福建漳浦凡納濱對蝦海水養殖中后期水體細菌群落多樣性分析
2017-08-10 12:17:13胡東王麗萍趙苒邵宗澤
海洋學報 2017年8期
關鍵詞:變形
胡東,王麗萍,趙苒,邵宗澤*
(1.廈門大學 公共衛生學院,福建 廈門 361102;2.國家海洋局第三海洋研究所 國家海洋局海洋生物遺傳資源重點實驗室,福建 廈門 361005)
?
福建漳浦凡納濱對蝦海水養殖中后期水體細菌群落多樣性分析
胡東1,2,王麗萍2,趙苒1,邵宗澤2*
(1.廈門大學 公共衛生學院,福建 廈門 361102;2.國家海洋局第三海洋研究所 國家海洋局海洋生物遺傳資源重點實驗室,福建 廈門 361005)
凡納濱對蝦;微生物群落多樣性;16S rRNA基因高通量測序;水質因子
1 引言
凡納濱對蝦(Litopenaeusvannamei)因其具有生長快、抗病能力強、養殖經濟效益顯著,成為當今世界上水產養殖產量最高的三大優良對蝦品種之一[1],其2011年產量占全球對蝦產量的76%[2]。凡納濱對蝦養殖要經歷幼蟲到成體5個生長階段[3],大約3~4個月的生長周期才可以上市。養殖動物腸道及養殖水體中微生物的種類和數量[4—5],水質因子[6](溶氧、溫度、pH、化學需氧量、氮等),飼料投喂,藻類生長狀況等是對蝦能否健康成長的關鍵因素,了解養殖水體中微生物群落結構、多樣性變化及微生物菌群與水體環境因子的相互作用對改善養殖水體、預防對蝦疾病發生至關重要。
第二代測序技術為我們研究環境中微生物多樣性提供了高效的方法,結合微生物獨有的16S rRNA保守基因,這種方法被廣泛應用到環境微生物多樣性分析[5, 7—9]。16S rRNA高通量測序技術已用于淡水環境凡納濱對蝦養殖微生物多樣性分析,Zhang等發現在凡納濱對蝦淡水養殖池中,變形菌門、藍細菌、放線菌門和擬桿菌門是水體微生物的主要類群;變形菌門、藍細菌、厚壁菌門、酸酐菌門、綠灣菌門、擬桿菌門是沉積物環境的主要類群[7]。但鮮有對蝦海水養殖環境微生物菌群的相關研究報道,因此,我們的研究可以彌補這方面的空白,豐富人們對凡納濱對蝦養殖環境微生物菌群多樣性的認識。
本研究利用Illumina高通量測序技術分析了凡納濱對蝦高位池養殖水體細菌多樣性的動態變化過程,并分析了菌群與水質參數間的相互關系。研究結果將有助于了解對蝦不同養殖時期水體菌群變化規律,認識影響對蝦養殖水體菌群變化的環境因子,以期通過調控水質因子,達到改善養殖環境微生物的群落結構的目的,從而促進對蝦健康養殖。
2 材料和方法
2.1 樣品采集
采樣點為福建漳浦縣凡納濱對蝦海水養殖場高位池[5號池,0.9畝(1畝=666.67 m2),40萬蝦苗] (24°01′43″ N,117°50′39″E)。采樣時間為2015年12月24日至2016年3月24日,即對蝦蝦苗度過“早期死亡綜合癥”到對蝦收成,實驗周期為3個月。在離岸2 m和蝦池中心設置2個采樣點(S1和S2,第131天只有S1點),使用經過高溫濕熱滅菌處理的有機玻璃采水器,采集上、中、下3層水樣共3 L于無菌采樣瓶中,共采集了7個時間點(養殖第40、59、67、77、95、115、131天 )的13個水體樣品。采集的水樣命名為P5D&S#,P5表示5號池,D&表示采樣時間編號,S#表示采樣點編號(下同)。水樣立即置于放有冰袋的樣品箱內保存,并運回實驗室用0.22 μm濾膜過濾,濾膜于-80℃冰箱中保存。
2.2 水質參數檢測

表1 主要水質指標及檢測方法
2.3 基因組DNA提取
利用MOBIO水樣基因組提取試劑盒(14900-100-NF,PowerWater? DNA Isolation Kit)進行基因組DNA提取。提取的基因組DNA質量和濃度分別用1%瓊脂糖凝膠電泳和nanodrop2000超微量分光光度計檢測,提取的DNA在-80℃冰箱中保存。
2.4 16S rRNA基因高通量測序
提取的基因組DNA送上海美吉生物醫藥科技有限公司,采用引物338F(5′-3′,ACTCCTACGGGAGGCAGCAG)和806R(5′-3′,GGACTACHVGGGTWTCTAAT)[10]對樣品進行PCR擴增,引物兩端帶有特異的barcode序列,以識別不同樣本的16S rRNA基因擴增產物;隨后采用Miseq PE300測序平臺對PCR產物測序。
2.5 微生物多樣性分析
對原始數據進行拼接、過濾,得到優化序列。然后基于有效數據將相似性達97%的序列進行OTU(Operational Taxonomic Units)聚類。采用RDP classifier貝葉斯算法[11]對97%相似水平的OTU代表序列進行分類學分析,并在各水平統計每個樣品的群落組成。基于OTU聚類分析結果,對OTU進行多種多樣性指數分析,如Alpha多樣性指數[12];基于分類學信息,進行群落結構的統計分析[13];基于系統發育,進行unifrac等分析[14];并結合水質因子進行RDA關聯分析。在上述分析的基礎上,進行一系列群落結構和系統發育等深入的統計學和可視化分析。
3 結果
3.1 養殖池水體的水質參數
為了探討水質與水體環境微生物菌群多樣性的關系,對采集水樣進行水質分析(表2)。在養殖中后期,溶氧基本維持在飽和狀態(6.0~8.0 mg/L);溫度變化范圍為 20~24.6℃,pH維持在弱堿性(7.2~7.5),變化較小;COD波動較大,變化范圍為6.9~12.0 mg/L;硝酸鹽氮值變化范圍為0.96~1.49 mg/L,亞硝酸鹽氮值變化范圍為0.02~0.35 mg/L,氨態氮值變化較大,波動范圍為0.11~4.96 mg/L,其中硝態氮的濃度變化與以往研究明顯不同。本研究發現,硝酸鹽濃度在養殖過程中總體上呈下降趨勢,而以往的報道是養殖后期濃度升高[15—16]。

表2 養殖池水質參數的7次調查結果
3.2 Illumina 序列多樣性評價
采用Illumina miseq測序平臺對采集的7個時間點13個水樣進行16S rRNA基因高通量測定,13個樣品中共得到823 363條序列,序列長度在281~520 bp之間,每個樣品的序列數目在55 449~73 485條之間,抽平后得到619 775條高質量序列,每個樣品的序列數目為47 675條。稀釋曲線分析(圖1a)顯示,隨著測序數量的增加,稀釋曲線逐漸趨于平緩,表明該測序量已經基本能夠反映該實驗中細菌群落多樣性。按照97%相似度對OTU進行劃分,其覆蓋度均大于 0.99,每個樣品得到的OTU數目在294~412之間。通過Shannon-Wiener多樣性指數對養殖水體細菌群落多樣性進行評估,如圖1b所示,第40天樣品微生物多樣性最高,而第67天樣品微生物多樣性最低,其余時間點采集的樣品微生物多樣性大致相似。
3.3 主成分和聚類分析
主成分分析(Pcoa)結果顯示(圖2a),每個時間點采集的兩個重復樣本都能很好地聚在一起,說明相同時間采集的重復樣本細菌群落多樣性相似性較高。Unifrac聚類分析和科水平的細菌群落Heatmap分析(圖2b,2c)也是如此。此外,其他水平的細菌群落Heatmap分析結果與科水平的細菌群落Heatmap分析結果基本一致,相同時間點采集的兩個重復樣本都能很好的聚在一起,不同時間差異較大。但是,不同時間段之間仍有一定的相似性,第40、77、95與131天之間的細菌組成相似,而第67天與115天細菌組成較為相似。
圖1 各樣品的細菌16S rRNA基因的α多樣性分析Fig.1 Diversity analysis of 16S rRNA genes in different samplesa.不同樣品的稀釋曲線;b.不同樣品的香農威納指數圖a.Rarefaction curve of different samples; b.Shannon-Wiener rarefaction curve plot of different samples
圖2 不同樣品細菌群落相似性分析Fig.2 Similarity analysis of microbial populations in different samplesa. 細菌群落的Pcoa分析; b. 科水平的細菌群落heatmap; c. 基于Unifrac的聚類分析a. Pcoa analysis; b. heatmap of bacterial populations at the family level; c. cluster analysis based Unifrac
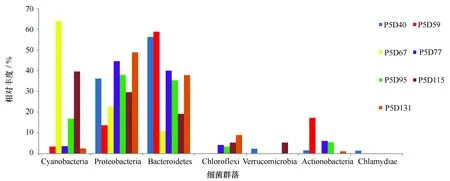
圖3 門水平細菌群落組成Fig.3 Bacterial community composition at the phylm level
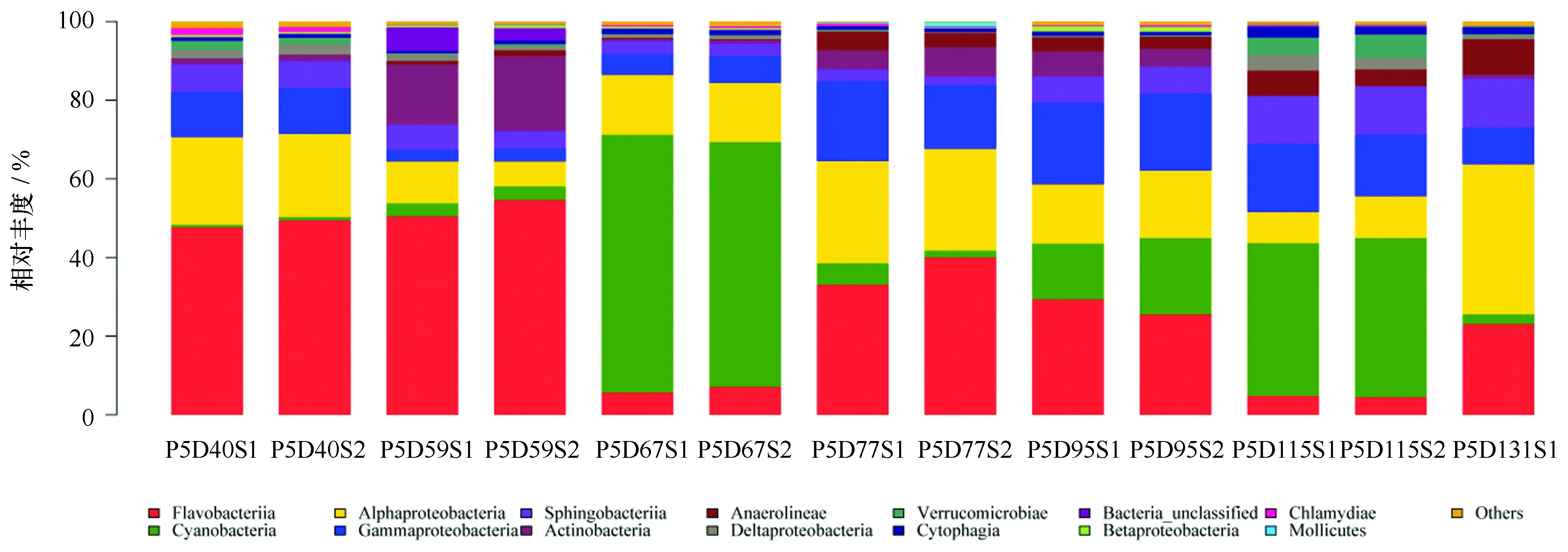
圖4 綱水平上細菌群落變化Fig.4 Changes of bacterial community in all samples at the class level
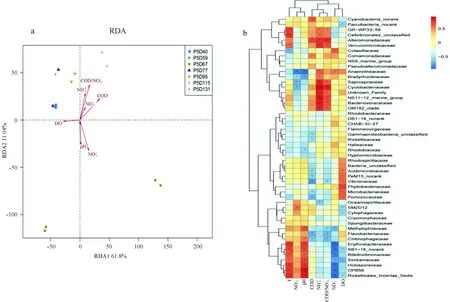
圖5 水質參數與細菌群落之間的關系Fig.5 The correlation between water quality parameters and bacterial community compositiona.細菌群落組成與水質的RDA分析; b.科水平上細菌群落與水質相關性熱圖a. Redundancy analysis (RDA) of the bacterial community composition with water quality parameters (red arrows); b. heatmap showing the correlation between bacterial community and the water quality parameters at the family level
3.4 細菌多樣性組成
在本研究采集的7個時間點13個樣品中,共檢測到19個門、35個綱、80個目、135個科、254個屬的細菌。另外,未能鑒定的序列數總共為5 337條,這表明樣本中存在著大量未知的類群。根據樣品中各微生物所測定的reads數,把每個樣品中reads數大于總reads數10%的門定義為優勢門。在不同養殖時期水樣檢測中,藍細菌(Cyanobacteria)、變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)、綠灣菌門(Chloroflexi)、疣微菌門(Verrucomicrobia)、放線菌門(Actinobacteria)、衣原體(Chlamydiae)等7個門是主要的優勢類群(圖3)。
根據細菌群落組成圖可知,養殖水體中的絕大多數序列可以歸類于變形菌門和擬桿菌門(圖3),并且這2個門在采樣期間一直是主要類群。其中,變形菌門在7個采樣時間點所占的比例分別為36.2%、13.7%、22.7%、44.6%、38.0%、29.6%、48.8%,主要包括α-變形菌綱(Alphaproteobacteria)和γ-變形菌綱(Gmmaproteobacteria)(圖4),α-變形菌綱在第59、115天樣品豐度較低(分別為8.5%和9.3%),而在其他采樣時間點豐度普遍較高(豐度變化為15.1%~38.1%),γ-變形菌綱則在第59、67天豐度較低(比例分別為3.2%和6.1%),其他采樣時間點豐度普遍偏高(豐度變化為9.3%~20.1%);擬桿菌門在7個采樣時間點所占的比例分別為56.3%、58.8%、10.7%、40.0%、35.4%、19.2%、37.9%,主要包括黃桿菌綱(Flavobacteriia)和Sphingobacteriia (圖4),黃桿菌綱在第67、115天樣品豐度較低(分別為6.3%和4.7%),而在其他采樣時間點豐度普遍偏高(豐度變化為23.1%~52.6%),Sphingobacteriia在養殖后期豐度較高(各時間點所占比例為7.0%、5.4%、3.2%、2.6%、6.7%、12.2%、12.4%)。藍細菌為養殖過程中處于次優勢的一個類群,在7個采樣時間點波動較大,所占的比例分別為0.7%、3.4%、63.9%、3.6%、16.8%、39.7%、2.5%(圖3)。
整個采樣期間,黃桿菌科(Flavobacteriaceae)、紅桿菌科(Rhodobacteraceae)、Cryomorphaceae、Cyanobacteria_norank、腐螺旋菌科(Saprospiraceae)、紅菌科(Rhodobiaceae)等優勢類群(圖2b),廣泛存在于各個養殖時期,并且基本都是優勢菌群。擬桿菌門的黃桿菌科在整個采樣階段占優勢地位,最高比例可達48.0%,但第67、115天所占比例僅為2.5%、1.5%。α-變形菌綱的紅桿菌科在整個采樣期間也都被檢測到,從第40天占整個種群的13.4%降低到第59、67天的3.1%和5.1%,在第77天又成為優勢種群(19.2%),隨后在第95、115、131天分別降低到9.4%、4.8%和10.7%;相似的現象在擬桿菌門的 Cryomorphaceae上被體現,其在整個種群中所占比例由第40天的13.8%減少到第59、67天的3.3%和3.2%,在第77天成為優勢種群(13.6%),隨后在第95、115、131天分別降低到5.8%、2.7%和5.3%。屬于擬桿菌門的腐螺旋菌科在整個采樣期間也是優勢類群,其在整個菌群中所占比例由第40天到第95天的低比例(所占比例分別為5.0%、2.6%、2.7%、1.7%、6.4%)升高到第115、131天的10.4%和12.0%;相似的現象也在α-變形菌綱的紅菌科上得到體現,其在整個種群中所占比例由采樣前期低比例(第40天、59天所占比例分別為0%和2.5%)升高到第131天的23.1%。Cyanobacteria_norank表現出與門水平藍細菌相同的變化,所占比例變化為0.7%~63.8%。
3.5 細菌群落與水質參數關系
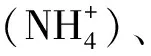
4 討論
本研究通過Illumina高通量測序技術初步揭示了凡納濱對蝦養殖中后期養殖水體中細菌群落多樣性變化及其與水質之間的相互關系。Pcoa分析和聚類分析表明,養殖中期第40天和第59天細菌群落組成相似,第77、95天與養殖后期第131天細菌群落組成更相似,而第67天和第115天樣品與其他時間點的樣品差異較大。此外,同一個時間點采集的兩個樣品重復性較好,說明數據較為可靠。從7個時間點采集的13個水體樣品中共獲得619 775條高質量序列,檢測到的細菌群落歸屬于19個門、35個綱、80個目、135個科、254個屬,說明該養殖水體中細菌群落具有很高的多樣性。注釋結果表明,這些序列主要屬于變形菌門、擬桿菌門和藍細菌等3個門類。此外,未能鑒定的序列數總共為5 337條,占總序列數比例為0.86%,說明凡納濱對蝦養殖環境中還存在著大量未描述過的類群。
在本研究中,變形菌門、擬桿菌門廣泛存在于凡納濱對蝦各個養殖時期的水體中,并且為優勢類群。相似的結果在前人的研究中也發現,在凡納濱對蝦淡水養殖系統中,養殖水體溫度高時,放線菌門、unclassified bacteria、變形菌門和擬桿菌門是主要的類群;養殖水體溫度低時,主要的優勢類群是放線菌門、變形菌門和擬桿菌門[18]。同樣,Zhang等發現在凡納濱對蝦淡水養殖池中,變形菌門、藍細菌、放線菌門和擬桿菌門是水體微生物的主要類群[7]。Huang等研究發現,不同生長時期凡納濱對蝦腸道的優勢類群是變形菌門、擬桿菌門和放線菌門[5]。由此可見,變形菌門和擬桿菌門是凡納濱對蝦腸道和水體環境常見的優勢類群。此外,在本研究中藍細菌在第67、115天樣品中占絕對優勢,文獻報道光照強并且營養鹽豐富,有利于藍細菌繁殖[19],但是實驗記錄同時期為陰雨天氣,說明其他環境參數影響著藍細菌豐度變化。
盡管不同養殖時期水體微生物在門水平上組成較相似,但在綱水平和科水平上,細菌類群構成變化較大。在綱水平上,黃桿菌綱、α-變形菌綱和γ-變形菌綱為第40天樣品的優勢類群,第59天樣品的主要類群為黃桿菌綱和放線菌綱(Actinobacteria),而第67天樣品的優勢類群波動較大,轉變為Cyanobacteria_norank和α-變形菌綱。第77、95、131天樣品菌群以黃桿菌綱、α-變形菌綱、和γ-變形菌綱為主,而第115天樣品的優勢類群為藍細菌、γ-變形菌綱和Sphingobacteriia。可以看出α-變形菌綱、γ-變形菌綱和黃桿菌綱是綱水平上的主要優勢類群。變形細菌是所有細菌中最大和最具多樣性的一個族群,具有較強的適應性,它們在水體中分布廣泛[20],其中α-和γ-變形菌綱在海水浮游細菌中尤為重要。已有的關于黃桿菌的研究表明,黃桿菌可以改善底質環境狀況,在維持菌落方面起重要作用,其多為有益菌[20],這可能是該蝦池健康養殖的主要原因。在科水平上,黃桿菌科、紅桿菌科、Cryomorphaceae、Cyanobacteria_norank、腐螺旋菌科、紅菌科等類群,廣泛存在于養殖水體中,并且為各個養殖時期的優勢或者次優勢菌。科水平上,黃桿菌科、紅桿菌科和Cryomorphaceae為第40天樣品的優勢菌類群,黃桿菌科和微桿菌科(Microbacteriaceae)為第59天樣品的優勢類群,藍細菌為第67天樣品的優勢菌,黃桿菌科、紅桿菌科和Cryomorphaceae 為第77天樣品的優勢類群,黃桿菌科、藍細菌和科韋爾氏科(Colwelliaceae)為第95天樣品的優勢類群,藍細菌、腐螺旋菌科和交替單胞菌科(Alteromonadaceae)為第115天樣品的優勢菌類群,交替單胞菌科、紅桿菌科、腐螺旋菌科和紅菌科為第131天樣品的優勢類群。黃桿菌科在整個采樣期間是占優勢的微生物類群,研究報道表明黃桿菌(Flavobacteriumpsychrophilum)在鱒魚(Trout)養殖中是一類嚴重的病原菌,可以導致虹鱒魚魚苗綜合癥(RTFS)和冷水病(CWT)[21];Tamlana作為一種潛在的益生菌可以產生瓊脂酶[22];Tenacibaculummaritimum可以導致魚類產生gliding bacterial disease和 tenacibaculosis/flexibacteriosis等疾病[23]。同樣紅細菌科在整個采樣期間是占優勢的微生物類群,研究表明玫瑰桿菌(Roseobacter)能在大菱鲆(Scophthatmusmaximus) 仔魚的飼養環境中建立一個有益的微生物群落,可以抑制致病菌的生存[24],降低大菱鲆幼魚的死亡率[25]。值得注意的是,上述這些主要的優勢類群在養殖的第59、67、115天表現出相對較低的豐度,如綱水平上α-變形菌綱在第59、115天豐度較低,γ-變形菌綱在59、67天豐度較低,黃桿菌綱在67、115天豐度較低;科水平上黃桿菌科在第67、115天豐度較低,紅桿菌科、Cryomorphaceae在第57、67、115天豐度較低。
在整個采樣期間還檢測到黃桿菌屬(Flavobacterium),希瓦氏菌屬(Shewanella),弧菌屬(Vibrio),假單胞菌屬(Pseudomonas)和Escherichia-Shigella等水生動物潛在的條件致病菌[7, 24, 26—27],除黃桿菌屬和弧菌屬外,其他菌屬在所有樣品中都維持低豐度(小于0.1%)。其中弧菌為對蝦常見的致病菌,已知副溶血弧菌(Vibrioparahemolyticus)、哈維氏弧菌(V.harveyi)、坎貝氏弧菌(V.campbellii)都是凡納濱對蝦主要的致病菌[28],本研究中發現的弧菌主要為坎貝氏弧菌。潛在致病菌低reads數表明在進水溫度適宜的情況下,增加換水頻數不失為一種預防對蝦病害發生的有效方法。此外,本研究在對各個時期養殖水體微生物的多樣性分析中,同樣檢測到一定豐度的潛在益生菌,主要包括芽孢桿菌屬(Bacillus)[29]、蛭弧菌屬(Bdellovibrio)[24]、擬桿菌屬(Bacteroides)[7]、乳球菌屬(Lactococcus)[24],盡管它們的豐度低,但這些益生菌會降低潛在致病菌等危害因素導致對蝦發病幾率增加的可能性。
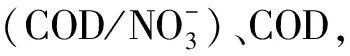
5 結論
致謝:感謝國家海洋局第三海洋研究所海化實驗室林彩老師對水質參數COD測定提供的幫助;感謝國家海洋局第三海洋研究所海洋生物遺傳資源重點實驗室楊洋、楊碩、黃兆斌、劉陽對水樣采集提供的幫助。感謝廈門大學公共衛生學院郭東北老師提供的水樣采集儀器。
[1] Tu C, Huang H T, Chuang S H, et al. Taura syndrome in pacific white shrimp penaeus vannamei cultured in Taiwan[J]. Diseases of Aquatic Organisms, 1999, 38(2): 159-161.
[2] Yu Yang, Wei Jiankai, Zhang Xiaojun, et al. SNP discovery in the transcriptome of white Pacific shrimpLitopenaeusvannameiby next generation sequencing[J]. PLoS One, 2014, 9(1): e87218.
[3] Zheng Yanfen, Yu Min, Liu Yan, et al. Comparison of cultivable bacterial communities associated with Pacific white shrimp (Litopenaeusvannamei) larvae at different health statuses and growth stages[J]. Aquaculture, 2016, 451: 163-169.
[4] Boutin S, Bernatchez L, Audet C, et al. Network analysis highlights complex interactions between pathogen, host and commensal microbiota[J]. PLoS One, 2013, 8(12): e84772.
[5] Huang Zhaobin, Li Xiaoyi, Wang Liping, et al. Changes in the intestinal bacterial community during the growth of white shrimp,Litopenaeusvannamei[J]. Aquaculture Research, 2014, 47(6): 1737-1746.
[6] Berdjeb L, Ghiglione J F, Jacquet S. Bottom-up versus top-down control of hypo-and epilimnion free-living bacterial community structures in two neighboring freshwater lakes[J]. Applied and Environmental Microbiology, 2011, 77(11): 3591-3599.
[7] Zhang Hao, Sun Zhenli, Liu Bo, et al. Dynamic changes of microbial communities inLitopenaeusvannameicultures and the effects of environmental factors[J]. Aquaculture, 2016, 455: 97-108.
[8] Rosselli R, Romoli O, Vitulo N, et al. Direct 16S rRNA-seq from bacterial communities: a PCR-independent approach to simultaneously assess microbial diversity and functional activity potential of each taxon[J]. Scientific Reports, 2016, 6: 32165.
[9] Yan Qingyun, Bi Yonghong, Deng Ye, et al. Impacts of the three gorges dam on microbial structure and potential function[J]. Scientific Reports, 2015, 5: 8605.
[10] Xu Nan, Tan Guangcai, Wang Hongyuan, et al. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure[J]. European Journal of Soil Biology, 2016, 74: 1-8.
[11] Wang Qiong, Garrity G M, Tiedje J M, et al. Na?ve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy[J]. Applied and Environmental Microbiology, 2007, 73(16): 5261-5267.
[12] Schloss P D, Gevers D, Westcott S L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies[J]. PLoS One, 2011, 6(12): e27310.
[13] Oberauner L, Zachow C, Lackner S, et al. The ignored diversity: complex bacterial communities in intensive care units revealed by 16S pyrosequencing[J]. Scientific Reports, 2013, 3: 1413.
[14] Rivas N M, Burton O T, Wise P, et al. A microbiota signature associated with experimental food allergy promotes allergic sensitization and anaphylaxis[J]. Journal of Allergy and Clinical Immunology, 2013, 131(1): 201-212.
[15] 李卓佳, 李奕雯, 曹煜成, 等. 凡納濱對蝦海水高位池養殖水體理化因子變化與營養狀況分析[J]. 農業環境科學學報, 2010, 29(10): 2025-2032.
Li Zhuojia, Li Yiwen, Cao Yucheng, et al. Analysis of physical-chemical factors variation and nutritional status of litopenaeus vannamei high level seawater ponds[J]. Journal of Agro-Environment Science, 2010, 29(10): 2025-2032.
[16] 張瑜斌, 章潔香, 詹曉燕, 等. 高位蝦池養殖過程主要理化因子的變化及水質評價[J]. 水產科學, 2009, 28(11): 628-634.
Zhang Yubin, Zhang Jiexiang, Zhan Xiaoyan, et al. Changes in main physicochemical factors in water and assessment of water quality during culture in higher-level shrimp ponds[J]. Fisheries Science, 2009, 28(11): 628-634.
[17] Sheik C S, Mitchell T W, Rizvi F Z, et al. Exposure of soil microbial communities to chromium and arsenic alters their diversity and structure[J]. PLoS One, 2012, 7(6): e40059.
[18] Tang Yuyi, Tao Peiying, Tan Jianguo, et al. Identification of bacterial community composition in freshwater aquaculture system farming ofLitopenaeusvannameireveals distinct temperature-driven patterns[J]. International Journal of Molecular Sciences, 2014, 15(8): 13663-13680.
[19] 李爽, 姜華, 張德民, 等. 光合細菌及其在水產養殖上的應用[J]. 中國水產, 2007(1): 81-82.
Li Shuang, Jiang Hua, Zhang Demin, et al. Photosynthetic bacteria and its application in aquaculture[J]. China Fisheries, 2007(1): 81-82.
[20] 王姣姣, 李丹, 宋堅, 等. 不同時期刺參養殖池塘海水菌群結構分析[J]. 中國農業科技導報, 2015, 17(2): 134-140.
Wang Jiaojiao, Li Dan, Song Jian, et al. Structural analysis of seawater bacterial community from culture pond of apostichopus japonicus at different periods[J]. Journal of Agricultural Science and Technology, 2015, 17(2): 134-140.
[21] Stenholm A R, Dalsgaard I, Middelboe M. Isolation and characterization of bacteriophages infecting the fish pathogenflavobacteriumpsychrophilum[J]. Applied and Environmental Microbiology, 2008, 74(13): 4070-4078.
[22] 姜海峰, 劉小林, 常亞青, 等. 皺紋盤鮑(Haliotisdiscushannai)腸道潛在益生菌的篩選及對幼鮑生長的影響[J]. 海洋與湖沼, 2013, 44(2): 482-487.
Jiang Haifeng, Liu Xiaolin, Chang Yaqing, et al. Selection of potential probiotics from abalone intestional and their effects on growth of juvenile small abaloneHaliotisdiscushannal[J]. Oceanologia et Limnologia Sinica, 2013, 44(2): 482-487.
[23] Romero M, Avendano-Herrera R, Magarios B, et al. Acylhomoserine lactone production and degradation by the fish pathogen Tenacibaculum maritimum, a member of the Cytophaga-Flavobacterium-Bacteroides (CFB) group[J]. FEMS Microbiology Letters, 2010, 304(2): 131-139.
[24] Ninawe A S, Selvin J. Probiotics in shrimp aquaculture: avenues and challenges[J]. Critical Reviews in Microbiology, 2009, 35(1): 43-66.
[25] Bruhn J B, Nielsen K F, Hjelm M, et al. Ecology, inhibitory activity, and morphogenesis of a marine antagonistic bacterium belonging to theRoseobacterclade[J]. Applied and Environmental Microbiology, 2005, 71(11): 7263-7270.
[26] Duchaud E, Boussaha M, Loux V, et al. Complete genome sequence of the fish pathogenFlavobacteriumpsychrophilum[J]. Nature Biotechnology, 2007, 25(7): 763-769.
[27] Khashe S, Janda J M. Biochemical and pathogenic properties ofShewanellaalgaandShewanellaputrefaciens[J]. Journal of Clinical Microbiology, 1998, 36(3): 783-787.
[28] Wang Liping, Chen Yanwu, Huang Hui, et al. Isolation and identification ofVibriocampbelliias a bacterial pathogen for luminous vibriosis ofLitopenaeusvannamei[J]. Aquaculture Research, 2015, 46(2): 395-404.
[29] Nimrat S, Suksawat S, Boonthai T, et al. PotentialBacillusprobiotics enhance bacterial numbers, water quality and growth during early development of white shrimp (Litopenaeusvannamei)[J]. Veterinary Microbiology, 2012, 159(3/4): 443-450.
[30] Tas S, Yilmaz I N, Okus E. Phytoplankton as an indicator of improving water quality in the golden horn estuary[J]. Estuaries and Coasts, 2009, 32(6): 1205-1224.
[31] Tian Chuan, Tan Jing, Wu Xin, et al. Spatiotemporal transition of bacterioplankton diversity in a large shallow hypertrophic freshwater lake, as determined by denaturing gradient gel electrophoresis[J]. Journal of Plankton Research, 2009, 31(8): 885-897.
The diversity changes of bacterial community in mariculture water ofLitopenasusvannameiat Zhangpu, Fujian Province
Hu Dong1,2, Wang Liping2, Zhao Ran1, Shao Zongze2
(1.SchoolofPublicHealth,XiamenUniversity,Xiamen361005,China; 2.KeyLaboratoryofMarineBiogeneticResources,ThirdInstituteofOceanograph,StateOceanicAdministration,Xiamen361005,China)
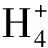
Litopenasusvannamei; microbial community; 16S rRNA gene high throughput sequencing; water quality parameters
10.3969/j.issn.0253-4193.2017.08.009
2017-01-18;
2017-02-22。
廈門南方海洋研究中心項目(14CZP034HJ08);深遠海(極地)微生物及其基因資源開發與產品應用示范 (201005032-1);國家自然科學基金項目(81673129)。
胡東(1990—),男,山東省新泰市人,主要研究方向為海洋環境微生物。E-mail:taianhudong@163.com
*通信作者:邵宗澤,男,研究員。E-mail:shaozz@163.com
S917.1
A
0253-4193(2017)08-0089-10
胡東,王麗萍,趙苒,等. 福建漳浦凡納濱對蝦海水養殖中后期水體細菌群落多樣性分析[J].海洋學報,2017,39(8):89—98,
Hu Dong, Wang Liping, Zhao Ran,et al. The diversity changes of bacterial community in mariculture water ofLitopenasusvannameiat Zhangpu, Fujian Province[J]. Haiyang Xuebao,2017,39(8):89—98, doi:10.3969/j.issn.0253-4193.2017.08.009
猜你喜歡
智慧少年·故事叮當(2020年10期)2020-11-06 06:19:00
中華詩詞(2020年1期)2020-09-21 09:24:52
河北理科教學研究(2020年1期)2020-07-24 08:14:34
作文成功之路·小學版(2020年11期)2020-02-01 06:26:58
作文周刊·小學二年級版(2018年29期)2018-11-26 11:20:28
小學生作文(中高年級適用)(2018年5期)2018-06-11 01:22:56
童話世界(2018年14期)2018-05-29 00:48:08
數學小靈通·3-4年級(2017年10期)2017-11-08 08:42:59
中學生數理化·七年級數學人教版(2017年11期)2017-04-23 07:18:00
數學大王·中高年級(2016年12期)2016-12-26 21:37:36
