廣東淡水魚類弓背青鳉分類形態(tài)性狀再描述
2017-10-12 10:46:20張方方姜昊辰金錦錦仇玉萍陳國柱
四川動物 2017年5期
張方方, 姜昊辰, 金錦錦, 仇玉萍, 陳國柱
(西南林業(yè)大學,國家高原濕地研究中心,昆明650224)
廣東淡水魚類弓背青鳉分類形態(tài)性狀再描述
張方方, 姜昊辰, 金錦錦, 仇玉萍, 陳國柱*
(西南林業(yè)大學,國家高原濕地研究中心,昆明650224)
食蚊魚Gambusiaaffinis的入侵導致弓背青鳉Oryziascurvinotus種群嚴重下降而趨于瀕危。其模式標本產(chǎn)地種群(海南島那大)已經(jīng)滅絕,野外種群已多年未見報道。過去,研究者對正模標本的描述存在嚴重錯誤且難于復檢,現(xiàn)亟需對其分類性狀進行再描述及模式標本重建。本文對近期采自廣州南沙島的弓背青鳉孑遺種群進行了分類形態(tài)性狀再描述,補充了活體及浸制標本彩色照片,并對其雌雄間的形態(tài)差異進行了統(tǒng)計分析。結果顯示,弓背青鳉區(qū)別于其他青鳉魚類的主要分類檢索性狀是臀鰭條數(shù)目為19.60±0.80(19~22)、體側縱列鱗數(shù)目為28.51±0.67(27~30),定種的關鍵特征是背鰭-吻端/體長:雄魚為(80.9±1.3)%、雌魚為(82.2±1.0)%、全部為(81.6±1.3)%;同時,通過單因素方差分析、主成分分析及判別分析等多元統(tǒng)計方法證實了其種內存在的雌雄異形現(xiàn)象,文中回顧并澄清了有關弓背青鳉的分類形態(tài)性狀問題。當前亟待加強弓背青鳉保護生物學研究并應采取有力的保護措施盡早對現(xiàn)存種群進行有效保護。
弓背青鳉; 形態(tài)性狀; 雌雄異形
Abstract: The population of ricefish (Oryziascurvinotus) had been significantly declined since the invasive species mosquitofish (Gambusiaaffinis) widely introduced to China during the recent few decades. As a result, the population of this species in type locality had been extinct. Unfortunately, the researcher had made some big mistake in the description of its holotype, and thus brought big confusions to the researchers during their identification to theOryziasspecies in China. In order to rebuild the holotype, relocate the type locality and amend the characteristics descriptions of thisOryziasspecies, in this paper, we conducted a characteristic re-description ofO.curvinotuswhich were collected from the Huangshanlu National Forest Park and its adjoin zone (Guangzhou city, China), and replenished the chromophotograph of this species. The result showed that the principal identification characteristics ofO.curvinotusfrom otherOryziasspecies should be as following: anal-fin rays are 19.60±0.80 (19-22), scales in a lateral series are 28.51±0.67 (27-30) and the ratio of pre-dorsal length/standard length is (81.6±1.3)% in total. Meanwhile, the sexual dimorphism of this species also was determined by one way analysis of variance, principal component analysis and discriminant analysis. As theO.curvinotuswas greatly threatened by the invasion of mosquitofish, and the invader had also been found in the sites whereO.curvinotuspopulations still remained, we strongly suggest that effective measures are urgently needed to protect the relict populations of this species.
Keywords:Oryziascurvinotus; morphological characteristics; sexual dimorphism
青鳉是廣泛分布于亞洲東部、東南部中南半島及印度尼西亞群島的小型魚類,成熟個體體長多在20~30 mm,因過去在稻田生境中極為常見而被稱為稻田魚,在分類上屬鳉形目Cyprinodontiformes大頜鳉科Adrianichthyidae青鳉屬Oryzias。迄今為止,被描述過的物種有20多種(Parenti,2008;Herder & Chapuis,2010;Magtoon,2010;Parenti & Hadiaty,2010;Asaietal.,2011;Herderetal.,2012;Parentietal.,2013;Mokodonganetal.,2014),并可能還存在許多有待鑒定的新物種。我國已記述的有4種,分別為中華青鳉Oryziassinensis、小青鳉O.minutillus、弓背青鳉O.curvinotus和日本青鳉O.latipes,而日本青鳉很可能是中華青鳉誤記(陳銀瑞等,1989),因而《中國動物志 硬骨魚綱 鳉形目》僅收錄了前3種。過去,除陳銀瑞等(1989)對云南青鳉魚類進行過調查外,我國境內究竟存在多少青鳉魚類物種尚未進行過深入調查,其中對弓背青鳉的記述尤為簡略。《中國動物志 硬骨魚綱 鳉形目》對弓背青鳉性狀的描述摘錄于Nichols和Pope(1927)對正模標本所作原始描述,缺乏對實物標本的檢視。對研究者早期采集的青鳉樣品檢視后,Uwa和Parenti(1988)指出Nichols和Pope(1927)對弓背青鳉模式標本臀鰭條數(shù)目原始描述出現(xiàn)的問題可能為誤記,Parenti(2008)更進一步在對20余種青鳉魚類分類性狀總結中更正弓背青鳉臀鰭條數(shù)為17~20,體側縱列鱗數(shù)為27~28。因此,由于Nichols和Pope(1927)對弓背青鳉鰭條數(shù)目的描述存在錯誤,《中國動物志 硬骨魚綱 鳉形目》檢索表的編制中將弓背青鳉的臀鰭數(shù)引述為25,體側縱列鱗數(shù)為35,也是錯誤的。弓背青鳉模式標本產(chǎn)地原為我國海南島那大(現(xiàn)儋州那大鎮(zhèn),1957年為那大縣,解放前為那恁村和大同村),近代以來由于棲息地的破壞及食蚊魚Gambusiaaffinis的廣泛入侵,弓背青鳉模式標本產(chǎn)地種群可能已滅絕,多年來弓背青鳉在我國海南島區(qū)域的野外種群已無報道。另一方面,Nichols和Pope(1927)描述所依據(jù)的標本僅1尾(未見副模標本表述),弓背青鳉模式標本過去雖產(chǎn)自我國海南島但保存于美國,距今年代久遠,是否尚存不得而知,今人無法對其進行復檢。盡管如此,Nichols和Pope(1927)對弓背青鳉關鍵特征,背鰭起點極為靠后的描述是成立的,因此該物種名被研究者承認(Uwa & Parenti,1988;Parenti,2008)。筆者于2016年8月在廣州南沙區(qū)南沙街道南沙島采集了一批青鳉魚類標本,經(jīng)鑒定確認為弓背青鳉,其形態(tài)性狀及關鍵分類特征與Uwa和Parenti(1988)、Parenti(2008)等諸多研究者的記述接近,而與《中國動物志 硬骨魚綱 鳉形目》的記述存在較大差異。鑒于正模標本的描述存在嚴重錯誤且難于復檢,且地模標本也無法獲得,現(xiàn)亟需對其分類性狀進行再描述及模式標本重建。
過去一些研究者對產(chǎn)于廣東的青鳉魚類曾作記述,但迄今為止尚未能明確有哪些物種,先后記述過的物種有日本青鳉(潘炯華等,1984;趙俊等,2010)、中華青鳉(賈銀濤等,2013;林小濤,張潔,2013)、弓背青鳉(Uwa & Parenti,1988;Wangetal.,2017)。本文通訊作者于2006年、2008年、2010年分別在上述文獻所記述的韶關曲江地區(qū)(潘炯華等,1984)、廣州從化地區(qū)流溪河流域(趙俊等,2010)、廣州增城增江流域(賈銀濤等,2013;林小濤,張潔,2013)采集到青鳉魚類標本,經(jīng)檢定應分屬2種不同魚類,前者應為鰭斑青鳉O.pectoralis,而后兩者應為弓背青鳉。而Uwa和Parenti(1988)研究中由于沒有具體地點而無法檢視,在其所記載區(qū)域周邊如廣州南沙、梧州鄰近區(qū)域肇慶市懷集縣近郊分別采得弓背青鳉。鰭斑青鳉與青鳉屬的其他種類關鍵特征在于其胸鰭基部前有一顯著黑斑。目前可以確定,廣東區(qū)域內至少存在2個青鳉魚類物種,分別為弓背青鳉和鰭斑青鳉,且均未作詳細的分類學描述,亟待補充相關數(shù)據(jù)。本文將對產(chǎn)自廣州南沙島的弓背青鳉進行分類性狀描述,且對其雌雄異形進行統(tǒng)計,以彌補我國對弓背青鳉分類資料的缺失,為進一步整理我國青鳉魚類分類問題積累基礎資料。
1 材料與方法
1.1樣品采集地
樣品采集地位于廣東省廣州市南沙區(qū)南沙街道南沙島[該區(qū)2005年建區(qū),從原番禺區(qū)劃出,該地為原廣州市番禺縣(區(qū))南沙鎮(zhèn)所在地],面積約100 km2。采集區(qū)域為該地中央黃山魯森林公園東側溪流,以所在地名稱暫命名為九王廟溪、時代南灣溪、盧灣涌等,共發(fā)現(xiàn)3個種群。這3條溪流最終均匯入珠江口北部伶仃洋水域,流程均不超過5 km。
1.2樣品采集與實驗室分析
2016年8月22—24日及10月2—5日進行采樣。利用網(wǎng)孔為2 mm,網(wǎng)口直徑30 cm的手抄網(wǎng)進行收集,所得標本用5%福爾馬林溶液固定備檢。本研究標本共51尾,30雌21雄,均為可辨別的性成熟個體。
在實驗室對青鳉樣品的可數(shù)性狀進行計數(shù)及對可量性狀進行測量,計數(shù)及測量性狀的選取主要依據(jù)《中國動物志 硬骨魚綱 鳉形目》所述。涉及的可數(shù)性狀參數(shù)主要有背鰭、臀鰭、胸鰭、腹鰭、尾鰭的鰭條數(shù)目以及體側縱列鱗數(shù)目等;可量性狀參數(shù)主要有體長、體高、背鰭-吻端、臀鰭始點-吻端、肛門-吻端、頭長、吻長、眼徑、眼間距、口寬、最長背鰭條、最長臀鰭條、胸鰭長、腹鰭長、尾鰭長、尾柄長、尾柄高等(圖1:A)。為進一步對弓背青鳉雌雄異形問題進行分析,采用框架測量方法對其形態(tài)進一步測量(圖1:B),數(shù)據(jù)采用多元統(tǒng)計方法分析。
特別說明,本文采用《中國動物志 硬骨魚綱 鳉形目》對青鳉魚類鰭條數(shù)目的處理方法,將不分枝鰭條和分枝鰭條數(shù)合并計算鰭條數(shù)目而不分開描述。同時,由于弓背青鳉存在雌雄異形現(xiàn)象,本文在性狀描述中先合并描述,再將雌魚、雄魚分開描述。
1.3數(shù)據(jù)分析
弓背青鳉可數(shù)及可量性狀數(shù)據(jù)均以平均值±標準差表示,利用SPSS 19.0進行統(tǒng)計分析。對雌雄魚各參數(shù)間的差異顯著性采用單因素方差分析(ANOVA)。可量性狀在分析前與體長或頭長作比,以比值進行數(shù)據(jù)分析,以消除各可量性狀參數(shù)因體長依賴所引起的系統(tǒng)誤差。
另一方面,基于上述傳統(tǒng)測量性狀及框架測量性狀共計57組可量性狀比例參數(shù),利用多元統(tǒng)計方法主成分分析、判別分析等對性別間的形態(tài)性狀差異進行深入分析,以確認雌雄魚是否存在形態(tài)性狀差異。
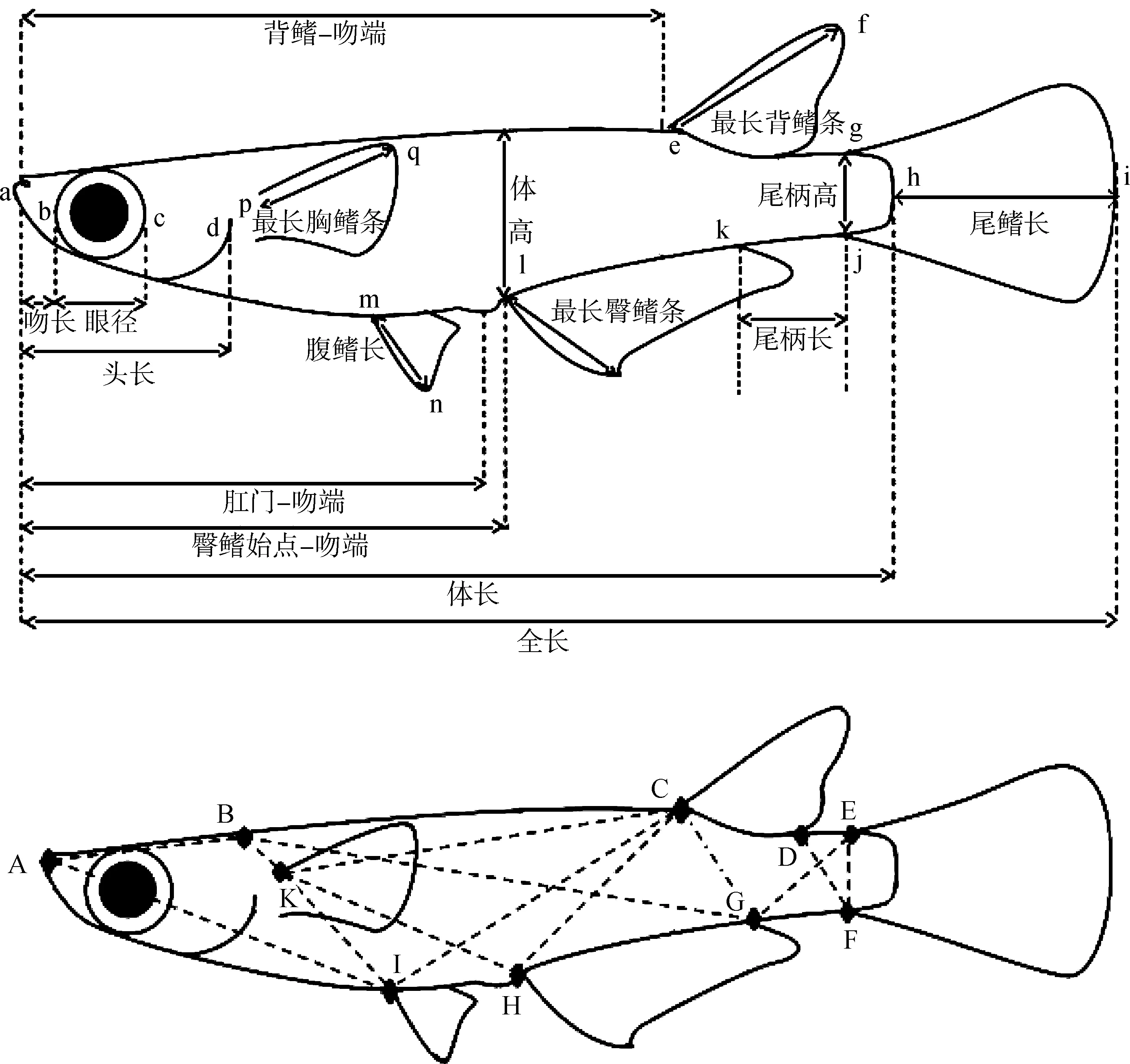
圖1 弓背青鳉測量性狀示意(仿Magtoon, 2010)Fig. 1 Measurements of Oryzias curvinotus (redrawing from Magtoon, 2010)
2 結果
2.1形態(tài)與性狀描述
弓背青鳉體修長,眼大,口上位(圖2)。在野外水體中,從背面觀察可見其頭頂部至背鰭前端呈狹長“V”字型亮銀白色反光,背鰭后至尾柄呈亮藍色反光。在水族缸中觀察時體呈淡青黃色;體透明,可觀察到脊柱骨及主要血管分布;腹部呈灰銀白色。尾鰭正截形,無分叉。各鰭第一鰭條側常見較淺的淡黃色,尤其尾鰭上下兩側淡黃色較為明顯。8月采樣期間正值其繁殖期,雄性間相互爭斗并追逐雌魚,雌雄魚腹鰭、臀鰭沒有中華青鳉所呈現(xiàn)的明顯黑灰色。雄魚臀鰭展開呈平行四邊形(圖2:b)。浸制標本體側有一黑色縱線(圖2),未發(fā)現(xiàn)側線。作為其物種特征之一,背部隆起呈彎弓狀這一形態(tài)特征只在成年個體(尤其是雄魚)中表現(xiàn)出來,幼魚、未性成熟的成體及雌魚多數(shù)個體該特征不明顯。背鰭-吻端/體長為(81.6±1.3)%,這是與其他青鳉魚類區(qū)別及種名確定的關鍵特征。雄魚臀鰭、背鰭最后鰭條明顯短小,鰭條膜呈現(xiàn)顯著凹缺(圖2:b,d)。最長臀鰭條出現(xiàn)在臀鰭后部。
可數(shù)性狀(表1):背鰭條數(shù)目平均6.04±0.20,變動范圍6~7;臀鰭條數(shù)目19.60±0.80,變動范圍19~22,中值19;胸鰭條數(shù)目8.69±0.47,變動范圍8~9;腹鰭條數(shù)目6.00±0.00;尾鰭條數(shù)目18.88±1.31,變動范圍17~21;體側縱列鱗數(shù)目28.51±0.67,變動范圍27~30,中值29。除尾鰭條數(shù)目外,雌雄魚間各測量的可數(shù)性狀差異無統(tǒng)計學意義。
可量性狀(表1):體長20.28 mm±1.91 mm,變動范圍15.82~24.55 mm,為體高的(4.11±0.25)倍(3.68~4.56倍),為背鰭-吻端的(1.23±0.02)倍(1.19~1.26倍),為臀鰭始點-吻端的(1.67 ± 0.07)倍(1.48~1.82倍),為肛門-吻端的(1.74±0.06)倍(1.63~1.86倍),為頭長的(3.96±0.16)倍(3.56~4.29倍)。頭長為吻長的(4.15±0.25)倍(3.65~4.70倍),為眼徑的(2.77±0.17)倍(2.42~3.24倍),為眼間距的(3.85±0.33)倍(2.88~4.55倍),為口寬的(2.60±0.15)倍(2.27~3.02倍),為最長背鰭條的(1.42±0.22)倍(1.03~1.92倍),為最長臀鰭條的(1.57±0.19)倍(1.16~1.95倍),為胸鰭長的(1.23±0.10)倍(1.02~1.43倍),為腹鰭長的(1.90±0.18)倍(1.58~2.26倍),為尾鰭長的(1.05±0.04)倍(0.096~1.14倍),為尾柄長的(2.36±0.18)倍(2.09~2.99倍)。尾柄長為尾柄高的(1.03±0.08)倍(0.73~1.22倍)。
雄魚。體長20.27 mm±2.20 mm,變動范圍16.32~24.55 mm,為體高的(4.33±0.17)倍(4.02~4.56倍),為背鰭-吻端的(1.24±0.02)倍(1.19~1.26倍),為臀鰭始點-吻端的(1.73±0.05)倍(1.63~1.82倍),為肛門-吻端的(1.78±0.05)倍(1.70~1.86倍),為頭長的(3.89±0.15)倍(3.67~4.15倍)。頭長為吻長的(4.16±0.21)倍(3.71~4.52倍),為眼徑的(2.72±0.18)倍(2.42~3.07倍),為眼間距的(3.89±0.38)倍(2.88~4.54倍),為口寬的(2.68±0.13)倍(2.40~2.88倍),為最長背鰭條的(1.19±0.08)倍(1.03~1.34倍),為最長臀鰭條的(1.38±0.14)倍(1.16~1.65倍),為胸鰭長的(1.24±0.07)倍(1.15~1.43倍),為腹鰭長的(2.06±0.10)倍(1.87~2.26倍),為尾鰭長的(1.05±0.04)倍(0.98~1.14倍),為尾柄長的(2.33±0.20)倍(2.11~2.99倍)。尾柄長為尾柄高的(1.04±0.10)倍(0.73~1.22倍)。

圖2 弓背青鳉Fig. 2 Oryzias curvinotus
(a)活體雌魚; (b) 活體雄魚; (c) 5%福爾馬林固定1周后的雌魚; (d) 5%福爾馬林固定1周后的雄魚; 比例尺=1 cm。
(a) alive female; (b) alive male; (c) preserved by 5% formalin solution one week later, female; (d) preserved by 5% formalin solution one week later, male; scale bar=1 cm.
雌魚。體長20.28 mm±1.72 mm,變動范圍15.82~22.78 mm,為體高的(3.95±0.16)倍(3.68~4.25倍),為背鰭-吻端的(1.22±0.01)倍(1.19~1.26倍),為臀鰭始點-吻端的(1.64±0.05)倍(1.48~1.72倍),為肛門-吻端的(1.70±0.04)倍(1.63~1.79倍),為頭長的(4.00±0.15)倍(3.56~4.29倍)。頭長為吻長的(4.14±0.28)倍(3.65~4.70倍),為眼徑的(2.81±0.16)倍(2.53~3.24倍),為眼間距的(3.82±0.30)倍(3.33~4.55倍),為口寬的(2.55±0.14)倍(2.27~23.02倍),為最長背鰭條的(1.58±0.12)倍(1.30~1.92倍),為最長臀鰭條的(1.67±0.10)倍(1.16~1.95倍),為胸鰭長的(1.22±0.17)倍(1.02~1.45倍),為腹鰭長的(1.79±0.13)倍(1.58~2.15倍),為尾鰭長的(1.06±0.05)倍(0.96~1.14倍),為尾柄長的(2.39±0.16)倍(2.09~2.75倍)。尾柄長為尾柄高的(1.02±0.07)倍(0.90~1.15倍)。
2.2雌雄形態(tài)性狀差異
2.2.1單因素方差分析在部分傳統(tǒng)分類性狀上,雌雄弓背青鳉間差異有統(tǒng)計學意義(表1)。進一步利用主成分分析也顯示了兩者間的形態(tài)性狀分離。
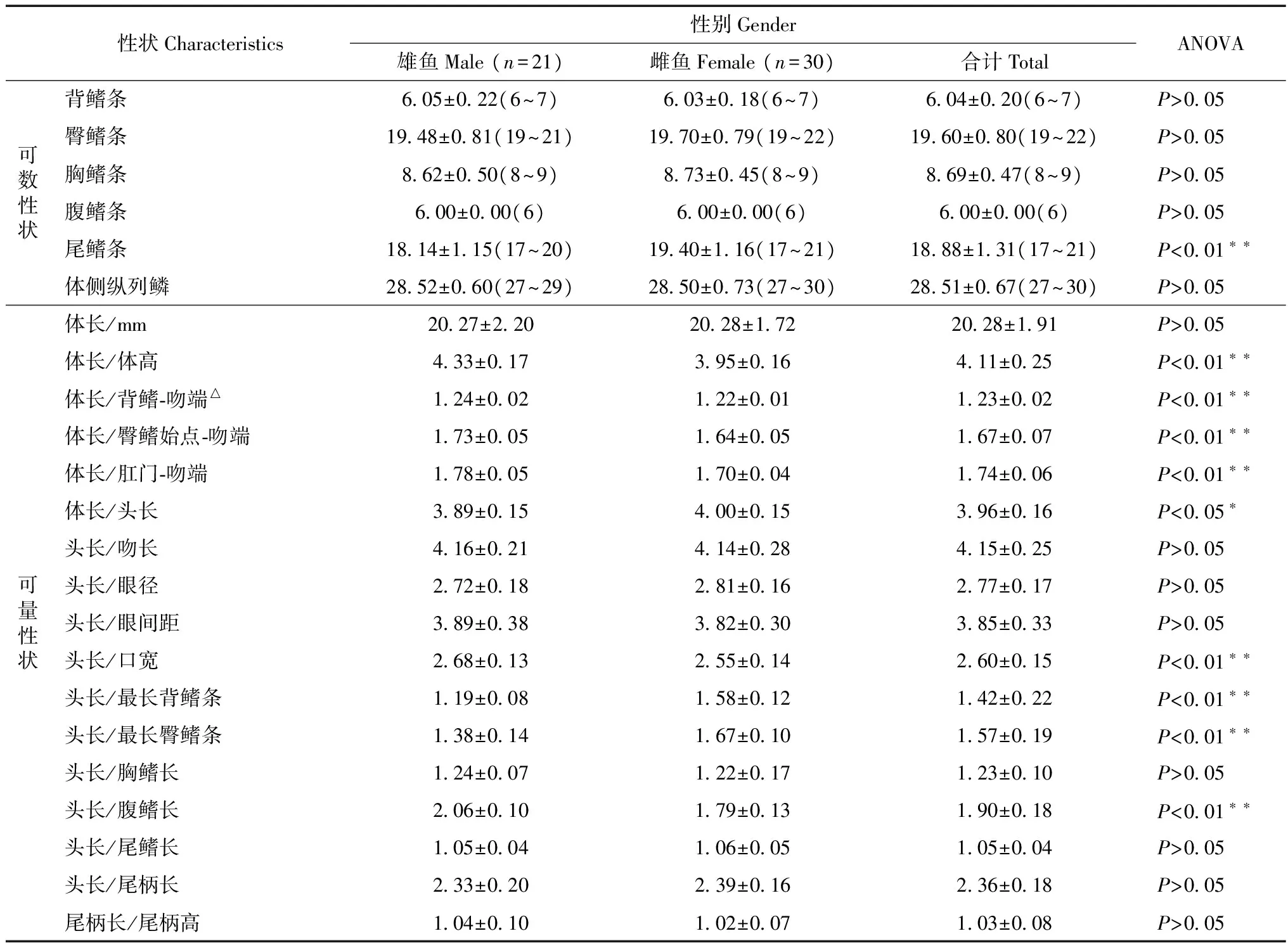
表1 弓背青鳉傳統(tǒng)可數(shù)及可量性狀Table 1 Morphometric and meristic data of Oryzias curvinotus from Nansha Island, Guangdong, China
注: 括號內為數(shù)值變動范圍;*統(tǒng)計結果在雌雄魚間差異有統(tǒng)計學意義(P<0.05),**統(tǒng)計結果在雌雄魚間差異有高度統(tǒng)計學意義(P<0.01);△背鰭-吻端/體長雄魚為(80.9±1.3)%, 雌魚為(82.2±1.0)%, 全部為(81.6±1.3)%。
Notes: The number in brackets is the range of data variation.*there is a significant difference (P<0.05) and and**there is an extremely significant difference between genders ofOryziascurvinotus(P<0.01);△the ratio of pre-dorsal length/standard length inO.curvinotusis (81.6±1.3)% in total and are (80.9±1.3)% and (82.2±1.0)% in male and female fish respectively.
2.2.2主成分分析分析結果顯示,在57組參與分析比例參數(shù)中共提取出14個主成分,它們對總變異的累積貢獻率為84.455%。其中前4個主成分對種群間形態(tài)差異的貢獻率分別為:PC1=31.637%、PC2=10.269%、PC3=6.889%、PC4=5.884%,累積貢獻率為64.679%。
PC1中體長/EH(0.901,EH為圖1中框架性狀測量點E到H的長度;下同)、體長/FH(0.868)、體長/肛門-吻端(0.839)、體長/GH(0.834)、體長/臀鰭始點-吻端(0.830)、體長/DH(0.806)、頭長/腹鰭長(0.800)、體長/體高(0.775)、體長/EG(0.729)、體長/CF(0.711)等10個比例性狀參數(shù)對PC1有主要貢獻(>0.700);PC2中體長/CI(0.764)、體長/DI(0.734)比例性狀參數(shù)對PC2有主要貢獻(>0.700)。弓背青鳉雌雄兩者在PC1軸方向明顯分離(圖3)。
2.2.3判別分析本文應用判別分析統(tǒng)計方法的主要目的是分析雌雄弓背青鳉是否存在差異及能否準確區(qū)分,故省略了判別函數(shù)表達式的表述,只呈現(xiàn)判別結果及判別效果檢驗。分析結果顯示,雌雄魚間的判別函數(shù)組質心分別為12.096和-17.280,距離較遠,兩者分離顯著。所建立的判別函數(shù)對原始分組案例中樣品正確判別率均為100.0%,交叉驗證分組案例的正確率為54.9%。
3 討論
過去對弓背青鳉的分類及相關研究主要是國外,尤其是日本。據(jù)Uwa和Parenti(1988)的記述,先后有Oshima(1926)、Nichols和Pope(1927)、Harada(1943)等對其種名確定及形態(tài)性狀描述的有爭論。Nichols和Pope(1927)作為弓背青鳉的定名者,對我國海南島模式標本產(chǎn)地那大地區(qū)個體部分性狀如臀鰭條數(shù)目的描述存在重要錯誤,但對該種的定種關鍵特征的記述是有效的。例如,弓背青鳉區(qū)別于其他青鳉魚類的關鍵特征之一是背鰭位置明顯靠后,導致外觀上背部呈隆起狀,故稱為“弓背”或“曲背”青鳉,背鰭-吻端/體長為80%~82%,而其他青鳉魚類一般為77%~79%(Uwa & Parenti,1988),該特征在本檢測種群為81.6%±1.3%,與之相符;第二特征為雄魚臀鰭展開為平行四邊形,與其他物種的三角形顯著區(qū)分(Parenti,2008),本文檢測種群與之相符。因此,從形態(tài)特征上可以確認本文所描述種群為弓背青鳉種群。而對于臀鰭條數(shù)目問題,Nichols和Pope(1927)的小失誤引起了后來的系列問題。由于早期分類中臀鰭條數(shù)目是青鳉魚類一個重要的分類檢索性狀,弓背青鳉與日本青鳉臀鰭條的數(shù)目十分相似,故Harada(1943)將采自海南的青鳉記述為日本青鳉,后來Uwa和Parenti(1988)檢視了日本學者早期在海南島采集到的青鳉標本后確定為弓背青鳉,并指出Nichols和Pope(1927)將其臀鰭條數(shù)目記述為25可能是誤記。日本學者很早就在海南島(那大及嘉積)采集到青鳉,但將之鑒定為日本青鳉,后來Nichols和Pope(1927)根據(jù)該地青鳉背鰭位置明顯靠后的特點區(qū)別于當時已知的數(shù)種青鳉魚類而定出弓背青鳉,且作為青鳉屬下一個亞屬,種名Aplocheiluscurvinotus。弓背青鳉的基本分類性狀描述在Uwa和Parenti(1988)后已經(jīng)得到澄清,并且種的有效性得到細胞生物學(Uwaetal.,1982;Uwa,1991)及基礎生物學研究(Hamaguchi & Sakaizumi,1992;Sakaizumietal.,1992)的支持。近代以來,海南島淡水魚類棲息生境由于食蚊魚廣泛入侵及近代急速城鎮(zhèn)化而被嚴重破壞,那大地區(qū)仍存有青鳉魚類種群可能性不大,這也可能是《中國動物志 硬骨魚綱 鳉形目》著者未能以實際標本進行描述的關鍵原因。本文所檢視的廣州南沙島區(qū)域數(shù)個現(xiàn)存弓背青鳉種群,可以作為重建弓背青鳉模式標本、副模標本或者等模式標本候選。
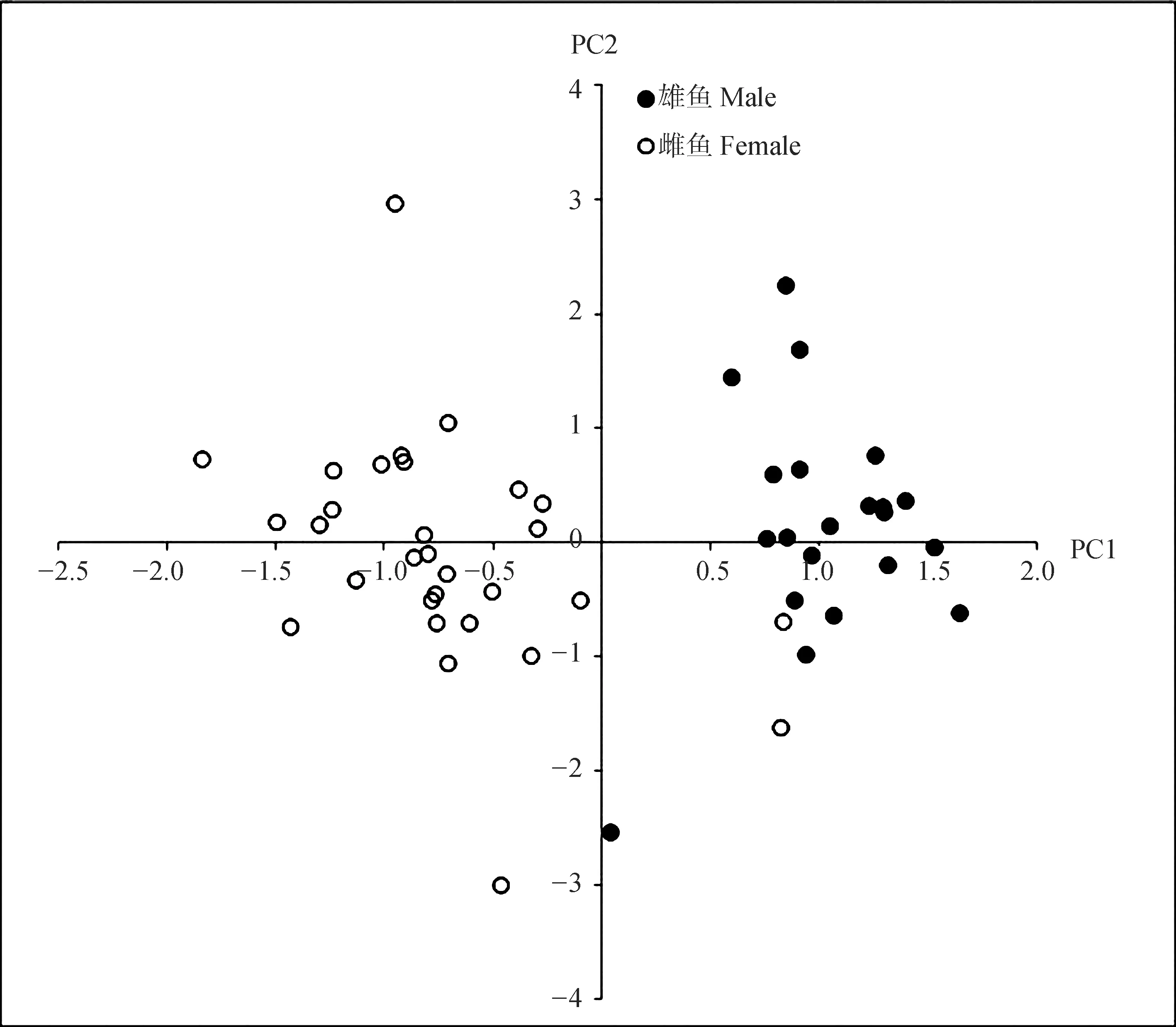
圖3 主成分PC1對PC2散點圖Fig.3 Scatter plot for PC1 vs. PC2
在近代,Uwa和Parenti(1988)對中國廣大區(qū)域的青鳉魚類進行過采集,當時是以對東亞區(qū)域日本青鳉與弓背青鳉形態(tài)與遺傳差異為研究目的,發(fā)現(xiàn)中國所產(chǎn)“日本青鳉”與日本所產(chǎn)日本青鳉形態(tài)上存在許多差別,是確定中華青鳉O.sinensis為青鳉屬新種的關鍵(Uwaetal.,1988;陳銀瑞等,1989)。在該文,作者使用了采集自廣東廣州、東莞太平、廣西梧州、香港以及海南島的標本,明確認定為弓背青鳉,由于該文沒有標注采集時間及具體地點,筆者尚無法重新檢視上述區(qū)域的青鳉魚類樣品。在關鍵分類指標臀鰭條數(shù)目上,本文所采集弓背青鳉樣品為19.60±0.80(19~22),與Uwa和Parenti(1988)所記述的采集自廣州(平均值19,變化區(qū)間19,n=3;下同)、太平(20,18~21,n=9)、梧州(18,16~19,n=32)、香港(20,18~21,n=12)、海南島(19,18~20,n=29)以及越南河內(19,18~19,n=3)等地弓背青鳉樣品基本相似(表2)。同時,本文明確證實了弓背青鳉間存在著雌雄異形現(xiàn)象。目前,弓背青鳉已知分布區(qū)域主要為我國華南地區(qū)以及國外的越南(Uwa &Parenti,1988;Kottelat,2001;Hayakawaetal.,2015),另有部分人工養(yǎng)殖種群主要存在于日本、泰國(Termvidchakorn & Magtoon,2012)等若干科研機構。日本人工養(yǎng)殖種群種源來自于中國香港,由日本青鳉魚類學者Uwa Hrioshi在1991年所采集(Uwa,1991),已被廣泛用于發(fā)育生物學、遺傳學、毒理學等方面研究(Hamaguchi & Sakaizumi,1992;Sakaizumietal.,1992;Shinomiyaetal.,2006)。

表2 弓背青鳉不同種群主要性狀比較Table 2 Comparison of the characteristics of Oryzias curvinotus in different populations
特別值得注意的是,和其他青鳉魚類如中華青鳉等相似(陳銀瑞等,1989;陳國柱,2016),弓背青鳉正面臨食蚊魚入侵的嚴重威脅。筆者在對中華青鳉滇池周邊種群的研究中,明確觀察到其模式標本產(chǎn)地中最大局部種群在數(shù)年內由于食蚊魚入侵而導致完全滅絕的過程。Dudgeon(1996)指出香港地區(qū)的弓背青鳉種群由于食蚊魚的入侵而嚴重下降,僅在食蚊魚尚未入侵區(qū)域殘存部分種群。本文在采集弓背青鳉的樣地中也同時采集到了大量的食蚊魚個體,并發(fā)現(xiàn)食蚊魚個體數(shù)量較多區(qū)域的弓背青鳉數(shù)量明顯較少。近代以來,食蚊魚在我國已形成廣泛入侵,弓背青鳉現(xiàn)存種群僅呈點狀分布,當前亟待加強其保護生物學研究并應采取有力的保護措施盡早對現(xiàn)存種群進行有效保護。
陳國柱. 2016. 八線腹鏈蛇食物分析及其對中華青鳉的捕食[J]. 四川動物, 35(4): 588-592.
陳銀瑞, 宇和纮, 褚新洛. 1989. 云南青鳉魚類的分類和分布: 鳉形目: 青鳉科[J]. 動物分類學報, 14(2): 239-246.
賈銀濤, 陳毅峰, 陶捐, 等. 2013. 增江魚類群落特征及其歷史變化[J]. 資源科學, 35(7): 1490-1498.
林小濤, 張潔. 2013. 東江魚類生態(tài)及原色圖譜[M]. 北京: 中國環(huán)境出版社: 91.
潘炯華, 劉成漢, 鄭文彪. 1984. 廣東北江魚類區(qū)系研究[J]. 華南師范大學學報(自然科學版), 1: 27-40.
趙俊, 易祖盛, 周先葉, 等. 2010. 廣州水生動植物本底資源[M]. 北京: 科學出版社: 190.
Asai T, Senou H, Hosoya K. 2011.Oryziassakaizumii, a new ricefish from northern Japan (Teleostei: Adrianichthyidae)[J]. Ichthyological Exploration of Freshwaters, 22(4): 289-299.
Dudgeon D. 1996. Anthropogenic influences on Hong Kong streams[J]. Geojournal, 40(1-2): 53-61.
Hamaguchi S, Sakaizumi M. 1992. Sexually differentiated mechanisms of sterility in interspecific hybrids betweenOryziaslatipesandO.curvinotus[J]. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 263(3): 323-329.
Hayakawa H, Le QD, Kinoshita M,etal. 2015. Genetic similarity of the Hainan medaka populations collected from hyper- and hypo-osmotic environments in northern Vietnam[J]. Ocean Science Journal, 50(2): 231-235.
Harada I. 1943. The freshwater fishes of Hainan[J]. Special Service Agency, Hairan Admjnistration, 2: 1-114.
Herder F, Chapuis S. 2010.Oryziashadiatyae, a new species of ricefish (Atherinomorpha: Beloniformes: Adrianichthyidae) endemic to Lake Masapi, central Sulawesi, Indonesia[J]. The Raffles Bulletin of Zoology, 58(2): 269-280.
Herder F, Hadiaty RK, Nolte AW. 2012. Pelvic-fin brooding in a new species of riverine ricefish (Atherinomorpha: Beloniformes: Adrianichthyidae) from Tana Toraja, central Sulawesi, Indonesia[J]. The Raffles Bulletin of Zoology, 60(2): 467-476.
Kottelat M. 2001. Freshwater fishes of northern Vietnam: a preliminary check-list of the fishes known or expected to occur in northern Vietnam with comments on systematics and nomenclature[C]. Environment and Social Development Unit, East Asia and Pacific Region. The World Bank. Freshwater Fish: 56.
Magtoon W. 2010.Oryziassongkhramensis, a new species of ricefish (Beloniformes; Adrianichthyidae) from northeast Thailand and central Laos[J]. Tropical Natural History, 10(1): 107-129.
Mokodongan DF, Tanaka R, Yamahira K. 2014. A new ricefish of the genusOryzias(Beloniformes, Adrianichthyidae) from Lake Tiu, central Sulawesi, Indonesia[J]. Copeia, 2014(3): 561-567.
Nichols JT, Pope CH. 1927. The fishes of Hainan[J]. Bulletion of the American Museum of Natural History, 54: 321-394.
Oshima M. 1926. Notes on a colletion of fishes from Hainan, obtained by professor S. F. Licht, 1926[J]. Annotationes Zoologicae Japonenses, 11(1): 1-25.
Parenti LR. 2008. A phylogenetic analysis and taxonomic revision of ricefishes,Oryziasand relatives (Beloniformes, Adrianichthyidae)[J]. Zoological Journal of the Linnean Society, 154(3): 494-610.
Parenti LR, Hadiaty RK. 2010. A new, remarkably colorful, small ricefish of the genusOryzias(Beloniformes, Adrianichthyidae) from Sulawesi, Indonesia[J]. Copeia, (2): 268-273.
Parenti LR, Hadiaty RK, Lumbantobing D,etal. 2013. Two new ricefishes of the genusOryzias(Atherinomorpha: Beloniformes: Adrianichthyidae) augment the endemic freshwater fish fauna of southeastern Sulawesi, Indonesia[J]. Copeia, (3): 403-414.
Sakaizumi M, Shimizu Y, Hamaguchi S. 1992. Electrophoretic studies of meiotic segregation in inter- and intraspecific hybrids among east Asian species of the genusOryzias(Pisces: Oryziatidae)[J]. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 264(1): 85-92.
Shinomiya A, Kato M, Yaezawa M,etal. 2006. Interspecific hybridization betweenOryziaslatipesandOryziascurvinotuscauses XY sex reversal[J]. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 305(10): 890-896.
Termvidchakorn A, Magtoon W. 2012. Development and identification of three species of Thai ricefish,Oryzias, in the Mekong Basin[J]. Tropical Natural History, 12(1): 75-88.
Uwa H. 1991. Cytosystematic study of the Hainan medaka,Oryziascurvinotus, from Hong Kong (Teleostei: Oryziidae)[J]. Ichthyological Exploration of Freshwaters, (1): 361-367.
Uwa H, Parenti LR. 1988. Morphometric and meristic variation in ricefishes, genusOryzias: a comparison with cytogenetic data[J]. Japanese Journal of Ichthyology, 35(2): 159-166.
Uwa H, Tanaka K, Formacion MJ. 1982. Karyotype and banding analyses of the Hainan medaka,Oryziascurvinotus(Pisces)[J]. Chromosome Inform Service, (33): 15-17.
Uwa H, Wang RF, Chen YR. 1988. Karyotypes and geographical distribution of ricefishes from Yunnan, southwestern China[J]. Japanese Journal of Ichthyology, 35(3): 332-340.
Wang Z, Long S, Liao J,etal. 2017. Complete mitogenome of Hainan medakaOryziascurvinotus(Teleostei: Beloniformes) and transcriptional differences between male and female liver[J]. Mitochondrial DNA Part B, 2(1): 157-158.
CharacteristicRe-descriptionofRicefishOryziascurvinotusfromGuangdong,China
ZHANG Fangfang, JIANG Haochen, JIN Jinjin, QIU Yuping, CHEN Guozhu*
(National Plateau Wetland Research Center, Southwest Forestry University, Kunming 650224, China)
2017-02-21接受日期2017-08-11
國家自然科學基金青年基金項目(31400477); 西南林業(yè)大學高原濕地科學云南省創(chuàng)新團隊項目(2012HC007)
張方方(1992—), 女, 碩士研究生, 主要從事魚類入侵生態(tài)學及瀕危物種保護生物學研究, E-mail:zhang120815@126.com
*通信作者Corresponding author, E-mail:chenguozhu79@163.com
10.11984/j.issn.1000-7083.20170048
Q959.4
A
1000-7083(2017)05-0564-08
