶切,并進行連接,分別構建pET28a/GL2489~776、pET28a/GL2489~669 、pET28a/GL2597~776重組質粒,轉化大腸桿菌TOP10感受態細胞并提取質粒進行雙酶切,結果(圖3)表明,3段表達序列均成功連入pET28a表達載體。[FL)]
23GL2重組蛋白的原核表達、純化及定量分析
將構建好的重組質粒pET28a/GL2489~776、pET28a/GL2489~669、pET28a/GL2597~776分別轉化大腸桿菌BL21感受態細胞,建立重組蛋白的原核表達系統。37 ℃條件下,對重組蛋白進行IPTG誘導使得目的蛋白大量表達。超聲破碎菌體后分別取上清和沉淀進行SDS-PAGE檢測目的蛋白誘導表達情況及其存在形式,以誘導前和誘導后的菌液為對照。由圖4可知,在3個表達體系中,誘導后菌液中均出現了目標蛋白條帶,而且其大小與重組蛋白的分子量對應,證明成功誘導得到重組蛋白。對比上清與沉淀的蛋白條帶可以判定3種重組蛋白主要以包涵體的形式存在
從SDS-PAGE結果可以看出,重組質粒pET28a/GL2489~669、 pET28a/GL2597~776的表達量較高,所以選用這2個重組質粒進行進一步的篩選。利用誘導表達獲得的重組蛋白和6×His抗體進行免疫印跡試驗,由圖5可知,超聲破碎后,rGL2597~776沉淀中的重組蛋白表達量高于rGL2489~669,所以最終選擇以rGL2597~776重組蛋白為抗原進行GL2抗體的制備。
重組蛋白rGL2597~776作為免疫抗原需要進一步進行純化。利用Ni Sepharose 6 Fast Flow親和柱對rGL2597~776進行親和層析純化,最終得到純度較高的重組蛋白rGL2597~776。以牛血清蛋白濃度梯度為標準對獲得的重組蛋白進行定量,通過軟件灰度分析計算,最終得到40 mL濃度為09 mg/mL的重組蛋白,重組蛋白總量為36 mg。endprint
24多克隆抗體的制備與免疫檢測
以純化后的重組蛋白rGL2597~776作為抗原對家兔進行4次免疫獲得含有GL2多克隆抗體的抗血清,將抗原稀釋 10 000、1 000、100倍后與稀釋1 000倍的抗血清進行免疫印跡試驗。由圖6可知,獲得的抗血清可以特異地與誘導表達后的抗原結合, 這說明成功得到重組蛋白特異性的多克隆抗
體。利用Image Lab軟件進行灰度分析計算,抗血清最低能夠檢測到的抗原量為86 ng。
25多克隆抗體的純化
所獲得的抗血清除了含有GL2抗體外,可能還包含其他的免疫球蛋白,為了提高GL2多克隆抗體的特異性,依據抗原-抗體特異性結合的原理對獲得的抗血清進行純化,以減少免疫檢測試驗中背景噪點的影響。首先將抗血清在硝酸纖維素膜上與含大量抗原的溶液進行免疫結合,然后通過調節溶液成分和pH值將特異性結合的抗體洗脫下來,成功獲得純度較高的多克隆抗體(圖7
3討論
擬南芥GL2蛋白相對較大,且含有2個較大的保守結構域同源區和起始翻譯區,所以選取部分蛋白序列作為免疫抗原以保證所制備抗體的特異性。在對GL2重組蛋白序列的選擇上充分考慮蛋白結構特征與不同結構域的保守性情況,在保證表達蛋白特異性的前提下選取多個表達區段同時進行載體構建和原核表達,避免氨基酸組成差異等因素對重組蛋白表達的影響。本試驗通過對比選擇了表達量高且免疫性好的重組蛋白rGL2597~776作為抗原。pET28a是常用的原核表達載體,它含有的T7轉錄系統能夠強啟動融合基因的表達,并且其多克隆位點兩端的6×His標簽可以選擇與表達蛋白的C端和N端進行融合。融合了的6×His標簽一方面可以幫助重組蛋白結合鎳柱純化,另一方面借助Anti-His免疫印跡試驗可以有效地驗證所表達的重組蛋白的正確性。通過對抗血清中GL2多克隆抗體的親和純化,能夠有效去除其他非特異性免疫球蛋白,使得獲得的多克隆抗體在植物組織蛋白等其他成分復雜的蛋白溶液中的檢測更為特異。
利用GL2多克隆抗體可以進行染色質免疫共沉淀以及ChIP-seq等試驗,在基因組水平上篩選GL2潛在的下游直接作用靶基因和互作因子,進而探索控制植物表皮毛和根毛發育的基因調控網絡,所以,GL2多克隆抗體的制備可以為植物表皮毛和根毛發育調控體系的研究奠定基礎。
參考文獻:
[ZK(#]Yang C,Ye Z Trichomes as models for studying plant cell differentiation[J] Cellular and Molecular Life Sciences,2013,70(11):1937-1948
Schellmann S,Hülskamp M Epidermal differentiation:trichomes in Arabidopsis as a model system[J] International Journal of Developmental Biology,2005,49(5/6):579-584
[3]Balkunde R,Pesch M,Hülskamp M Trichome patterning in Arabidopsis thaliana from genetic to molecular models[J] Current Topics in Developmental Biology,2010,91(10):299-321
[4]An L,Zhou Z,Yan A,et al Progress on trichome development regulated by phytohormone signaling[J] Plant Signaling & Behavior,2011,6(12):1959-1962
[5]Guan X Y,Li Q J,Shan C M,et al The HD-Zip IV gene [WTBX][STBX]GaHOX1[WTBZ][STBZ] from cotton is a functional homologue of the Arabidopsis [WTBX][STBX]GLABRA2[WTBZ][STBZ][J] Physiologia Plantarum,2008,134(1):174-182
Tominaga-Wada R,Ishida T,Wada T New insights into the mechanism of development of Arabidopsis root hairs and trichomes[J] International Review of Cell and Molecular Biology,2011,286:67-106
[7]Ishida T,Kurata T,Okada K,et al A genetic regulatory network in the Arabidopsis trichome and root hair initiation[J] Annual Review of Plant Biology,2008,59:365-386
[8]Grebe M The patterning of epidermal hairs in Arabidopsis-updated[J] Current Opinion in Plant Biology,2012,15(1):31-37
[9]Morohashi K,Grotewold E A systems approach reveals regulatory circuitry for Arabidopsis trichome initiation by the GL3 and GL1 selectors[J] PLoS Genetics,2009,5(2):e1000396
[10][ZK(#]Yu F,Park S S,Liu X,et al Suppessor of [WTBX][STBX]variegation4[WTBZ][STBZ],a new [WTBX][STBX]var2[WTBZ][STBZ] suppressor locus,encodes a pioneer protein that is required for chloroplast biogenesis[J] Molecular Plant,2011,4(2):229-240
[11]王增,馬會勤,張文,等 包涵體蛋白的分離和色譜法體外復性純化研究進展[J] 中國生物工程雜志,2009,29(7):102-107
[12]單麗偉,唐如春,劉三陽,等 小麥種子過氧化物酶[WTBX][STBX]WP1[WTBZ][STBZ]基因的原核表達、純化及多克隆抗體制備[J] 生物工程學報,2011,27(1):26-30
[13]李芳,羅軍,許會芬,等 西農薩能羊MAT基因的原核表達及多克隆抗體制備[J] 西北農林科技大學學報(自然科學版),2014,42(12):1-6,12
[14]許君,喬帥,常嵐,等 苦瓜核糖體失活蛋白MAP30 的原核表達與抗體制備[J] 河南農業大學學報,2009,43(5):552-556
[15]張鈺,李亞會,屠唯一,等 擬南芥AtERF1蛋白的原核可溶性表達及多克隆抗體的制備[J] 上海師范大學學報(自然科學版),2013,42(4):378-383endprint
