叢枝菌根真菌對(duì)柳枝稷苗期生長(zhǎng)作用機(jī)制的研究
2017-09-13 05:19:42程亭亭劉曙東孫風(fēng)麗奚亞軍
草地學(xué)報(bào) 2017年5期
徐 鵬, 程亭亭, 張 超, 劉曙東, 孫風(fēng)麗*, 奚亞軍*
(1. 西北農(nóng)林科技大學(xué)農(nóng)學(xué)院, 陜西 楊凌 712100; 2. 西北農(nóng)林科技大學(xué)旱區(qū)作物逆境生物學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室, 陜西 楊凌 712100)
柳枝稷(PanicumvirgatumL)是禾本科(Gramineae)黍?qū)?Panicum)的多年生C4植物,原產(chǎn)于北美大草原[1]。它具有適應(yīng)性廣、產(chǎn)量高、對(duì)環(huán)境友好等優(yōu)點(diǎn),已成為最理想的能源作物之一[2-3]。但播種后柳枝稷出苗較晚,苗期生長(zhǎng)緩慢,導(dǎo)致其與同時(shí)期生長(zhǎng)的雜草在水分、營(yíng)養(yǎng)、光照等競(jìng)爭(zhēng)中處于不利地位[4-5]。如果種植當(dāng)年不能建成較好的植被群體,會(huì)提高柳枝稷的生產(chǎn)成本,降低其經(jīng)濟(jì)價(jià)值[2]。相關(guān)研究發(fā)現(xiàn)柳枝稷苗期生長(zhǎng)緩慢可能與土壤類(lèi)型、雜草競(jìng)爭(zhēng)、種子大小等因素相關(guān)[6]。
共生是植物適應(yīng)環(huán)境的一種重要策略。叢枝菌根真菌(Arbuscularmycorrhizalfungi, AMF)可以與大多數(shù)陸生植物形成互利共生的內(nèi)生性菌根,促進(jìn)植物根系養(yǎng)分的吸收和利用[7]。盆栽試驗(yàn)和大田試驗(yàn)都表明柳枝稷根系也能與真菌共生形成菌根,有助于促進(jìn)柳枝稷對(duì)養(yǎng)分的吸收,提高柳枝稷的生物產(chǎn)量[8-10]。因此菌根共生可能是促進(jìn)柳枝稷苗期生長(zhǎng)的重要手段。
植物苗期的生長(zhǎng)需要多方面的協(xié)作與調(diào)控,其中植物激素起著重要的調(diào)控作用。獨(dú)腳金內(nèi)酯(strigolactones, SLs)是一類(lèi)來(lái)源于類(lèi)胡蘿卜素的萜內(nèi)酯,近來(lái)被認(rèn)定為一類(lèi)新型植物激素,控制植物分枝的發(fā)育[11-12]。同時(shí)他還能促進(jìn)叢枝菌根的分枝[13-14],誘導(dǎo)寄生植物如獨(dú)腳金金(Strigaspp.)和列當(dāng)(Orobanchespp.)種子的萌發(fā)[15]。獨(dú)腳金內(nèi)酯途徑相關(guān)基因的研究取得了一系列進(jìn)展,至今為止,利用突變體及功能分析法從不同植物中克隆得到SLs合成途徑中的4個(gè)基因,包括編碼類(lèi)胡蘿卜素裂解雙加氧酶7的同源基因(MAX3/RMS5/DAD3/D17)、編碼類(lèi)胡蘿卜素裂解雙加氧酶8的同源基因(MAX4/RMS1/DAD1/D10)、一個(gè)編碼含鐵蛋白的基因(D27/AtD27)、一個(gè)編碼細(xì)胞色素 P450 蛋白的基因(MAX1)。外源施加人工合成的SLs類(lèi)似物GR24能夠恢復(fù)這些基因的突變體表型[16]。高粱和玉米與菌根真菌共生后能減輕寄生植物獨(dú)腳金的侵染[17-18],可能是由菌根形成后獨(dú)腳金內(nèi)酯含量的下降所導(dǎo)致[19]。因此獨(dú)腳金內(nèi)酯含量的變化很可能是菌根共生過(guò)程中的重要調(diào)控手段。本文研究了叢枝菌根真菌對(duì)柳枝稷苗期生長(zhǎng)的影響,并通過(guò)測(cè)定生理指標(biāo)及獨(dú)腳金內(nèi)酯的含量研究叢枝菌根真菌影響柳枝稷苗期生長(zhǎng)的機(jī)制。
1 材料與方法
1.1 材料與基質(zhì)
供試材料為來(lái)自于柳枝稷低地型栽培品種‘Alamo’的2個(gè)品系Ma和Mg。所用的接種物為Agrauxine公司(英國(guó))商業(yè)化的菌根真菌混合孢子,由中國(guó)科學(xué)院上海生命科學(xué)研究院植物生理生態(tài)研究所王二濤研究員饋贈(zèng)。試驗(yàn)于2016年在西北農(nóng)林科技大學(xué)農(nóng)學(xué)院的光照培養(yǎng)箱以及人工氣候室進(jìn)行。將珍珠巖與沙子按1∶2的比例混合,用121℃高溫滅菌1 h,平均分成2份。取全新的4×8育苗缽用酒精消毒后晾干,將混合基質(zhì)裝入育苗缽中備用。瓜列當(dāng)種子由西北農(nóng)林科技大學(xué)水土保持研究所馬永清研究員饋贈(zèng)。
1.2 種子處理及試驗(yàn)設(shè)計(jì)
將Ma和Mg種子先用10% NaClO處理2 min,再用雙蒸水清洗5次后,在滅菌的濾紙上發(fā)芽[20]。在30℃培養(yǎng)箱中培養(yǎng)10天后,選取長(zhǎng)勢(shì)一致的幼苗移入育苗缽中,每個(gè)育苗缽加入200個(gè)左右孢子單位的接種物作為實(shí)驗(yàn)組,另一份作為對(duì)照組,每種材料種植32株,每個(gè)處理設(shè)置3個(gè)重復(fù)。放入光培養(yǎng)12 h,溫度28℃,光照強(qiáng) 度20 000 lux;暗培養(yǎng)12 h,溫度24℃的循環(huán)培養(yǎng)箱中生長(zhǎng),隔天澆1次超純水。40天后調(diào)查菌根侵染及柳枝稷的生長(zhǎng)發(fā)育狀況。
1.3 菌根墨水及WGA染色
參照水稻菌根染色的方法,取柳枝稷根系用墨水染色觀察侵染情況[21]。先用10% KOH在95℃水浴鍋中處理6 min,然后流水沖洗3遍。隨后用墨水染料(100 mL水+5 mL派克標(biāo)準(zhǔn)墨水+5 mL冰醋酸)95℃染色3 min。水沖洗4~5次后靜置2~3天,每天換水使其脫色,最后制片并在顯微鏡下觀察。
取柳枝稷根系,用WGA染色的方法觀察侵染情況[22]。根系在50%酒精中處理過(guò)夜;用水沖洗去除酒精,在20% KOH中室溫放置2~3天;用水沖洗去除KOH,在0.1 M HCL中處理1~2小時(shí);用水沖洗去除HCL,用PBS溶液 (每800 mL加入8 g NaCl,0.2 g KCl,1.44 g Na2HPO4,0.24 g KH2PO4)再?zèng)_洗2遍;用水沖洗后,加入WGA-Alexafluor 488染液,室溫處理6 h左右,制片后在熒光顯微鏡 (激發(fā)波長(zhǎng)488 nm)下觀察。
1.4 生理指標(biāo)的測(cè)定
將幼苗整株取出,用自來(lái)水將根系仔細(xì)地沖洗干凈,然后用吸水紙將水吸干,分別測(cè)定各組的鮮重及干重、根長(zhǎng)、株高、葉綠素含量。
根長(zhǎng):測(cè)定10株幼苗單株的根長(zhǎng),取其平均值。
鮮重、干重:取10株幼苗測(cè)定其鮮重,于105℃下殺青15 min,85℃下烘干至恒重,測(cè)定干重。
葉綠素含量:利用葉綠素測(cè)定儀(SPAD-502Plus)直接測(cè)定葉片中葉綠素含量。
1.5 根系分泌物的提取及瓜列當(dāng)萌發(fā)處理
將幼苗根系清洗干凈后進(jìn)行風(fēng)干處理,研缽研成粉末。取100 mg粉末加入1.5 mL甲醇,超聲30 min后離心取上清作為原液,稀釋10倍備用。參照安雨[23]的方法處理瓜列當(dāng)種子,分別以GR24和去離子水處理為正、負(fù)對(duì)照。
1.6 RNA提取,cDNA合成及熒光定量PCR
取處理后的柳枝稷根系液氮速凍,用Trizol法抽提RNA,然后參照天根反轉(zhuǎn)試劑盒去除gDNA以及反轉(zhuǎn)錄合成cDNA。使用柳枝稷中SLs合成相關(guān)基因的定量引物(表1)在伯樂(lè)公司的CFX96型實(shí)時(shí)熒光定量?jī)x中進(jìn)行擴(kuò)增。
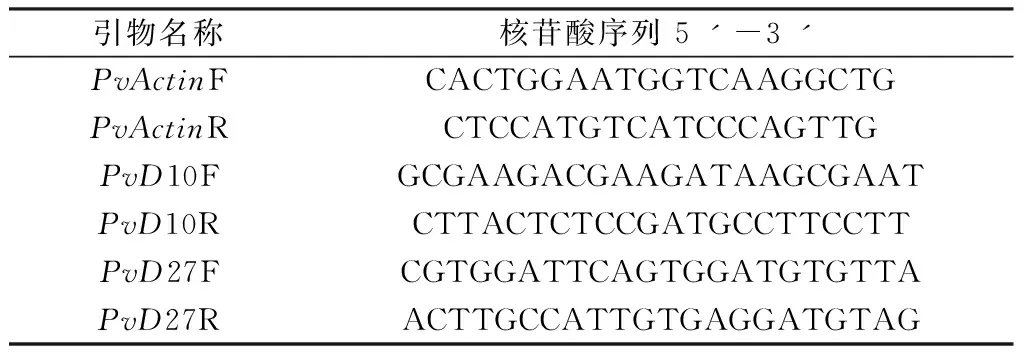
表1 SLs合成相關(guān)基因及內(nèi)參基因定量引物序列Table 1 The primers of gene in SLs synthesis pathway and internal reference gene actin
1.7 數(shù)據(jù)分析
利用Excel 2010做圖表,利用LDS法對(duì)各指標(biāo)進(jìn)行方差分析。
2 結(jié)果與分析
2.1 叢枝菌根真菌侵染的檢測(cè)
叢枝菌根是內(nèi)生菌根的主要類(lèi)型之一,其菌絲沒(méi)有橫膈膜,可在根的內(nèi)皮層細(xì)胞內(nèi)形成叢枝狀結(jié)構(gòu),其中有些還能在細(xì)胞內(nèi)或皮層細(xì)胞間形成泡囊結(jié)構(gòu)。墨水染色(圖1)和WGA染色(圖2)均能明顯觀察到含有叢枝狀菌絲結(jié)構(gòu)和充滿泡囊的細(xì)胞,說(shuō)明菌根真菌侵染成功。對(duì)照中則沒(méi)有觀察到菌根真菌侵染的跡象。
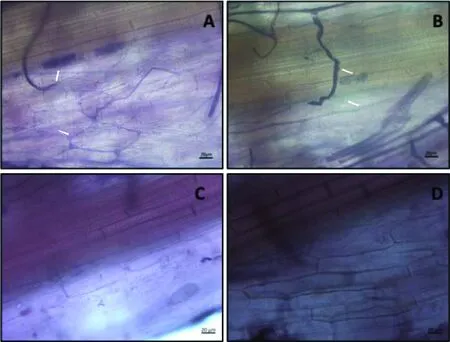
圖1 柳枝稷菌根侵染墨水染色(A. Ma接菌;B. Mg接菌;C. Ma對(duì)照;D. Mg對(duì)照)Fig.1 Staining of AMF in switchgrass roots with ink ( A. Ma-AMF; B. Mg-AMF; C. Ma-CK; D. Mg-CK)
圖2 柳枝稷菌根侵染W(wǎng)GA染色(A-B. Ma接菌;C-D. Ma對(duì)照;E-F. Mg接菌;G-H. Mg對(duì)照)Fig.2 Staining of AMF in switchgrass roots with WGA( A and B. Ma-AMF; C and D. Ma-CK; E and F. Mg-AMF; G and H. Mg-CK)
2.2 叢枝菌根真菌侵染對(duì)柳枝稷苗期表型的影響
由表2可知,Ma和Mg 2個(gè)品系接種菌根真菌后,除了根長(zhǎng)沒(méi)有變化外,株高、干重、鮮重、葉綠素含量和主莖葉片數(shù)等均極顯著高于對(duì)照(P<0.01),這說(shuō)明菌根真菌與苗期柳枝稷共生后,通過(guò)促進(jìn)柳枝稷根部養(yǎng)分吸收,加快柳枝稷地上葉片的生長(zhǎng),最終增加柳枝稷的生物量。菌根共生對(duì)柳枝稷苗期的生長(zhǎng)具有促進(jìn)作用。
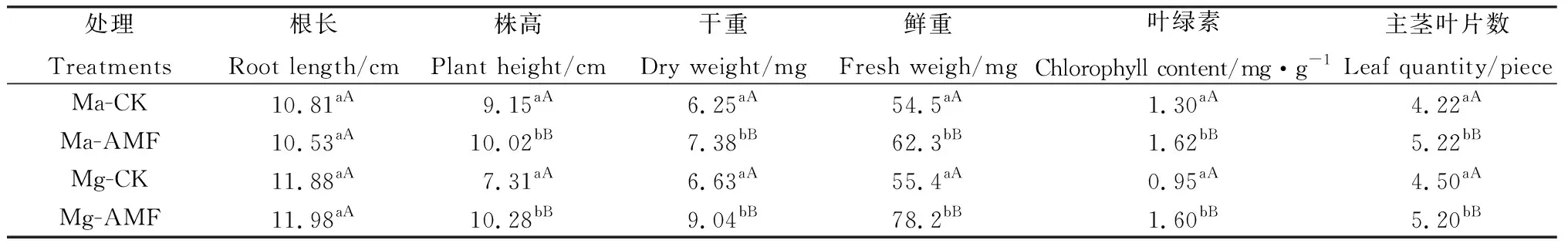
表2 菌根真菌侵染對(duì)柳枝稷苗期生長(zhǎng)指標(biāo)的影響Table 2 Effects of Arbuscular Mycorrhizal Fungi on the growth of switchgrass at seedling stage
注:同列不同小寫(xiě)字母表示不同處理間差異顯著(P<0.05),不同大寫(xiě)字母表示差異極顯著(P<0.01)
Note: Different lowercase letters in the same column indicate significant differences at the 0.05 level among different treatments; different capital letters indicate significant differences at the 0.01 level
2.3 叢枝菌根真菌侵染對(duì)根系獨(dú)腳金內(nèi)酯含量的影響
寄主植物根系分泌的獨(dú)腳金內(nèi)酯能夠促進(jìn)獨(dú)腳金、列當(dāng)?shù)燃纳参锏拿劝l(fā)[15],因此我們利用柳枝稷根系粉末的甲醇提取液處理寄生植物瓜列當(dāng)?shù)姆N子,其發(fā)芽率作為判斷根系獨(dú)腳金內(nèi)酯含量的間接指標(biāo)。作為陽(yáng)性對(duì)照的GR24處理,瓜列當(dāng)種子萌發(fā)率達(dá)到了37.7%,而陰性對(duì)照H2O處理,瓜列當(dāng)種子萌發(fā)率只有2.8%。比較Ma和Mg 2個(gè)品系的處理發(fā)現(xiàn),Ma和Mg未接菌處理的根系提取物處理,瓜列當(dāng)?shù)拿劝l(fā)率分別達(dá)到了22.6%和57.3%,而接菌處理的根系提取物處理后其萌發(fā)率分別只有9.6%和19.8%(圖3)。由此可以初步推測(cè),接種菌根真菌后,根系中獨(dú)腳金內(nèi)酯含量會(huì)顯著下降,從而導(dǎo)致瓜列當(dāng)?shù)拿劝l(fā)率只有未接菌的1/3。為進(jìn)一步驗(yàn)證獨(dú)腳金內(nèi)酯的變化,我們選擇了Mg接菌和未接菌的根系材料提取RNA,反轉(zhuǎn)cDNA后利用實(shí)時(shí)熒光定量的方式檢測(cè)SLs合成相關(guān)標(biāo)記基因的表達(dá)情況,利用柳枝稷的Actin基因作為內(nèi)參基因,結(jié)果發(fā)現(xiàn)在Mg未接菌材料中,SLs合成相關(guān)標(biāo)記基因D10和D27的表達(dá)量顯著高于接菌的Mg(圖4)。綜合以上結(jié)果,接種叢枝菌根真菌后柳枝稷根系中獨(dú)腳金內(nèi)酯含量會(huì)明顯減少。
3 討論
柳枝稷作為新型能源作物而倍受關(guān)注,但因其苗期生長(zhǎng)緩慢,植株建植時(shí)間長(zhǎng),降低了柳枝稷的利用價(jià)值,加大了柳枝稷推廣的難度。共生是植物適應(yīng)環(huán)境的一種重要策略,叢枝菌根真菌能與大多數(shù)陸地上的維管植物形成專(zhuān)性共生體系,促進(jìn)根部營(yíng)養(yǎng)吸收以及植物生長(zhǎng)發(fā)育并增強(qiáng)植物的環(huán)境適應(yīng)能力。因此本試驗(yàn)利用來(lái)自于柳枝稷‘Alamo’的Ma和Mg 2個(gè)品系為試驗(yàn)材料,接種叢枝菌根真菌的混合孢子,研究菌根共生對(duì)柳枝稷苗期生長(zhǎng)的影響。結(jié)果發(fā)現(xiàn)菌根真菌能成功侵染柳枝稷根系。接菌后的柳枝稷株高增加,葉片數(shù)增多,生物量顯著增長(zhǎng),說(shuō)明菌根真菌與植株共生后能加快柳枝稷苗期的生長(zhǎng)發(fā)育。因此,在種植柳枝稷時(shí)可通過(guò)施撒菌根真菌孢子的方法促進(jìn)柳枝稷苗期的生長(zhǎng),加快其建植速度。
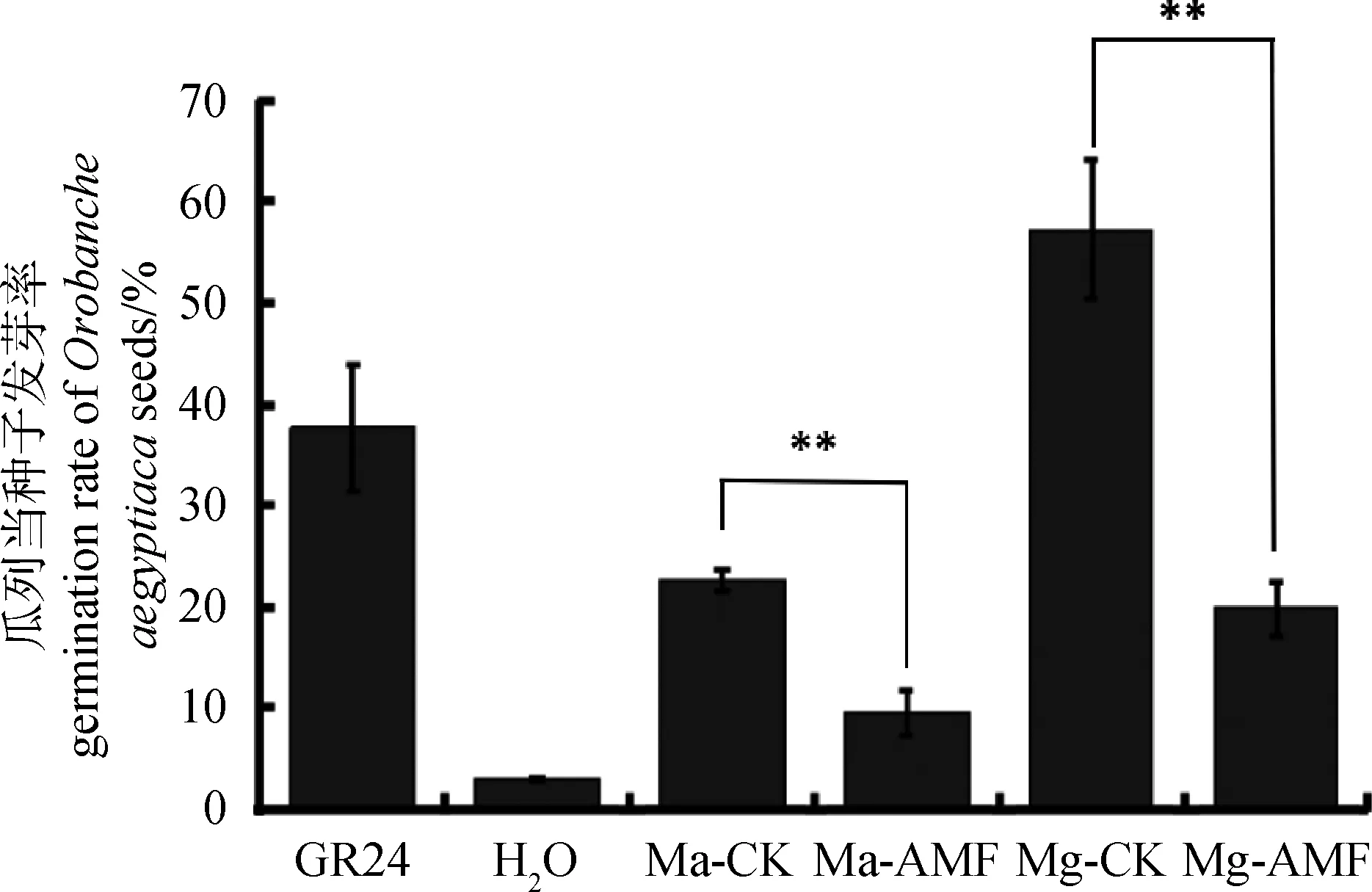
圖3 柳枝稷根系甲醇浸提液處理瓜列當(dāng)?shù)陌l(fā)芽率Fig.3 Germination rate of Orobanche aegyptiacainduced with extracts from switchgrass roots
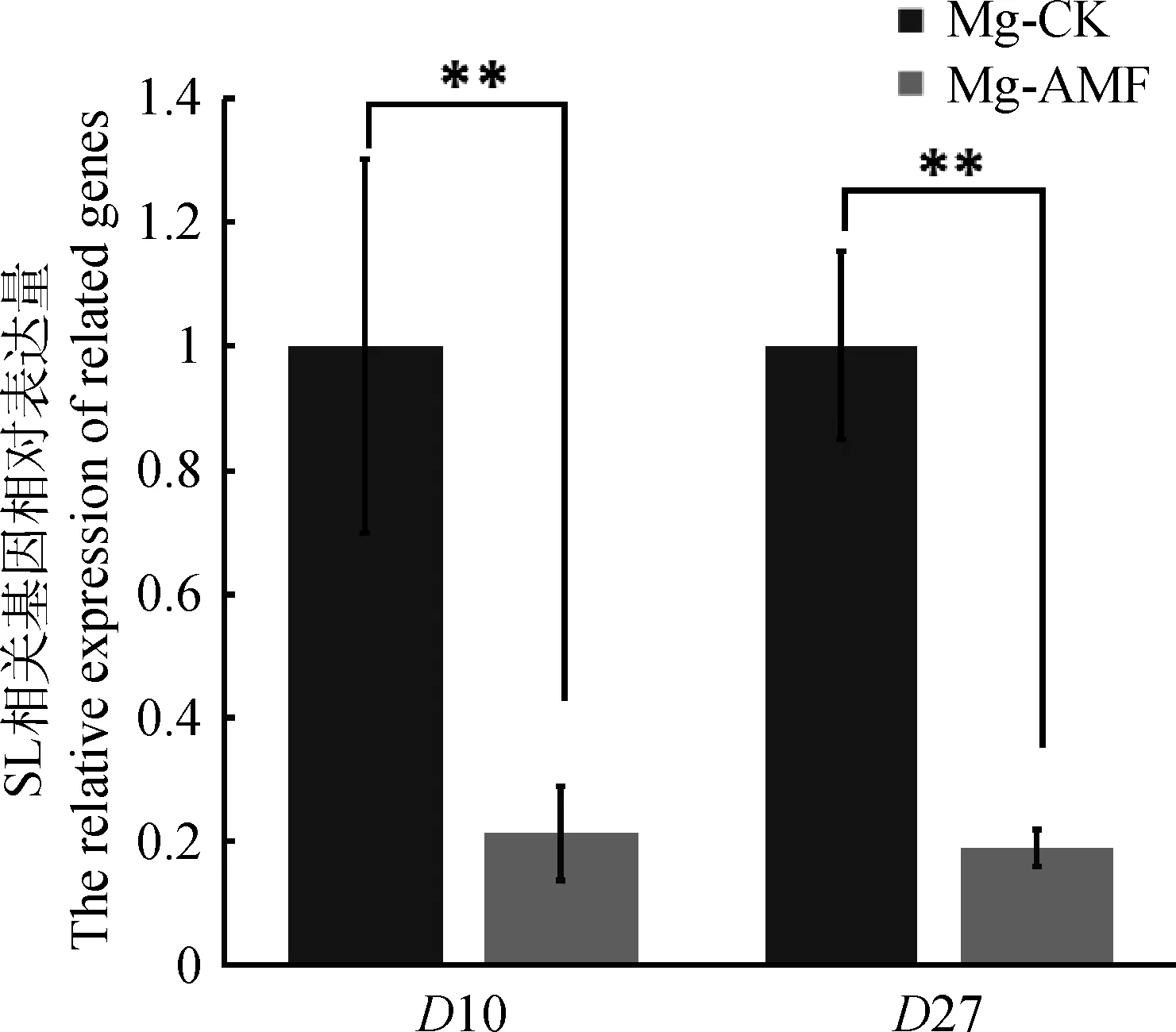
圖4 柳枝稷中SLs合成基因表達(dá)量檢測(cè)Fig.4 Expression of genes involved in SLs synthesis in switchgrass
菌根真菌促進(jìn)柳枝稷苗期生長(zhǎng)的過(guò)程中,獨(dú)腳金內(nèi)酯很可能發(fā)揮著重要的調(diào)控作用。植物與菌根真菌的共生起始于信號(hào)交換,寄主植物根系可能通過(guò)分泌一種“分枝因子”(branching factors,BFs)來(lái)誘導(dǎo)叢枝真菌菌絲大量分枝,從而增加菌絲與寄主植物根系接觸的機(jī)會(huì)[24]。而真菌自身也會(huì)釋放一種可擴(kuò)散的“Myc因子”(myc factor,MF)誘導(dǎo)寄主植物根系相關(guān)基因的表達(dá),從而為菌根真菌的侵染提供有利條件[25]。目前BFs已被證實(shí)是SLs[13],而MFs的成分目前仍不清楚。寄主植物根系分泌的SLs可以通過(guò)誘導(dǎo)一系列的分子和細(xì)胞生物學(xué)事件,促進(jìn)叢枝菌絲形成大量分枝。Ma和Mg接種菌根真菌后,其葉綠素含量、生物量、株高與對(duì)照相比都有了顯著升高,說(shuō)明當(dāng)植物與真菌的共生關(guān)系形成后,植物的營(yíng)養(yǎng)狀況會(huì)因?yàn)楣采婢淖饔枚玫礁纳啤T谶@種情況下,我們用根系的甲醇提取液處理寄生植物瓜列當(dāng),測(cè)定其發(fā)芽率,發(fā)現(xiàn)瓜列當(dāng)在菌根共生的柳枝稷的提取液處理后,發(fā)芽率顯著低于未接菌柳枝稷的處理,而SLs的重要作用就是促進(jìn)寄生植物的萌發(fā)。進(jìn)一步檢測(cè)Mg根系中SLs合成標(biāo)記基因的表達(dá)水平,發(fā)現(xiàn)未接菌的Mg根系中D10和D27的表達(dá)量顯著高于接菌的Mg。這就說(shuō)明柳枝稷在共生后會(huì)減少獨(dú)角金內(nèi)酯的合成,促進(jìn)植物分枝的形成,對(duì)植物的生長(zhǎng)有促進(jìn)作用。相反,未接菌根真菌的柳枝稷由于得不到充分的無(wú)機(jī)營(yíng)養(yǎng),會(huì)加速獨(dú)腳金內(nèi)酯的分泌,從而誘導(dǎo)土壤中的菌根真菌孢子萌發(fā)和促進(jìn)分枝,以期盡快形成促進(jìn)營(yíng)養(yǎng)吸收的菌根結(jié)構(gòu)。
4 結(jié)論
以來(lái)自于低地型品種Alamo的2個(gè)品系Ma和Mg的柳枝稷為材料,研究叢枝菌根真菌對(duì)柳枝稷苗期生長(zhǎng)發(fā)育的影響,研究發(fā)現(xiàn)叢枝菌根真菌能夠成功侵入苗期柳枝稷的根內(nèi),形成有功能的共生菌根,并且能顯著增加柳枝稷苗期的株高、葉綠素含量、葉片數(shù)和生物量。提取根系分泌物處理寄生植物瓜列當(dāng)?shù)姆N子,發(fā)現(xiàn)接菌后柳枝稷的根系分泌物會(huì)顯著降低瓜列當(dāng)?shù)陌l(fā)芽率,同時(shí)觀察到Mg接菌后植物體內(nèi)獨(dú)腳金內(nèi)酯合成關(guān)鍵基因D10和D27表達(dá)量顯著下調(diào),提示柳枝稷形成菌根后會(huì)抑制根系獨(dú)腳金內(nèi)酯的合成和分泌,從而解除獨(dú)腳金內(nèi)酯對(duì)柳枝稷分蘗的抑制作用,促進(jìn)柳枝稷苗期的生長(zhǎng)。
