晚期氧化蛋白產(chǎn)物調(diào)節(jié)人腎小管上皮細(xì)胞自噬的研究
2017-10-13 03:31:57向曉紅梁秀潔束雙雙姜婷婷郭婷婷
重慶醫(yī)學(xué) 2017年25期
章 俊,向曉紅,梁秀潔,束雙雙,姜婷婷,郭婷婷,湯 珣
(南方醫(yī)科大學(xué)珠江醫(yī)院腎內(nèi)科,廣州 510280)
論著·基礎(chǔ)研究doi:10.3969/j.issn.1671-8348.2017.25.008
晚期氧化蛋白產(chǎn)物調(diào)節(jié)人腎小管上皮細(xì)胞自噬的研究
章 俊,向曉紅,梁秀潔,束雙雙,姜婷婷,郭婷婷,湯 珣△
(南方醫(yī)科大學(xué)珠江醫(yī)院腎內(nèi)科,廣州 510280)
目的探討晚期氧化蛋白產(chǎn)物(AOPPs) 對(duì)人腎小管上皮細(xì)胞(HK-2)自噬的影響。方法AOPPs刺激HK-2細(xì)胞,采用RT-qPCR和Western blot檢測(cè)自噬相關(guān)蛋白LC3-Ⅱ/LC3-Ⅰ、Beclin1和p62的表達(dá);Western blot檢測(cè)p38 MAPK通路的激活;加入p38 MAPK抑制劑(SB203580)與AOPPs共處理,觀察自噬的改變,加入自噬誘導(dǎo)劑雷帕霉素與AOPPs共處理,并用RT-qPCR和Western blot檢測(cè)細(xì)胞周期抑制蛋白p27的表達(dá),用BCA法檢測(cè)細(xì)胞總蛋白,觀察細(xì)胞肥大的改變。結(jié)果AOPPs下調(diào)LC3-Ⅱ/LC3-Ⅰ、Beclin1,上調(diào)p62的表達(dá),并激活p38 MAPK 通路;與AOPPs單獨(dú)處理組相比,SB203580與AOPPs共處理組LC3-Ⅱ/LC3-Ⅰ和Beclin升高而p62降低;雷帕霉素與AOPPs共處理組p27的表達(dá)和細(xì)胞總蛋白下調(diào)。結(jié)論AOPPs通過激活p38 MAPK通路抑制HK-2細(xì)胞自噬,自噬抑制參與HK-2細(xì)胞肥大。
自噬; p38 MAPK信號(hào)通路; 晚期氧化蛋白產(chǎn)物
晚期氧化蛋白產(chǎn)物(AOPPs)是血清蛋白被自由基和反應(yīng)性氧系統(tǒng)氧化后的產(chǎn)物,在糖尿病腎病(DN)患者血漿中明顯增加,參與DN腎臟實(shí)質(zhì)和間質(zhì)的損害[1]。本課題組前期研究證明AOPPs導(dǎo)致人腎小管上皮細(xì)胞(HK-2)肥大[2],然而其機(jī)制尚未完全明確。自噬是普遍存在于真核細(xì)胞中的一種依賴溶酶體的胞內(nèi)降解系統(tǒng),能清除細(xì)胞中受損或衰老的細(xì)胞器和大分子物質(zhì),維持細(xì)胞內(nèi)環(huán)境穩(wěn)定。自噬受到多種信號(hào)通路的精密調(diào)控,p38 MAPK屬于其中的非哺乳動(dòng)物雷帕霉素靶蛋白(mTOR)依賴途徑[3]。前期研究表明,AOPPs可誘導(dǎo)HK-2細(xì)胞中p38 MAPK信號(hào)通路活化[4]。然而,AOPPs對(duì)HK-2細(xì)胞自噬有無影響,是否與p38 MAPK信號(hào)通路有關(guān)尚不明確。本實(shí)驗(yàn)通過研究AOPPs對(duì)HK-2細(xì)胞自噬的影響及其可能機(jī)制,以明確自噬在AOPPs導(dǎo)致HK-2細(xì)胞肥大中的作用。
1 材料與方法
1.1材料 HK-2細(xì)胞株為條件永生性細(xì)胞系,購自美國ATCC細(xì)胞庫;胎牛血清(FBS,美國Gibco);DMEM/F12細(xì)胞培養(yǎng)基(美國Hyclone);無毒素牛血清清蛋白(BSA,美國Sigma);Trizol試劑、cDNA第一鏈合成試劑盒和SYBR Premix Ex TaqTM熒光定量PCR試劑盒(日本Takara);p27、p62、Beclin1及內(nèi)參GAPDH引物由生物工程上海(股份)有限公司合成;p27、LC3B、Beclin1、p62、phospho-p38 MAPK (Thr180/Tyr182)、p38 MAPK兔抗人一抗及p38 MAPK信號(hào)通路特異性抑制劑SB203580,自噬誘導(dǎo)劑雷帕霉素 (美國CST);GAPDH兔抗人一抗(美國Bioworld);HRP標(biāo)記的羊抗兔二抗(北京Bioss);ECL檢測(cè)試劑盒(美國Millipore)。
1.2方法
1.2.1體外制備AOPPs AOPPs的制備參見文獻(xiàn)[4]。
1.2.2HK-2細(xì)胞的培養(yǎng) 將HK-2細(xì)胞培養(yǎng)于含10%FBS和青霉素鏈霉素雙抗的DMEM/F12培養(yǎng)基中,胰酶消化后接種,加藥前換用無血清培養(yǎng)基同步過夜。加SB203580和雷帕霉素組先分別用10 μmol/L SB203580和100 nmol/L雷帕霉素預(yù)處理細(xì)胞1 h,再與AOPPs共同處理細(xì)胞24 h。
1.2.3實(shí)驗(yàn)分組 為明確AOPPs對(duì)HK-2細(xì)胞自噬和p38 MAPK信號(hào)通路的影響,將細(xì)胞分為3組:(1)Ctrl組;(2)BSA組:200 μg/mL BSA;(3)AOPPs組:200 μg/mL AOPPs作用24 h。為明確p38 MAPK信號(hào)通路是否參與AOPPs對(duì)HK-2細(xì)胞自噬的調(diào)節(jié),將細(xì)胞分為3組:(1)Ctrl組;(2)AOPPs組:200 μg/mL AOPPs;(3)AOPPs+SB203580組:10 μmol/L SB203580預(yù)處理1 h后,與200 μg/mL AOPPs共同作用24 h。為明確自噬在AOPPs導(dǎo)致HK-2細(xì)胞肥大中的作用,將細(xì)胞分為3組:(1)Ctrl組;(2)AOPPs組:200 μg/mL AOPPs;(3)AOPPs+雷帕霉素組:100 nmol/L雷帕霉素預(yù)處理1 h后,與200 μg/mL AOPPs共同作用24 h。
1.2.4RT-qPCR 按實(shí)驗(yàn)分組處理HK-2細(xì)胞后,用Trizol提取各樣本細(xì)胞總RNA,于核酸分析儀中檢測(cè)RNA的濃度和純度。參照試劑盒說明操作反轉(zhuǎn)錄合成cDNA。采用SYBR Premix Ex TaqTM熒光定量PCR試劑盒檢測(cè),管家基因GAPDH作為內(nèi)參,引物序列見表1,反應(yīng)條件為95 ℃預(yù)變性10 min,95 ℃變性10 s,60 ℃退火延伸30 s,擴(kuò)增40個(gè)循環(huán)。上述實(shí)驗(yàn)重復(fù)3次,采用2-DDCt法進(jìn)行統(tǒng)計(jì)學(xué)分析。
1.2.5單位細(xì)胞蛋白含量測(cè)定 按實(shí)驗(yàn)分組處理HK-2細(xì)胞后,胰酶消化細(xì)胞取出微量細(xì)胞混懸液稀釋后在細(xì)胞計(jì)數(shù)板下計(jì)數(shù),RIPA裂解液裂解細(xì)胞提取細(xì)胞總蛋白,梯度稀釋0.2 μg/μL蛋白標(biāo)準(zhǔn)品,向96孔板中加入標(biāo)準(zhǔn)品或待測(cè)樣品,加入BCA工作液200 μL (A液∶B液為 50∶1)。將酶標(biāo)板振蕩30 s后,37 ℃避光孵育30 min,用酶標(biāo)儀測(cè)定562 nm波長處吸光度值。用Excel繪制標(biāo)準(zhǔn)曲線方程,要求r2>0.995,根據(jù)所測(cè)樣品的吸光值和回歸方程,計(jì)算出待測(cè)樣品的蛋白濃度(μg/μL)和總蛋白含量,用總蛋白含量和總細(xì)胞數(shù)量的比值表示單位細(xì)胞總蛋白含量。
1.2.6Western blot 按實(shí)驗(yàn)分組處理HK-2細(xì)胞后,用RIPA裂解液提取細(xì)胞總蛋白,采用BCA法測(cè)定蛋白濃度,配平上樣量和上樣體積。取20~50 μg總蛋白行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE)后,轉(zhuǎn)膜至聚偏氟乙烯(PVDF)膜。用5%脫脂奶粉室溫封閉2 h后,放入稀釋的一抗中, 4 ℃孵育過夜。一抗稀釋比分別為LC3B (1∶500),Beclin1 (1∶1 000),p62 (1∶2 000),p27 (1∶500),磷酸化-p38 MAPK (Thr180/Tyr182) (1∶1 000),p38 MAPK (1∶1 000),GAPDH (1∶5 000)。回收一抗,TBST洗膜10 min×3次。二抗(稀釋比1∶10 000)室溫孵育1 h后TBST洗膜10 min×3次,ECL顯色。以GAPDH作為內(nèi)參,用Image J軟件進(jìn)行條帶定量。

表1 基因引物序列及擴(kuò)增產(chǎn)物大小
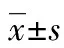
2 結(jié) 果
2.1AOPPs抑制HK-2自噬 根據(jù)文獻(xiàn)[5]報(bào)道和前期研究[2]結(jié)果,筆者將200 μg/mL未修飾的BSA或者200 μg/mL AOPPs分別刺激HK-2細(xì)胞24 h。RT-qPCR和Western blot結(jié)果顯示,AOPPs刺激使HK-2細(xì)胞Beclin1基因和蛋白表達(dá)降低而p62基因和蛋白的表達(dá)增高。此外,Western blot結(jié)果顯示,AOPPs作用下,LC3-Ⅰ向LC3-Ⅱ的轉(zhuǎn)化減少(LC3-Ⅱ/LC3-Ⅰ),P<0.05,而未經(jīng)修飾的BSA組與Ctrl組相比上述三者表達(dá)差異無統(tǒng)計(jì)學(xué)意義(P>0.05),見圖1,提示AOPPs可能抑制HK-2自噬。

A:Beclin1 mRNA 表達(dá)分析圖;B:p62 mRNA表達(dá)分析圖;C:LC3-Ⅱ/LC3-Ⅰ、Beclin1、p62蛋白Western blot條帶;D:LC-3Ⅱ/LC3-Ⅰ蛋白表達(dá)分析圖;E:Beclin1蛋白表達(dá)分析圖;F:p62蛋白表達(dá)分析圖;*:P<0.05,與Ctrl組比較
圖1 AOPPs抑制HK-2自噬
2.2P38 MAPK信號(hào)通路介導(dǎo)AOPPs抑制HK-2自噬 為進(jìn)一步觀察AOPPs抑制HK-2自噬是否與其激活p38 MAPK信號(hào)通路有關(guān),本研究使用磷酸化p38 MAPK特異性抑制劑SB203580與AOPPs共同刺激HK-2細(xì)胞。與前期研究[4,6]一致,AOPPs導(dǎo)致p38 MAPK磷酸化,而10 μmol/L SB203580抑制該通路(圖2A、B,P<0.05)。與AOPPs組相比,AOPPs+SB203580組自噬相關(guān)基因和蛋白Beclin1和p62表達(dá)分別上調(diào)和下調(diào),LC3-Ⅱ/LC3-Ⅰ也增高(圖2C~F,P<0.05),說明磷酸化p38 MAPK特異性抑制劑可部分逆轉(zhuǎn)AOPPs對(duì)HK-2細(xì)胞的自噬抑制作用,因而提示p38 MAPK信號(hào)通路介導(dǎo)了AOPPs抑制HK-2自噬。
2.3自噬抑制介導(dǎo)AOPPs導(dǎo)致HK-2肥大 本課題組前期研究[2]證明AOPPs導(dǎo)致HK-2肥大。為明確AOPPs誘導(dǎo)細(xì)胞肥大的機(jī)制是否與自噬抑制有關(guān),根據(jù)文獻(xiàn)[7]報(bào)道,筆者使用100 nmol/L雷帕霉素增強(qiáng)HK-2細(xì)胞自噬,進(jìn)一步觀察了AOPPs誘導(dǎo)的肥大是否隨之改善。本研究顯示,與AOPPs組相比,AOPPs+雷帕霉素組單位細(xì)胞蛋白含量減少,p27基因和蛋白表達(dá)降低(P<0.05),見圖3,說明自噬增加后HK-2肥大減輕。提示自噬抑制可能為AOPPs導(dǎo)致HK-2肥大的機(jī)制之一。

A:p-p38/p38蛋白Western blot條帶;B:p-p38蛋白表達(dá);C:Beclin1、p62 mRNA表達(dá);D:LC3Ⅱ/Ⅰ、Beclin1、p62蛋白Western blot條帶;E:LC3Ⅱ/Ⅰ;F:Beclin1、p62蛋白表達(dá)分析圖灰度值與p38灰度值的比值柱形圖;1:Ctrl組;2:BSA組;3:AOPPs組;4:AOPPs+SB203580組;*:P<0.05,與Ctrl組比較;#:P<0.05,與AOPPs組比較
圖2 p38 MAPK信號(hào)通路介導(dǎo)AOPPs抑制HK-2自噬

A:細(xì)胞總蛋白含量/細(xì)胞數(shù);B:p27 mRNA 表達(dá);C:p27蛋白Western blot條帶;D:p27蛋白表達(dá);1:Ctrl組;2:AOPPs組;3:AOPPs+雷帕霉素組;*:P<0.05,與Ctrl組比較;#:P<0.05,與AOPPs組比較
圖3 AOPPs抑制自噬介導(dǎo)HK-2肥大
3 討 論
本研究發(fā)現(xiàn)AOPPs抑制HK-2自噬,并提示自噬抑制受p38 MAPK信號(hào)通路調(diào)控,且自噬抑制參與了AOPPs導(dǎo)致的HK-2肥大。
HK-2肥大是腎小管細(xì)胞遇到急慢性應(yīng)激反應(yīng)時(shí)的早期表現(xiàn),其主要表現(xiàn)為蛋白和mRNA水平的升高,細(xì)胞阻滯在G1期[8]。細(xì)胞周期受周期蛋白的調(diào)控,p27為周期抑制蛋白家族成員之一,在HK-2肥大時(shí)可觀察到p27上調(diào)[9]。本研究發(fā)現(xiàn)HK-2肥大與自噬抑制有關(guān)。
自噬是一個(gè)動(dòng)態(tài)的生物學(xué)過程,包括自噬起始,自噬體與溶酶體融合及在溶酶體內(nèi)的降解[10-11]。近年來大量的研究報(bào)道,自噬參與DN、藥物性腎損害、遺傳性腎臟病、腎臟缺血再灌注等腎臟疾病的發(fā)生、發(fā)展。其中,在自噬參與DN進(jìn)展的研究方面,Barbosa等[12]早在1992年就已證明DN中HK-2自噬功能受損。Kitada等[13]用高脂建立小鼠的DN模型,發(fā)現(xiàn)其HK-2內(nèi)長壽蛋白p62/SQSTM1堆積,證實(shí)2型糖尿病導(dǎo)致的DN中,HK-2的自噬受到抑制,而限制飲食后,觀察到自噬增加,DN癥狀得到改善。在研究不同刺激物對(duì)HK-2自噬的影響方面,Liu等[14]的研究證明從微小病變腎病綜合征患者中提取的尿蛋白體外刺激HK-2,表現(xiàn)為自噬起始激活。而在DN患者體內(nèi)明顯升高的晚期糖基化終末產(chǎn)物(AGEs)則使HK-2自噬起始激活,而自噬在溶酶體降解階段流量受阻,自噬被抑制[15]。而Nolin等[16]的研究表明,尿蛋白能抑制自噬體和溶酶體的融合,抑制自噬。本研究結(jié)果表明,AOPPs使HK-2中LC3-Ⅱ/LC3-Ⅰ、Beclin1降低,p62升高,提示在AOPPs的作用下,自噬可能在起始環(huán)節(jié)受到抑制。
自噬起始受到各種信號(hào)通路的精密調(diào)節(jié),其中p38 MAPK對(duì)自噬起雙重調(diào)節(jié)作用[17]。一方面,Paillas等[18]的研究證明,p38 MAPK磷酸化誘導(dǎo)自噬導(dǎo)致化療藥物耐藥;另一方面,Cruz-Morcillo等[19]研究證明化療藥物五氟尿嘧啶(5-FU)通過抑制p38 MAPK通路磷酸化激活自噬。最近的研究表明,AOPPs激活HK-2 p38 MAPK信號(hào)通路[4]。于是本研究進(jìn)一步觀察了AOPPs抑制自噬是否與p38 MAPK信號(hào)通路有關(guān)。研究表明抑制p38通路,可以觀察到自噬上調(diào),表明p38 MAPK信號(hào)通路介導(dǎo)了AOPPs抑制HK-2自噬。
本實(shí)驗(yàn)發(fā)現(xiàn)AOPPs激活p38 MAPK信號(hào)通路抑制HK-2自噬,自噬抑制介導(dǎo)AOPPs導(dǎo)致的HK-2胞肥大,進(jìn)一步闡明了DN的進(jìn)展機(jī)制。然而仍有許多信號(hào)通路有效調(diào)控自噬活動(dòng),尚待進(jìn)一步研究。
[1]Kalousova M,Skrha J,Zima T.Advanced glycation end-products and advanced oxidation protein products in patients with diabetes mellitus[J].Physio Res,2002,51(6):597-604.
[2]Tang X,Rong G,Bu Y,et al.Advanced oxidation protein products induce hypertrophy and epithelial-to-mesenchymal transition in human proximal tubular cells through induction of endoplasmic reticulum stress[J].Cell Physiol Biochem,2015,35(2):816-828.
[3]Yang YP,Liang ZQ,Gu ZL,et al.Molecular mechanism and regulation of autophagy[J].Acta Pharmacol Sin,2005,26(12):1421-1434.
[4]黃麗麗,祝小林,鄧偉謙,等.p38 MAPK信號(hào)通路介導(dǎo)晚期氧化蛋白產(chǎn)物誘導(dǎo)的腎小管上皮細(xì)胞間充質(zhì)轉(zhuǎn)分化[J].南方醫(yī)科大學(xué)學(xué)報(bào),2016,36(9):1209-1214.
[5]Witko-Sarsat V,Nguyen-Khoa T,Jungers P,et al.Advanced oxidation protein products as a novel molecular basis of oxidative stress in uraemia[J].Nephrol Dial Transplant,1999,14 Suppl 1:76-78.
[6]Webber JL.Regulation of autophagy by p38 alpha MAPK[J].Autophagy,2010,6(2):292-293.
[7]Xu Y,Liu L,Xin W,et al.The renoprotective role of autophagy activation in proximal tubular epithelial cells in diabetic nephropathy[J].J Diabetes Complications,2015,29(8):976-983.
[8]Preisig PA,Franch HA.Renal epithelial cell hyperplasia and hypertrophy[J].Semin Nephrol,1995,15(4):327-340.
[9]Shankland SJ,Wolf G.Cell cycle regulatory proteins in renal disease:role in hypertrophy,proliferation,and apoptosis[J].Am J Physiol Renal Physiol,2000,278(4):F515-F529.
[10]Boya P,Reggiori F,Codogno P.Emerging regulation and functions of autophagy[J].Nat Cell Biol,2013,15(7):713-720.
[11]Wang ZB,Choi ME.Autophagy in kidney health and disease[J].Antioxid Redox Signal,2014,20(3):519-537.
[12]Barbosa JA,Zhou H,Hultenschmidt D,et al.Inhibition of cellular autophagy in proximal tubular cells of the kidney in streptozotocin-diabetic and uninephrectomized rats[J].Virchows Arch B Cell Pathol Incl Mol Pathol,1992,61(6):359-366.
[13]Kitada M,Takeda A,Nagai T,et al.Dietary restriction ameliorates diabetic nephropathy through Anti-Inflammatory effects and regulation of the autophagy via restoration of Sirt1 in diabetic wistar fatty (fa/fa) rats:a model of type 2 diabetes[J].Exp Diabetes Res,2011:908185.
[14]Liu WJ,Luo MN,Tan J,et al.Autophagy activation reduces renal tubular injury induced by urinary proteins[J].Autophagy,2014,10(2):243-256.
[15]Liu WJ,Shen TT,Chen RH,et al.Autophagy-Lysosome pathway in renal tubular epithelial cells is disrupted by advanced glycation end products in diabetic nephropathy[J].J Biol Chem,2015,290(33):20499-20510.
[16]Nolin AC,Mulhern RM,Panchenko MV,et al.Proteinuria causes dysfunctional autophagy in the proximal tubule[J].Am J Physiol Renal Physiol,2016,311(6):F1271-F1279.
[17]Sui XB,Kong N,Ye L,et al.p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents[J].Cancer Lett,2014,344(2):174-179.
[18]Paillas S,Causse A,Marzi L,et al.MAPK14/p38 alpha confers irinotecan resistance to TP53-defective cells by inducing survival autophagy[J].Autophagy,2012,8(7):1098-1112.
[19]Cruz-Morcillo LA,Valero ML,Callejas-Valera JL,et al.P38MAPK is a major determinant of the balance between apoptosis and autophagy triggered by 5-fluorouracil:implication in resistance[J].Oncogene,2012,31(9):1073-1085.
Advancedoxidationproteinproductsmodulateautophagyofrenaltubuleepithelialcells*
ZhangJun,XiangXiaohong,LiangXiujie,ShuShuangshuang,JiangTingting,GuoTingting,TangXun△
(DepartmentofNephrology,ZhujiangHospitalofSouthernMedicalUniversity,Guangzhou,Guangdong510280,China)
ObjectiveTo investigate the effect of advanced oxidation protein products (AOPPs) on human renal tubular epithelial cells(HK-2) autophagy.MethodsHK-2 cells were stimulated with AOPPs.RT-qPCR and Western blot were used to determine the expression of autophagy related protein LC3-Ⅱ/LC3-Ⅰ,Beclin1 and p62;Western blot was utilized to examine the activation of p38 MAPK pathway.Then p38 MAPK inhibitor (SB203580) was added and co-processed with AOPPs.The change of autophagy was observed.Also,autophagy inducer rapamycin was added and co-processed with AOPPs.RT-qPCR and Western blot were used to detect the expression of cell cycle inhibitory protein p27.The cell total protein level was detected by the bicinchoninic acid (BCA) method.The hypertrophy change was observed.ResultsAOPPs down-regulated the expression of LC3-Ⅱ/LC3-Ⅰ and Beclin1,up-regulated expression of p62 and activated p38 MAPK pathway;in comparison with the AOPPs alone treatment group,the expression of LC3-Ⅱ/LC3-Ⅰ and Beclin in the SB203580 co-processing group was increased,while p62 was decreased;the p27 expression and cells total protein in the sirolimus co-processing group were down-regulated.ConclusionAOPPs inhibits the autophagy of HK-2 cells by activating p38 MAPK pathway and the autophagy inhibition participates in HK-2 cell hypertrophy.
autophagy;p38 MAPK signaling pathway;advanced oxidation protein products
R692.5
A
1671-8348(2017)25-3480-04
2016-12-15
2017-06-09)
國家自然科學(xué)基金資助項(xiàng)目(81202842)。
章俊(1966-),博士,主任醫(yī)師,主要從事糖尿病腎病、晚期氧化蛋白產(chǎn)物的研究。
△通信作者,E-mail:1847672118@qq.com。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
