CDH17調(diào)節(jié)TGF-β自分泌影響胃癌細胞侵襲作用的研究*
2017-09-08 07:29:13孟慶斌邵永勝
重慶醫(yī)學 2017年24期
盧 昕,孟慶斌,邵永勝
(武漢市第一醫(yī)院胃腸外科 430022)
論著·基礎研究
CDH17調(diào)節(jié)TGF-β自分泌影響胃癌細胞侵襲作用的研究*
盧 昕,孟慶斌,邵永勝△
(武漢市第一醫(yī)院胃腸外科 430022)
目的 探討轉(zhuǎn)化生長因子(TGF)-β自分泌在鈣黏蛋白17(CDH17)調(diào)節(jié)胃癌細胞侵襲性中的作用及可能機制。方法 構(gòu)建siRNA-CDH17并轉(zhuǎn)染入胃癌細胞系MKN-45,沉默其表達。通過免疫熒光染色、Western blot、ELISA檢測CDH17沉默前后細胞TGF-β表達及培養(yǎng)上清液中TGF-β水平變化,觀察TGF-β的自分泌情況。同時采用Western blot檢測TGF-β/Smad3信號通路活化情況,并通過Transwell侵襲實驗觀察在給予信號通路抑制劑后,胃癌細胞侵襲力的變化,評價TGF-β自分泌及相關信號通路活化在CDH17調(diào)節(jié)胃癌細胞侵襲性中的作用。結(jié)果 轉(zhuǎn)染siRNA-CDH17沉默MKN-45細胞CDH17表達后,細胞TGF-β表達較未轉(zhuǎn)染組明顯下降,其培養(yǎng)上清液中TGF-β水平也顯著減少[(510±55.0)pg/mLvs.(115±20)pg/mL,P<0.05]。Western blot顯示CDH17沉默后Smad3磷酸化水平也明顯降低,而給予TGF-β/Smad3信號通路抑制劑SIS3(10 μmol/L)也可抑制CDH17高表達時的Smad3磷酸化水平。同時,沉默CDH17及抑制Smad3磷酸化均可明顯降低MKN-45細胞侵襲性(P<0.05)。結(jié)論 CDH17可能通過促進自分泌TGF-β活化TGF-β/Smad3信號通路參與調(diào)節(jié)胃癌細胞侵襲性。
胃腫瘤;腫瘤浸潤;轉(zhuǎn)化生長因子-β;CDH17;信號通路
目前研究發(fā)現(xiàn)鈣黏蛋白17(CDH17)是參與調(diào)節(jié)胃癌細胞侵襲及淋巴轉(zhuǎn)移的重要分子[1],然而目前其確切調(diào)控機制仍不明確,筆者前期研究發(fā)現(xiàn)CDH17 可通過誘導胃癌細胞向間質(zhì)表型轉(zhuǎn)換促進胃癌細胞侵襲,提示上皮間質(zhì)轉(zhuǎn)化可能是CDH17調(diào)控胃癌細胞侵襲性的可能機制之一。轉(zhuǎn)化生長因子(TGF)-β及相關信號通路是參與調(diào)節(jié)上皮間質(zhì)轉(zhuǎn)化的重要分子信號[2]。目前報道多種腫瘤可通過自分泌TGF參與調(diào)節(jié)自身的增殖、侵襲、轉(zhuǎn)移等生物學行為[3-4]。因此推測TGF-β自分泌也可能在CDH17誘導的胃癌細胞侵襲中發(fā)揮作用。本實驗通過比較胃癌細胞CDH17沉默前后,TGF-β的表達及分泌變化和相關信號通路活化情況,觀察其對胃癌細胞侵襲性的影響,探討TGF-β自分泌在CDH17調(diào)節(jié)胃癌細胞侵襲力中的作用及其可能機制。
1 材料與方法
1.1 主要材料和試劑 蛋白裂解液M-PER購自美國Thermo Pierce公司。CDH17和TGF-β一抗購自美國Abcam公司,磷酸化p-Smad3、總t-Smad3和SIS3一抗購自美國Cell Signaling Technology公司。β-actin一抗及相應的二抗和鏈霉親和素-生物復合物(SABC)免疫熒光試劑盒購自武漢博士德生物工程有限公司。超敏電化學發(fā)光(ECL)試劑盒購自碧云天生物技術研究所。胎牛血清和高糖DMEM培養(yǎng)基購自美國Hyclone公司。TGF-β ELISA試劑盒購自美國R&D公司。去細胞因子基質(zhì)膠購自美國Sigma-Aldrich公司。24孔嵌套Transwell(聚碳酯膜孔徑8μm)購自美國Corning公司。
1.2 方法
1.2.1 細胞培養(yǎng)及實驗分組 胃癌細胞系MKN-45由華中科技大學同濟醫(yī)學院附屬協(xié)和醫(yī)院普通外科實驗室保存提供。細胞培養(yǎng)于含10%胎牛血清的高糖DMEM完全培養(yǎng)基中,置于37℃,5%CO2培養(yǎng)箱,2~3 d傳代一次。實驗共分為3組:轉(zhuǎn)染siRNA-CDH17組;轉(zhuǎn)染CDH17空白質(zhì)粒(vector)組和非轉(zhuǎn)染組。
1.2.2 siRNA-CDH17的構(gòu)建與轉(zhuǎn)染 CDH17基因的cDNA序列由 GenBank(NM_001144663.1) 獲得,針對該序列的3種不同siRNA 由在線RNAi設計算法設計(http://www.ambion.com/techlib/misc/siRNA_finder.html)。所有設計的siRNAs由上海吉瑪生物制藥有限公司合成純化。合成純化的siRNAs 用全式金脂質(zhì)體轉(zhuǎn)染試劑盒(北京全式金生物技術有限公司)根據(jù)說明書轉(zhuǎn)染入MKN-45細胞。經(jīng)實時定量PCR 鑒定,siRNA-CDH17 序列正義鏈:5′-TGC AUT TCC UAA GGC TGA-3′,反義鏈5′- GAU CCT GGT TCA AUA ACA T-3′,具有最佳干擾效率。再將siRNA-CDH17用上述方法和試劑盒分別轉(zhuǎn)染入MKN-45細胞。細胞轉(zhuǎn)染48 h后用于實驗,細胞活性由臺盼藍染色鑒定。
1.2.3 細胞免疫熒光染色 分別將轉(zhuǎn)染與非轉(zhuǎn)染MKN-45細胞接種于蓋玻片上,以SABC法進行免疫熒光染色。4%多聚甲醛固定后,以0.1%Triton-x-100對細胞膜打孔15 min,30%過氧化氫與甲醇混合浸泡60 min以滅活內(nèi)源性過氧化物酶,洗滌后以5%牛血清清蛋白封閉60 min,分別加入CDH17和TGF-β一抗(1∶100),4℃濕盒內(nèi)孵育過夜。再滴加生物素化二抗室溫孵育20 min后加入異硫氰酸熒光素(FITC)標記的SABC熒光反應液室溫孵育20 min,脫水,透明,封片,熒光顯微鏡觀察。
1.2.4 ELISA 收集上述各組細胞培養(yǎng)上清液,按TGF-β ELISA試劑盒說明測定標準曲線后加樣,每孔終體積50 μL。用封板膜封板后置37 ℃溫育30 min后甩干、洗滌,重復5次,拍干。每孔加入酶標試劑50 μL,空白孔除外,重復上述步驟5次,拍干。每孔加入顯色劑A 50 μL,再加入顯色劑B 50 μL,輕輕振蕩混勻,37 ℃避光顯色15 min。每孔加終止液50 μL,終止反應,最后以空白孔凋零,450 nm波長依序測量各孔的吸光度。
1.2.5 Western blot 將轉(zhuǎn)染siRNA-空載體組細胞預先用SIS3(10 μmol/L)孵育24 h,然后同上述轉(zhuǎn)染和未轉(zhuǎn)染細胞洗滌加入蛋白裂解液提取總蛋白。蛋白定量后,煮沸變性后按每孔30 μg上樣,經(jīng)12%十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(DS-PAGE)凝膠電泳后轉(zhuǎn)移至PVDF膜,以5%脫脂奶粉封閉60 min,分別加入TGF-β(1∶1 000)、p-Smad3(1∶1 000)和t-Smad3(1∶1 000)及β-actin(1∶1 000)一抗4 ℃孵育過夜,二抗室溫孵育1 h后,采用ECL,以Bio-Rad熒光成像系統(tǒng)顯影。目的蛋白表達以β-actin作為參照相對定量。
1.2.6 Transwell侵襲實驗 將上述經(jīng)SIS3孵育的細胞及轉(zhuǎn)染和未轉(zhuǎn)染細胞按1×105/孔接種于鋪有去細胞因子基質(zhì)膠的24孔Transwell嵌套(濾膜孔徑為8 μm)中,上室加入不含血清的高糖DMEM培養(yǎng)基,下室均加入含10%胎牛血清的完全培養(yǎng)基,孵育8 h后,蘇木素染色,顯微鏡下計數(shù)穿膜細胞。
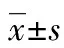
2 結(jié) 果
2.1 siRNA-CDH17轉(zhuǎn)染對TGF-β表達的影響 細胞免疫熒光顯示轉(zhuǎn)染siRNA-CDH17后,MKN-45細胞CDH17及TGF-β表達明顯下降,Western blot也顯示沉默CDH17表達后,TGF-β表達也明顯減少。轉(zhuǎn)染siRNA-vector(空載體)組與未轉(zhuǎn)染對照組TGF-β呈現(xiàn)相對高表達,二者間表達水平比較差異無統(tǒng)計學意義(圖1)。
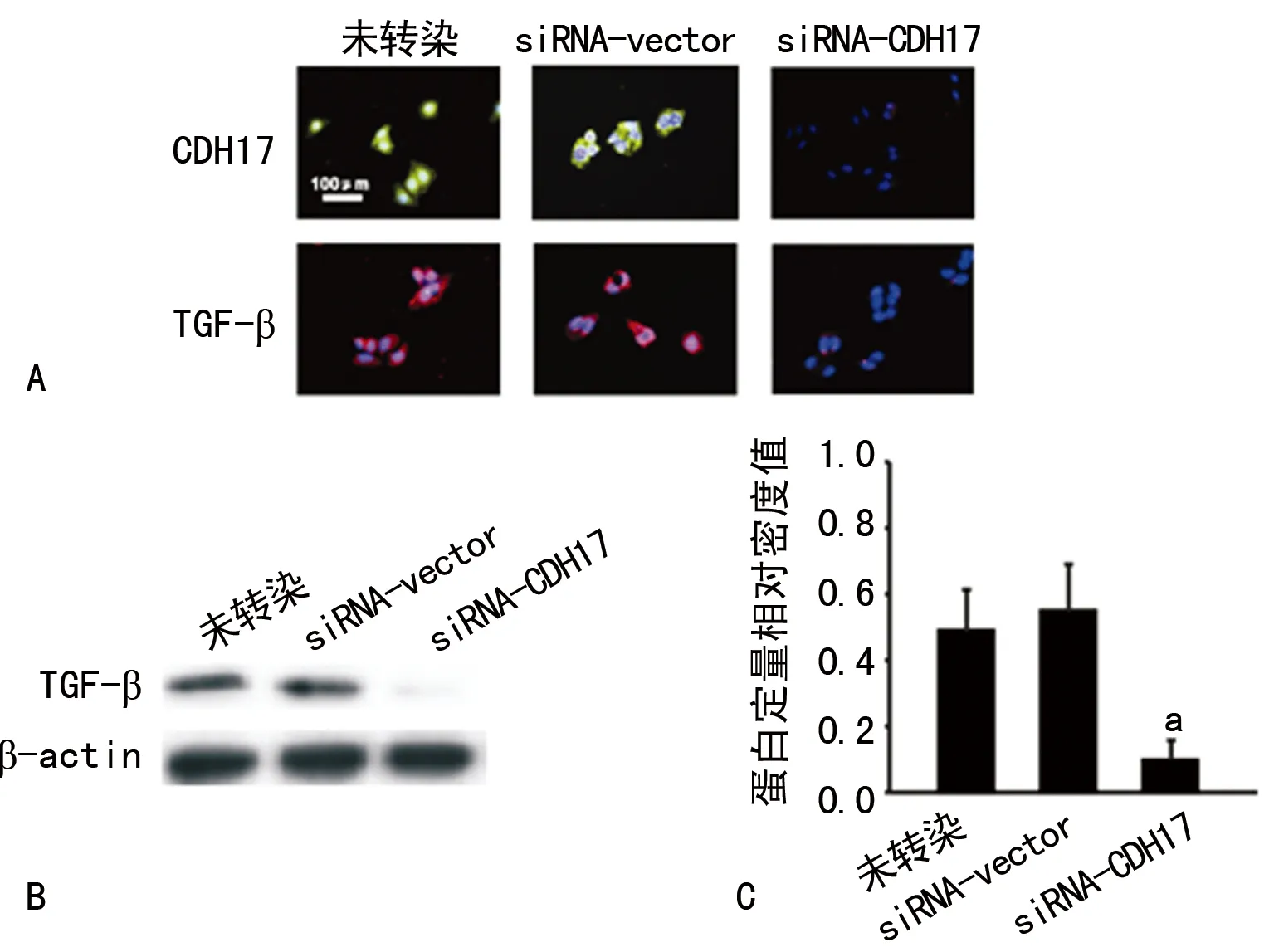
A:免疫熒光染色;B:Western blot;C:Western blot相對定量。a:P<0.05
圖1 siRNA-CDH17轉(zhuǎn)染對TGF-β表達的影響
2.2 siRNA-CDH17轉(zhuǎn)染對TGF-β分泌的影響 ELISA檢測顯示,轉(zhuǎn)染siRNA-CDH17,沉默MKN-45細胞CDH17表達后,其培養(yǎng)上清液中TGF-β水平明顯降低[(510±55)pg/mLvs.(115±20) pg/mL,P<0.05],而轉(zhuǎn)染siRNA-vector組與未轉(zhuǎn)染組上清液中TGF-β水平相對較高,兩組比較差異無統(tǒng)計學意義[(510±55)pg/mLvs.(485±38) pg/mL,P>0.05]。
2.3 siRNA-CDH17轉(zhuǎn)染對TGF-β/Smad3信號通路活化的影響 轉(zhuǎn)染siRNA-CDH17,沉默CDH17表達后,胃癌MKN-45細胞Smad3磷酸化水平較未轉(zhuǎn)染組明顯下降(P<0.05),而轉(zhuǎn)染siRNA-vector組與未轉(zhuǎn)染組細胞Smad3磷酸化水平較高,兩組比較差異無統(tǒng)計學意義(P>0.05)。而TGF-β/Smad3信號通路抑制劑SIS3可明顯減少siRNA-vector組的Smad3磷酸化水平(P<0.05),見圖2。
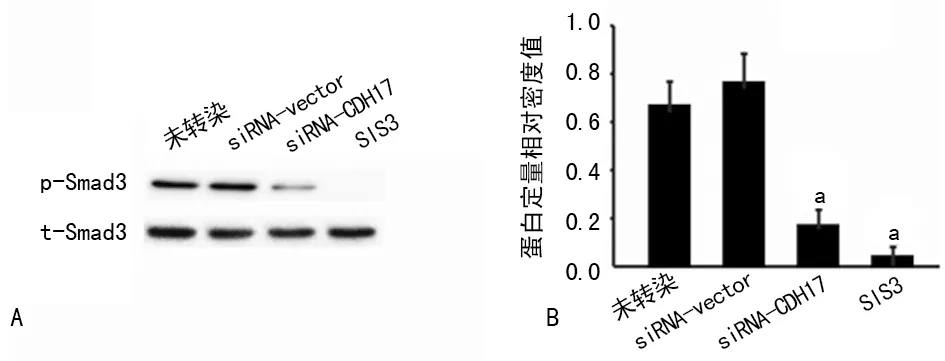
A:Western blot;B:Western blot相對定量。a:P<0.05
圖2 siRNA-CDH17轉(zhuǎn)染對TGF-β/Smad3信號通路活化的影響
2.4 TGF-β/Smad3信號通路對MKN-45細胞侵襲性的影響 Transwell侵襲實驗檢測顯示,較未轉(zhuǎn)染組(9 525±1 000)及siRNA-vector組(9 525±1 000),siRNA-CDH17轉(zhuǎn)染(2 650±410)及應用SIS3處理(3 800±525)均可明顯減少胃癌MKN-45細胞的侵襲數(shù),差異有統(tǒng)計學意義(P<0.05)。
3 討 論
前期研究發(fā)現(xiàn)CDH17表達可誘導胃癌細胞向間質(zhì)細胞表型轉(zhuǎn)化,提示上皮間質(zhì)轉(zhuǎn)化可能在CDH17調(diào)控胃癌細胞轉(zhuǎn)移、侵襲中發(fā)揮作用。因此探討上皮間質(zhì)轉(zhuǎn)化相關信號機制與CDH17侵襲調(diào)控間的關系,有助于進一步闡明胃癌侵襲調(diào)控的信號生物學機制。本實驗通過檢測CDH17對胃癌細胞TGF-β產(chǎn)生和TGF-β/Smad3信號通路活化的影響,以及TGF-β/Smad3信號活化在胃癌細胞侵襲中的作用,探討TGF-β及其相關信號通路在CDH17調(diào)控胃癌細胞侵襲性中的可能作用及機制。
本研究發(fā)現(xiàn)在胃癌細胞高表達CDH17時,細胞TGF-β的表達及分泌均較高,而抑制CDH17表達后,TGF-β的表達及分泌均明顯減少,提示CDH17可能對胃癌細胞TGF-β的自分泌發(fā)揮作用。盡管CDH17是鈣黏蛋白家族成員,通常被作為下游蛋白受上游分子信號調(diào)節(jié)。但近來研究表明,采用siRNA干擾技術,抑制CDH17的基因表達,可抑制上游β-catenin/Wnt和Ras/MEK信號通路活化,進而影響胃癌細胞的侵襲[5-6]。本研究則發(fā)現(xiàn),CDH17高表達伴隨的TGF-β高表達可增加Smad3磷酸化水平,而沉默CDH17表達后,在TGF-β產(chǎn)生減少的同時,Smad3磷酸化水平也顯著下降,表明CDH17也可介導上游TGF-β/Smad3信號通路活化,進而促進胃癌細胞侵襲。
TGF-β/Smad3信號通路在介導上皮間質(zhì)轉(zhuǎn)化(epithelial-mesenchymal transition,EMT)中發(fā)揮最重要功能[7]。而EMT通過細胞表型向間質(zhì)轉(zhuǎn)化,使細胞高表達基質(zhì)金屬蛋白酶,從而易于分解細胞外基質(zhì),利于細胞遷移;同時通過間質(zhì)轉(zhuǎn)化本身,細胞內(nèi)骨架重排,遷移能力增強也有利于細胞遷移力及侵襲性增強[8]。前期研究證實,CDH17可通過誘導胃癌細胞EMT促進其侵襲。本研究也發(fā)現(xiàn),Smad3磷酸化水平與胃癌細胞侵襲性相關,無論抑制CDH17表達或直接抑制Smad3活化均可顯著減少胃癌細胞侵襲。進一步證實TGF-β/Smad3信號通路可能通過誘導EMT在介導胃癌細胞侵襲性中發(fā)揮重要作用。
本研究發(fā)現(xiàn)為CDH17在調(diào)節(jié)胃癌細胞侵襲性中的作用提供了又一新的分子細胞生物學機制,較以往的研究不僅關注分子信號機制本身,結(jié)合前期研究更進一步揭示分子信號與侵襲終效應間的細胞生物學行為聯(lián)系。但由于β-catenin/Wnt和Ras/MEK等信號通路也在CDH17介導的胃癌細胞侵襲中發(fā)揮重要作用,且細胞信號本身存在多種信號交聯(lián)機制。因此進一步深入探尋信號交聯(lián)分子在其中的作用,更有助于進一步闡明胃癌細胞侵襲轉(zhuǎn)移機制。同時CDH17介導上游信號通路的活化調(diào)節(jié)是直接作用還是需要中間分子機制參與是下一個研究目標。
[1]Park SS,Kang SH,Park JM,et al.Expression of liver-intestine cadherin and its correlation with lymph node metastasis in gastric cancer:can it predict N stage preoperatively? [J].Ann Surg Oncol,2007,14(1):94-99.
[2]Zavadil J,Bottinger EP.TGF-beta and epithelial-to-mesenchymal transitions [J].Oncogene,2005,24(37):5764-5774.
[3]蔡洪培,高勇,張翼翔.人肝癌細胞系SMMC-7721細胞TGF-α 的自分泌[J].第二軍醫(yī)大學學報,2001,22(4):393-394.
[4]洪祖蓓,狄文,丁傳偉.TGF-α/EGFR自分泌環(huán)調(diào)控卵巢癌細胞增殖轉(zhuǎn)移的分子機制[J].中國癌癥雜志,2006,16(11):899-902.
[5]Qiu HB,Zhang LY,Ren C,et al.Targeting CDH17 suppresses tumor progression in gastric cancer by downregulating Wnt/β-catenin signaling[J].PLoS One,2013,8(3):e56959.
[6]Wang J,Kang WM,Yu JC,et al.Cadherin-17 induces tumorigenesis and lymphatic metastasis in gastric cancer through activation of NFκB signaling pathway [J].Cancer Biol Ther,2013,14(3):262-270.
[7]Micalizzi DS,Farabaugh SM,Ford HL.Epithelial-mesenchymal transition in cancer:parallels between normal development and tumor progression[J].J Mammary Gland Biol Neoplasia,2010,15(2):117-134.
[8]Yang J,Weinberg RA.Epithelial-mesenchymal transition:at the crossroads of development and tumor metastasis[J].Dev Cell,2008,14(6):818-829.
Study on role of CDH17 regulating TGF-β autocrine for affecting invasion of gastric cancer cells*
LuXin,MengQingbin,ShaoYongsheng△
(DepartmentofGastrointestinalSurgery,WuhanMunicipalNo.1Hospital,Wuhan,Hubei430022,China)
Objective To explore the role and possible mechanism of transforming growth factor (TGF)-β autocrine in CDH17 regulating invasion of gastric cancer cells.Methods Construction and transfection of siRNA-CDH17 into MKN-45 gastric cancer cell line to silence the expression of CDH17.Expression of TGF-β and concentrations of TGF-β in supernatants were detected before and after CDH17 silence by immunofluorescence,immunoblotting and ELISA.The autocrine situation of TGF-β was observed.Meanwhile,the activation of TGF-β/Smad3 signaling pathway was also detected by immunoblot.After giving signaling pathway inhibitor,the changes of invasion ability of MKN-45 cells were observed by Transwell invasion experiment.The role of TGF-β autocrine and related signaling pathway activation in CDH17-regulated invasion of gastric cancer cells was evaluated.Results After transfecting siRNA-CDH17 for silencing CDH17 expression in MKN-45 cells,the expression of TGF-β was significantly decreased compared with non-transfection group,its concentration in supernatants was also significantly reduced[(510±55)pg/mLvs. (115±20) pg/mL,P<0.01].The immunoblots revealed that phosphorylation level of Smad3 after CDH17 silence was also significantly diminished.However,giving the TGF-β/Smad3 signaling inhibitor SIS3 (10 μmol/L) could also suppress the phosphorylation level of Smad3 when CDH17 was highly expressed,meanwhile silencing CDH17 and inhibiting Smad3 phosphorylation could significantly decrease the invasion of MKN-45 gastric cancer cells (P<0.05).Conclusion CDH17 could participate in the invasion of gastric cancer cells by promoting TGF-β autocrine to activate TGF-β/Smad3 signaling pathway.
stomach neoplasms;neoplasm invasiveness;transforming growth factor -β;CDH17;signaling pathway
2015年武漢市衛(wèi)生和計劃生育委員會科研項目(WX15C31).
盧昕(1971-),副主任醫(yī)師,博士,主要從事胃腸腫瘤分子生物學研究。
△通信作者,E-mail:shaoyongsheng1211@medmail.com.cn。
10.3969/j.issn.1671-8348.2017.24.003
R735.2
A
1671-8348(2017)24-3321-03
2016-12-19
2017-03-07)
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
文苑(2018年21期)2018-11-09 01:23:06
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國衛(wèi)生標準管理(2015年3期)2016-01-14 03:41:46
中國衛(wèi)生(2015年9期)2015-11-10 03:11:12
醫(yī)學研究雜志(2015年9期)2015-07-01 17:28:27
中國當代醫(yī)藥(2015年20期)2015-03-01 02:04:29
中國衛(wèi)生(2014年3期)2014-11-12 13:18:12
