混播比例和澆水頻率對熱研2號柱花草和巴西含羞草生長早期種間競爭的影響
2017-09-13 05:19:42李雪楓趙慶杰
草地學報 2017年5期
李雪楓, 趙慶杰, 王 堅*
(1. 海南大學熱帶農林學院, 海南 海口570228; 2. 海南大學應用科技學院, 海南 儋州571737)
外來入侵植物不但嚴重影響侵入地的生物多樣性和生態環境,還威脅著全球的生態環境和經濟發展[1-2]。得天獨厚的地理位置和水熱資源條件使海南省成為熱帶、亞熱帶植物生長繁衍的“天堂”,也使其成為易遭受外來生物入侵的區域[3]。巴西含羞草(MimosainvisaMart. ex Colla)[4],又名美洲含羞草(M.diplotrichaC. Wright ex Sauvalle)[5],豆科(Leguminosae)含羞草屬(MimosaL.)一年生或多年生亞灌木狀草本植物,原產于巴西[6],生長力頑強,對土壤要求不高,現已廣泛侵入亞洲地區[6-7]、非洲地區[8-10]及一些太平洋島國[11-12],也是入侵海南島的惡性雜草之一[3-5]。巴西含羞草五棱柱狀的莖棱上密生堅硬鉤刺,動物采食、刈割和砍伐困難[12],還會攀爬于其他植物上致使其窒息死亡[8];其花果期通常4~6 月,無寒冷天氣時4~12 月[12],莢果的邊緣及莢節有刺毛,可粘附在動物皮毛、人類衣物、農作機具和各種交通工具上傳播,侵入休閑地、住宅區、路邊、牧場及各種種植園區[7,13],不但影響當地的植物多樣性,還會降低作物產量、增加養殖成本、增大作物損失和土壤退化程度[8,12];此外,含羞草屬植物各部分都有毒,如果攝入,產生的毒素Mimosin(一種非蛋白質氨基酸)可引起牛血管內皮損傷、心臟和肝臟壞死及貧血等病癥[12]。因此,巴西含羞草是侵入區一個重要的經濟、農業和生態負擔,控制巴西含羞草在海南滋生蔓延將成為我國熱帶和亞熱帶地區農業經濟發展和生態安全的重要內容。
替代控制是進行入侵植物防控的一種重要的生態防治方法,指根據植物種間競爭規律和群落演替規律,利用有經濟價值或有生態價值的植物通過占位覆蓋的方法來排擠控制入侵植物的擴張和蔓延,達到防除入侵植物的目的,即“以草制草”。應用替代控制技術,不僅可以避開化學防治和引入天敵的風險,在替代植物定植成功后還能長期控制入侵植物,保持水土、改良土壤、提高環境質量,具有生態、經濟和可持續的特點[14],目前已在紫莖澤蘭(Ageratinaadenophora)[15]、黃頂菊(Flaveriabidentis)[16]和飛機草(Eupatoriumodoratum)[17]等外來入侵雜草的防控方面進行了相關的試驗研究,并取得較好的成果,但有關巴西含羞草替代防治的研究工作目前鮮有報道。
熱研2號柱花草(Stylosanthesguianensis‘Reyan No. 2’),豆科柱花草屬(S. Sw. ex Willd)多年生草本植物,是中國熱帶農業科學院熱帶牧草研究中心選育,于1991年經牧草品種審定委員會審定登記的牧草品種。該品種莖葉產量高,營養豐富,適應性強,從砂質土到重粘土均可良好生長,耐干旱,耐酸性瘦土,現已成為我國熱帶、南亞熱帶地區的當家豆科牧草品種,在海南、廣東、廣西、福建等省及云南和四川的熱帶地區大面積推廣[18]。根據替代植物必須具有速生、郁閉度高、利用價值好的特性,本研究以熱研2號柱花草為研究對象,旨在通過對熱研2號柱花草與巴西含羞草的苗期競爭效應的研究,探討熱研2號柱花草替代控制巴西含羞草的最佳栽培管理模式,為巴西含羞草的生態控制及其入侵地修復提供必要參考。
1 材料與方法
1.1 試驗材料
熱研2號柱花草種子:由中國熱帶農業科學院熱帶牧草研究中心提供。
巴西含羞草種子:采自海南大學海甸校區周邊,成熟飽滿的種子在背光通風處自然風干后,紙袋貯存備用。
土壤:試驗用土壤取自海南省澄邁縣,土壤為玄武巖母質發育的磚紅壤,采集表層0~30 cm土壤,風干后,過1 mm篩備用。土壤基礎性狀:pH 5.03、土壤有機質14.96 g·kg-1、全氮1.32 g·kg-1、全磷0.91 g·kg-1、全鉀3.95 g·kg-1、堿解氮73.49 mg·kg-1、速效磷17.70 mg·kg-1、速效鉀106.8 mg·kg-1,鹽基飽和度15.2%,土壤質地為重粘土。
營養土:椰糠、有機肥、泥炭和生物肥的混合物發酵而成,pH 6.5~7.0,海南源源園藝有限公司生產。
1.2 試驗方法
1.2.1盆栽準備及種子預處理 在海南大學環植學院實驗基地大棚內,將風干后的土壤篩除石塊、草根等雜質,以土壤:營養土=4:1的比例均勻混合,裝至花盆(上徑33 cm,下徑25 cm,高25 cm)上口徑邊沿2 cm處,備用。為保障種子同期萌發,巴西含羞草種子播種前參考含羞草(M.pudica)種子處理方法, 95℃的熱水浴處理1 min[19];熱研2號柱花草種子播種前用80℃熱水浸種3~5 min[18],然后將兩種種子分別置于育苗盤育苗,幼苗長至2~3葉期待用。
1.2.2試驗設計 參考丁文利[20]等的生態替代法,每個花盆均勻設8 穴,按照每個花盆內熱研2號柱花草:巴西含羞草(簡稱柱:巴)為 8:0,6:2,4:4,2:6,0:8共5 個混播比例,每穴移栽大小一致、長勢基本相同的2~3 葉期幼苗1 株,每盆共8 株,每個混播比例種12 盆。根據同一花盆中同種植物株數占總株數的比例(簡稱占比),將熱研2號柱花草和巴西含羞草的占比記為8/8、6/8、4/8和2/8。然后將每種混播比例均設置1次·1d-1﹑1次·3d-1﹑1次·5d-1和1次·7d-1共4 個澆水處理,每次澆水量為試驗用土壤最大持水量的70%,每個處理3 個重復。整個試驗期間不施肥,花盆隨機挪動,以消除光照、邊際等影響。試驗期間大棚內最高溫度38℃,最低溫度27℃,平均溫度33℃,空氣濕度75%。
1.2.3指標測定 試驗期間,對每株植物掛牌標記,每5 d量測1 次每株植物的伸直高度。第30 d,考慮巴西含羞草受擾動小葉合攏的特性,首先測量每株植物的伸直高度,對每株植物的復葉總數進行計數,再按照統一標準分別在每株植物的上部、中部和下部分別取代表性復葉各6 片,分別稱重后用方格法測定植物葉面積,并計算葉面積指數;然后將植株從花盆取出,小心抖落根系土壤,再用清水清洗干凈,先將每株植物的地上與地下部分分開,再將地上部分的莖和葉分開(其中葉柄和小葉柄均計入葉),與根系分別置于105℃烘箱殺青30 min,然后75℃烘干,稱取各部分干重。
1.3數據分析 由于設置了混播比例和澆水頻率兩個影響因子,在該試驗無法使用目前比較種間競爭常用的各指標相對值[15,21],否則將出現單播條件下不同澆水頻率處理時各指標相對值均為1的情況,故直接應用各指標絕對值進行比較。采用SPSS 18.0對柱花草和巴西含羞草的生物量指標進行一般線性模型(GLM)單變量分析;然后對同一因素處理下各指標間的差異進行單因素方差分析,其中方差齊性時用Tukey法在P<0.05水平進行顯著性檢驗,方差齊性檢驗不適合時用Dunnett’s C法在P<0.05水平進行顯著性檢驗。所有數值均表示為平均值±標準差;Microsoft Office Excel 2003作圖。
2 結果與分析
2.1 混播比例和澆水頻率對植物單株總生物量的影響
混播比例、澆水頻率和這兩種處理的交互作用對熱研2號柱花草和巴西含羞草生長早期(30 d)的單株總生物量(干重)均有顯著影響(表1)。同一混播占比下,隨澆水間隔期延長,柱花草和巴西含羞草的單株總生物量均呈先增后降的趨勢(圖1),其中柱花草在1次·3d-1澆水時單株總生物量均顯著大于其他澆水處理,1次·1d-1次之,1次·5d-1和1次·7d-1間差異不顯著(圖1Ⅰ);而巴西含羞草在1次·5d-1澆水時單株總生物量最大,1次·1d-1最小,1次·3d-1和1次·7d-1間無顯著差異。同一澆水頻率下,隨柱花草所占比例減少(即混播株數減少),單株總生物量均顯著降低(P<0.05),其中降幅最顯著為1次·7d-1澆水處理,1次·5d-1次之,1次·3d-1最小(圖1Ⅰ);同一澆水頻率下,隨巴西含羞草所占比例減少,其單株總生物量逐漸增大,其中1次·5d-1增幅最顯著,1次·1d-1增幅最小,1次·3d-1和1次·7d-1增幅間無顯著差異(圖1Ⅱ)。
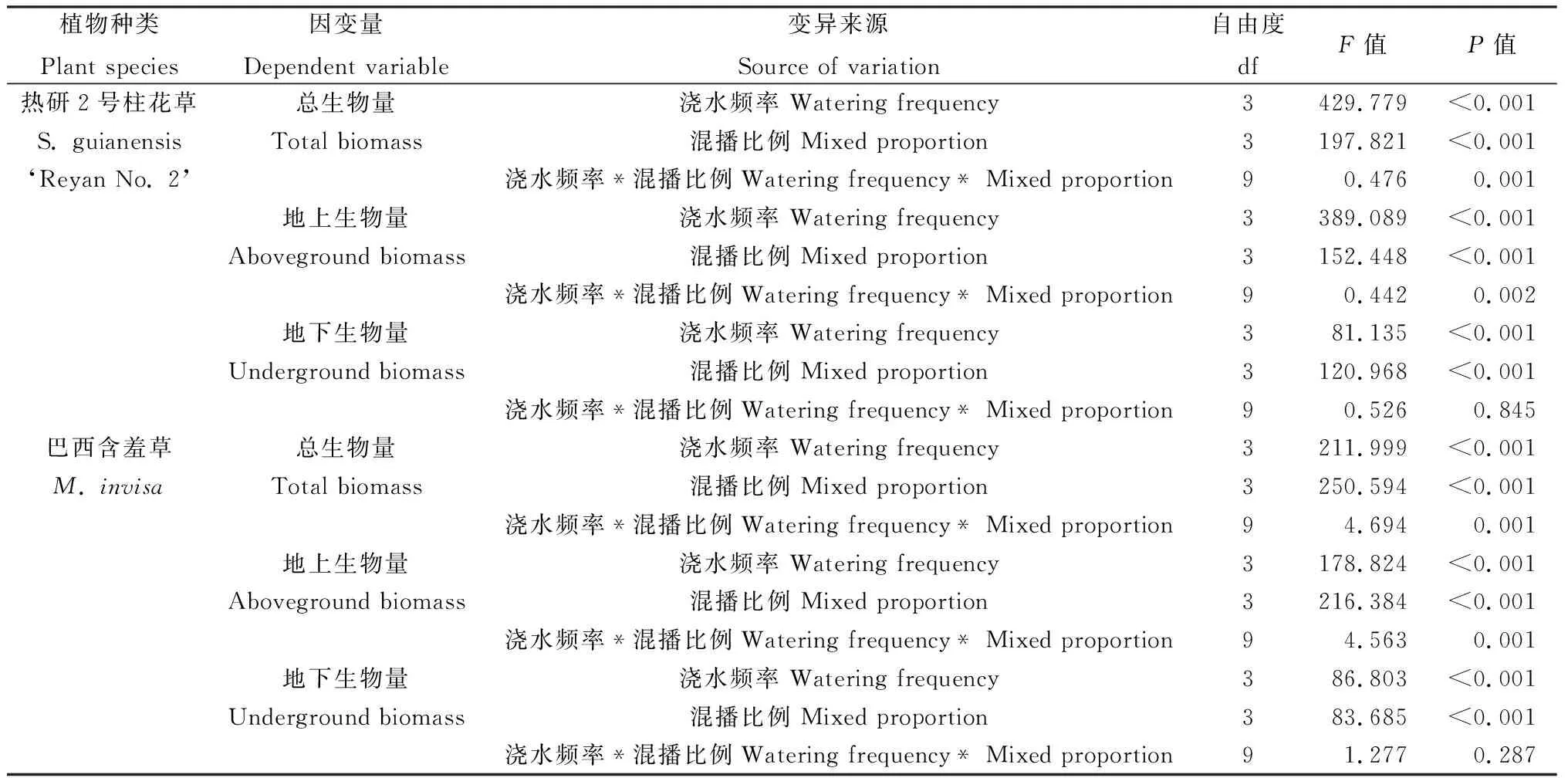
表1 熱研2號柱花草和巴西含羞草生物量的單變量分析Table 1 Univariate analysis of general linear model for biomass of S. guianensis ‘Reyan No. 2’ and M. invisa

圖1 混播比例、澆水頻率對熱研2號柱花草(Ⅰ)和巴西含羞草(Ⅱ)單株總生物量的影響Fig.1 Effects of the mixed sowing proportion and watering frequency on the total biomass per plant ofS. guianensis ‘Reyan No. 2’ (Ⅰ) and M. invisa (Ⅱ) 注:不同小寫字母表示同一混播比例下受試植物在不同澆水頻率處理間差異顯著性,不同大寫字母表示同一澆水頻率下受試植物在不同混播比例時的差異顯著性(P<0.05),下同Note: Different lowercase letters indicate significant difference among the same proportion of the test plants under differentwatering frequencies at the 0.05 level; different capital letters indicate significant difference among the same watering frequency of thetest plants under different proportions at the 0.05 level. The same below
2.2 混播比例和澆水頻率對植物根冠比的影響
1次·1d-1澆水時,柱花草各占比間根冠比無顯著差異,其他3種澆水處理根冠比均隨占比降低均逐漸減小,且僅6/8與2/8占比之間差異顯著(P<0.05)(表2);1次·3d-1澆水時,巴西含羞草各占比間根冠比無顯著差異,其他澆水頻率時隨混播占比的減小略有升高,但僅8/8與2/8占比間差異顯著(P<0.05)。在占比8/8時,不同澆水頻率間柱花草的根冠比無顯著差異,其他占比下,1次·3d-1的澆水頻率下根冠比顯著小于其他澆水頻率(P<0.05);同一混播占比下,巴西含羞草的根冠比隨澆水間隔期的延長逐漸增大(表2)。
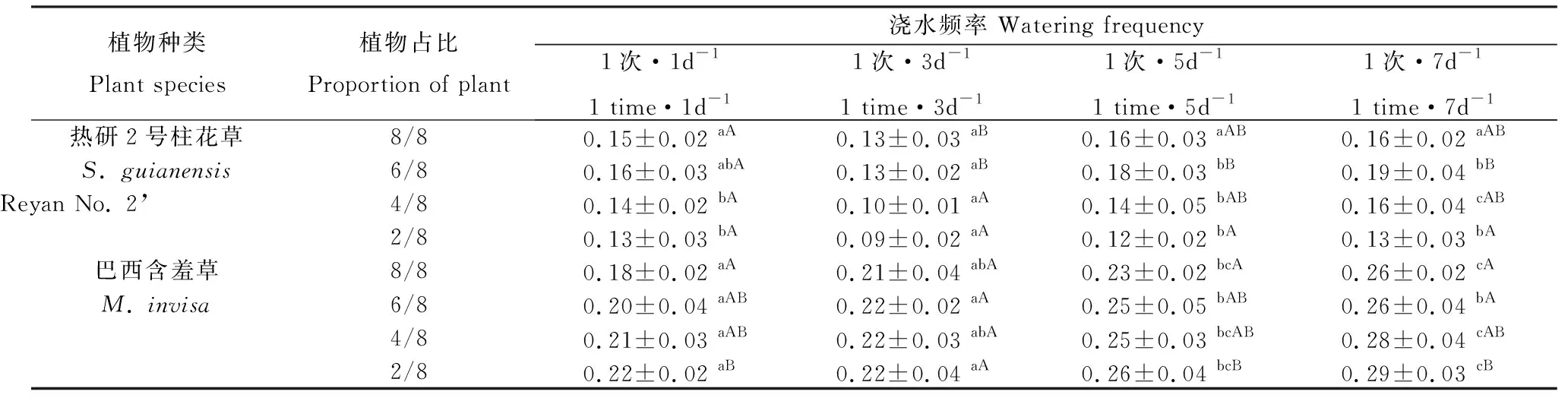
表2 澆水頻率、混播比例對熱研2號柱花草和巴西含羞草根冠比的影響Table 2 Effects of the mixed sowing proportion and watering frequency on root/shoot ratio of S. guianensis ‘Reyan No. 2’ and M. invisa
注:同行不同小寫字母表示同一混播比例下各受試植物在不同澆水頻率間差異顯著(P<0.05);同列不同大寫字母表示同一澆水頻率下各受試植物在不同混種比例間差異顯著(P<0.05),下同
Note: Different lowercase letters in the same row indicate significant difference under different watering frequencies among the same proportion of the test plants at the 0.05 level; different capital letters in the same column indicate significant difference under different proportions among the same watering frequency of the test plants at the 0.05 level. The same below
2.3 混播比例和澆水頻率對植物生物量分配的影響
混播比例、澆水頻率對熱研2號柱花草和巴西含羞草生長早期(30 d)地上生物量(干重)有顯著的交互作用,對地下生物量無顯著交互作用(表1)。生長早期,各處理下柱花草和巴西含羞草的生物量均優先分配給葉,莖次之,根系最少(圖2)。同一占比在不同澆水頻率下,隨澆水間隔期延長,柱花草和巴西含羞草的根生物量、莖生物量和葉生物量均呈先增后降趨勢(圖2),其中,柱花草在澆水處理1次·1d-1與1次·3d-1間、1次·5d-1與1次·7d-1間其根生物量均無顯著差異(圖2Ⅰ-R),莖和葉生物量在1次·5d-1與1次7d-1間均無顯著差異,其他處理間均差異顯著(圖2Ⅰ-S,L);僅在占比為8/8時,1次·3d-1和1次·7d-1的澆水處理間巴西含羞草的葉生物量差異顯著(P<0.05),其他占比下均無顯著差異,各占比下巴西含羞草莖生物量的變化趨勢同根生物量相似,1次·3d-1和1次·7d-1澆水處理間無顯著差異(圖2Ⅱ-R,S,L)。同一澆水頻率下,柱花草根、莖、葉生物量均在占比8/8時最大(圖2Ⅰ),隨占比減少,各部分生物量均逐漸降低(圖2Ⅰ),且1次·3d-1的降幅最小。同一澆水頻率下,巴西含羞草各部分生物量隨占比減少而逐漸增大,1次·5d-1澆水處理增幅最大(圖2Ⅱ)。
2.4 混播比例和澆水頻率對植物株高的影響
在同一占比下,隨澆水間隔期延長,柱花草和巴西含羞草株高均呈先增后降趨勢,其中1次·3d-1澆水時各占比均最大,1次·7d-1時均最小;巴西含羞草的株高在1次·5d-1時各占比均最大,1次·3d-1次之,1次·7d-1最小(表3)。同一澆水頻率,柱花草的株高隨占比減少逐漸降低,占比8/8時的株高均顯著大于其他占比的(P<0.05), 4/8與6/8占比間株高無顯著差異;同一澆水頻率,巴西含羞草株高隨占比增大而逐漸增大, 2/8與4/8占比間的株高無顯著差異,但2/8與8/8和6/8占比間的株高有顯著差異(P<0.05)(表3)。

表3 澆水頻率、混播比例對熱研2號柱花草和巴西含羞草株高的影響Table 3 Effects of the mixed sowing proportion and watering frequency on the height of S. guianensis‘Reyan No. 2’ and M. invisa /cm
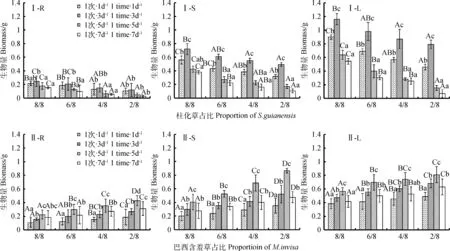
圖2 混播比例和澆水頻率對熱研2號柱花草(Ⅰ)和巴西含羞草(Ⅱ)生物量分配的影響Fig.2 Effects of the mixed sowing proportion and watering frequency on the biomass allocationof S. guianensis ‘Reyan No. 2’(Ⅰ)and M. invisa(Ⅱ)注:字母R表示受試植物的根系;S表示受試植物的莖;L表示受試植物的葉Note: the letter “R”, “S” and “L” refer to the root, stem and leaf biomass of the test plants
2.5 混播比例和澆水頻率對植物生長速度的影響
同一占比下,柱花草的早期平均生長速度均在1次·3d-1澆水時最大,1次·1d-1次之,1次·7d-1最小(圖3Ⅰ),巴西含羞草則在1次·5d-1澆水時最大,1次·3d-1次之,1次·7d-1最小(圖3Ⅱ)。1次·1d-1、1次·3d-1和1次·5d-1澆水處理下,各占比柱花草生長速度峰值均在6-10 d時出現,1次·7d-1澆水處理推遲至11-15 d時出現;占比8/8時各澆水處理下柱花草生長速度的峰值均顯著大于其他占比對應澆水處理時的對應值(P<0.05),隨占比減小,各澆水處理下柱花草平均生長速度的峰值逐漸降低(圖3Ⅰ)。同一澆水頻率下,巴西含羞草各階段的生長速度隨占比減小而逐漸增大;1次·3d-1和1次·5d-1澆水時生長速度峰值均出現于11~15 d間,最大值分別為1.18 cm·d-1和1.34 cm·d-1(圖3Ⅱ);而1次·1d-1和1次·7d-1澆水時生長速度峰值出現于6~10 d,最大值分別為0.86 cm·d-1和0.91 cm·d-1(圖3Ⅱ)。

圖3 混播比例和澆水頻率對熱研2號柱花草(Ⅰ)和巴西含羞草(Ⅱ)生長速度的影響Fig.3 Effects of the mixed sowing proportion and watering frequency on thegrowth rate of S. guianensis ‘Reyan No. 2’(Ⅰ)and M. invisa(Ⅱ)
2.6 混播比例和澆水頻率對植物葉面積指數的影響
同一澆水頻率下,柱花草葉面積指數隨占比減少逐漸減小,而巴西含羞草葉面積指數隨占比減少逐漸增大(表4),且2種植物的葉面積指數在占比為4/8與6/8間均無顯著差異。同一占比下,隨澆水間隔期延長,柱花草和巴西含羞草的葉面積指數均先增后降,其中柱花草1次·3d-1澆水時的葉面積指數顯著大于其他澆水處理的,1次·1d-1和1次·5d-1的葉面積指數間無顯著差異,1次·7d-1澆水時葉面積指數最小;巴西含羞草1次·5d-1澆水時的葉面積指數顯著大于其他澆水處理時的(P<0.05),1次·1d-1和1次·7d-1處理間的葉面積指數無顯著差異。
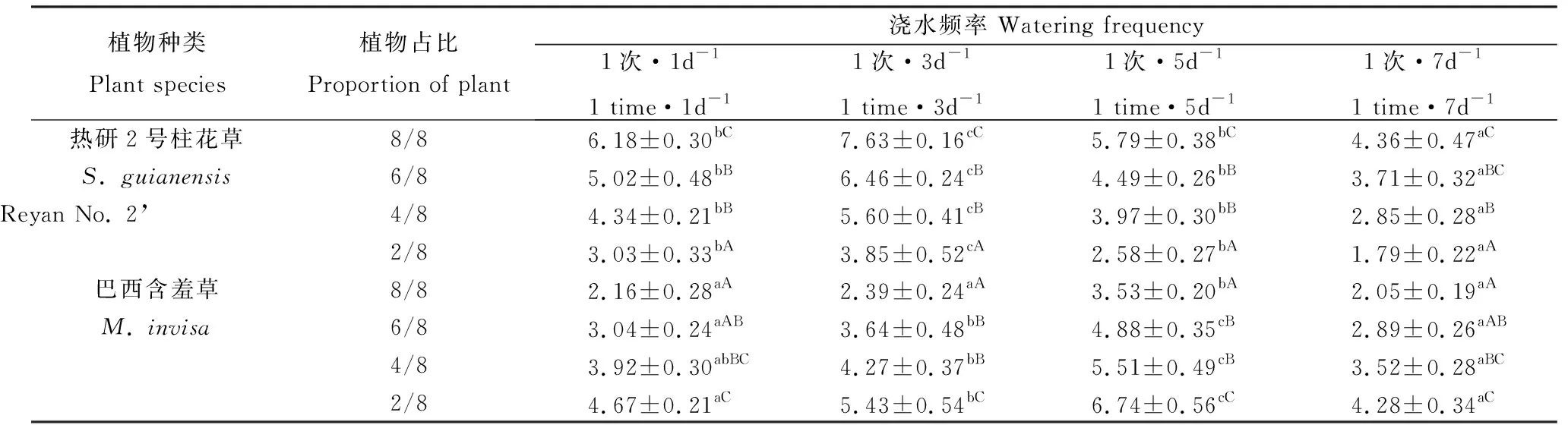
表4 澆水頻率、混播比例對熱研2號柱花草和巴西含羞草葉面積指數的影響Table 4 Effects of the mixed sowing proportion and watering frequency on the leaf area index of S. guianensis ‘Reyan No. 2’ and M. invisa
3 討論與結論
本研究結果表明,混生群落中巴西含羞草會干擾柱花草早期的正常生長,且巴西含羞草的數量越大,柱花草生長受到的影響也越大;占比4/8(即:混播比例柱:巴=4:8)時巴西含羞草對柱花草的干擾較小,二者對資源的獲取能力基本上處于相對平衡狀態,當柱:巴混播比例突破4:4(即二者株數比例1:1)時,巴西含羞草的競爭能力將迅速增大,強烈抑制柱花草的生長。因此,利用柱花草的優質牧草特性替代控制巴西含羞草蔓延滋生的前提條件為加大熱研2號柱花草的播種量,使柱花草在與巴西含羞草混生的群落中成株比例大于1:1。
植物生物量的分配是植物生存與生殖權衡的結果[22]。本試驗中2種植物在營養生長早期均優先葉部生長,以保證生長早期能夠接收有效光輻射來合成足量的光合產物,進而促進植物生物量積累,提高其競爭能力[23],混播占比不影響2種植物的總生物量。在有限的養分和水分資源條件下,隨澆水間隔期延長,2種植物的生物量均傾向于分配給養分吸收器官,增大根冠比,增加對養分的吸收,從而增強競爭能力[24-26],但巴西含羞草比柱花草耐干旱,澆水頻率較高更有利于柱花草生物量累積生長,說明植物資源競爭水平的差異會改變植物競爭對植物某一形態特征的影響效果[27]。
株高也是與植物競爭能力有關的重要因子,株高增大不但能夠提高植物的光照獲取效率[28-29],還能增加植物維持和支撐結構的比例[28]。植物株高與水分顯著相關[30],且大多數植物隨著水分減少其總葉面積會減小,形態生長受到抑制[31-32],而較大的葉面積指數更易于獲得光照,使光合作用面積增大,有利于提高植物競爭能力[23,29],同等環境條件下的植物個體僅通過對有限資源的搶先占有就能在競爭中勝出[22,26,37]。因此,本試驗中澆水頻率高有利于柱花草以較快的速度生長,1次·3d-1澆水柱花草對巴西含羞草的競爭性伸長生長更佳,要利用熱研2號柱花草替代控制巴西含羞草,就要在二者混生群落生長早期澆水頻率至少1次·3d-1,且每次澆水量為土壤最大持水量70%。但是,植物的競爭能力受其遺傳特性影響[33-34],還受諸多環境因素的影響[35-36],要通過替代控制的方法來控制外來有害植物巴西含羞草,僅僅通過混播比例和澆水頻率對其早期生長的影響還難以確定具體的替代控制效果,因為在某些環境條件下表現出強競爭力的植物,可能在另一些環境條件下變為較弱的競爭者[38],而替代控制的目的是希望利用其經濟、高效、安全和可持續利用等優點[15]。因此,要具體確定熱研2號柱花草對巴西含羞草的替代控制效果,還應當進一步考察肥力和刈割利用等因素對二者競爭情況的影響。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
