溫度脅迫對三七光合特性及生理指標的影響
2017-05-30 10:48:04左應梅張金渝楊天梅楊紹兵楊維澤李紀潮楊美權許宗亮簡邦麗
南方農業學報 2017年12期
左應梅 張金渝 楊天梅 楊紹兵 楊維澤 李紀潮 楊美權 許宗亮 簡邦麗
摘要:【目的】研究三七對溫度脅迫的生理響應,揭示三七的耐寒和耐熱機制,為三七抗逆品種的選育、區域化種植和栽培管理提供理論依據。【方法】以盆栽生長第2年的三七植株為試驗材料,利用人工氣候培養箱進行5、10、25(對照)和35℃溫度處理,持續處理48 h后測定三七葉片的光合特性及葉綠素含量、保護酶活性等生理指標,并分析其變化規律。【結果】在5~25℃的溫度條件下,三七葉片的葉綠素a(Chla)、葉綠素b(Chlb)和總葉綠素[Chl(a+b)]含量呈先升后降的變化趨勢,與25℃處理相比,5℃處理分別降低21.91%、23.30%和22.78%,10℃處理分別提高51.69%、52.43%和51.96%,差異均達顯著水平(P<0.05,下同);與25℃處理相比,35℃高溫條件下,Chla、Chlb和Chl(a+b)含量及Chl(a/b)分別降低14.61%、6.80%、11.74%和8.10%,差異均達顯著水平。低溫脅迫下,三七葉片的最大凈光合速率(Pmax)、表觀量子效率(a)、光飽和點(LSP)、凈光合速率(Pn)、氣孔導度(G)和蒸騰速率(Tr)均顯著下降,胞間二氧化碳濃度(Ci)顯著升高,光補償點(LCP)呈先升后降的變化趨勢;高溫脅迫下,三七葉片的Pmax、LSP、Pn、Gs和Tr均顯著下降,Ci、a和LCP則呈上升趨勢。在低、高溫脅迫條件下,三七葉片的過氧化氫酶(CAT)活性顯著降低,過氧化物酶(POD)和超氧化物歧化酶(SOD)活性總體上顯著升高;丙二醛(MDA)含量變化不明顯。低溫脅迫下三七葉片的可溶性蛋白含量顯著升高。【結論】低、高溫脅迫均顯著抑制三七的光合能力。低溫脅迫下,三七主要通過提高POD和SOD活性,增加可溶性蛋白和葉綠素含量來增強其抗冷性;高溫脅迫下,三七主要通過提高POD和SOD活性來增強其抗熱性。
關鍵詞:三七;溫度脅迫;光合特性;生理特性
中圖分類號:S567.211 文獻標志碼:A 文章編號:2095-1191(2017)12-2145-07
0引言
【研究意義】三七[Panax notoginseng(Burk.)F.H.Chen]為五加科人參屬多年生藥用植物,是我國傳統常用名貴藥材,也是云南省最具特色的藥用生物資源。三七以其干燥后的根及莖葉入藥,具有抗炎、止血、抗衰老等藥理活性(王瑩等,2015)。三七對其生長的環境溫度要求嚴格,適宜種植在夏涼冬暖,熱量充沛,年溫差變化較小,年均溫15.8~19.3℃的低緯度區域。云南省地處低緯度高原山地,地形條件復雜多變,受季風氣候影響明顯,極端或異常天氣頻發,對當地三七的生長造成較大影響。此外,三七連作障礙嚴重,輪作需間隔10~20年,導致主產區的種植場地越來越少,三七種植帶逐漸北移,但北移后三七的生長易受冬春季極端低溫和持續低溫天氣的影響。同時,由于三七種植過程中病害發生較嚴重,常采用避雨栽培以減少病蟲害發生,但該種植模式易造成棚內溫度過高,對三七的生長發育、產量及品質產生不良影響。因此,深入研究溫度對三七光合特性、生理指標及抗逆性的影響,可為三七區域化種植、栽培管理和抗逆性品種的選育提供科學參考依據。【前人研究進展】低溫或高溫逆境將導致植物體內的活性氧代謝失調,發生氧化脅迫,引起細胞膜脂過氧化加劇、蛋白質變性及核苷酸損傷等,脅迫危害加劇可致使細胞死亡(Kuk et al.,2003),而植物可通過調節自身抗氧化系統對溫度脅迫作出適應性響應。同時,低溫或高溫脅迫可破壞葉片光合機構,使葉綠素合成受阻,光合電子傳遞鏈受限,影響碳同化所需能量,導致光合效率下降(武輝等,2014;李孟洋等,2015;許娟等,2016)。高溫脅迫方面,許桂芳和張朝陽(2009)研究表明,隨著高溫脅迫的加劇,4種珍珠菜屬植物葉片的質膜透性上升,超氧化物歧化酶(SOD)和過氧化物酶(POD)活性先升后降,葉綠素、可溶性蛋白和脯氨酸含量增加;任飛等(2012)研究表明,高溫脅迫處理24 h,日本無刺野薔薇和七姊妹葉片中的SOD活性比中國無刺野薔薇和普通月季顯著提高,4種供試薔薇植物葉片的POD活性均比對照(常溫)低;李孟洋等(2015)研究表明,隨著高溫脅迫時間的延長,不同產地茅蒼術葉片的凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(G)和蒸騰速率(Tr)均呈下降趨勢,且葉片SOD、POD和過氧化氫酶(CAT)活性及丙二醛(MDA)含量均隨脅迫時間的延長呈上升趨勢。低溫脅迫方面,武輝等(2014)研究表明,低溫脅迫下棉花幼苗葉片的Pn、Gs和Ci等指標均小幅度降低,脅迫解除后恢復正常,但隨著脅迫強度增加,各指標變化幅度顯著,脅迫后恢復能力降低,同時,低溫脅迫下葉片對光輻射的利用能力降低,且脅迫加強,葉片最大凈光合速率(Pmax)、表觀量子率(a)及光飽和點(LSP)急劇下降;許娟等(2016)研究表明,低溫脅迫后馬鈴薯葉片的CAT、POD和SOD活性及MDA含量上升,且可溶性糖、可溶性蛋白和脯氨酸含量整體均呈上升趨勢,但隨著脅迫時間的延長,各生理指標變化表現出不同趨勢;陳奇等(2017)研究白菜型油菜在低溫脅迫下葉片光合特性與內源激素的響應對其抗寒性的影響,結果表明,低溫脅迫均降低了Pn和Gs,且抗寒性弱的品種降幅較大。【本研究切入點】目前,關于溫度脅迫對三七光合特性及生理指標影響的研究報道極少。【擬解決的關鍵問題】以生長第2年的三七植株為試驗材料,研究持續低、高溫脅迫對三七葉片光合特性及生理指標的影響,揭示三七的耐寒和耐熱機制,為三七抗逆品種的選育及其區域化種植和栽培管理提供理論依據。
1材料與方法
1.1試驗材料
試驗用三七為1年生三七苗,由昆明市祿勸縣翠華鎮種植農戶提供,以生長第2年的三七植株為試驗材料。
試驗用土壤為石灰巖母質發育的山地紅壤,其基礎養分:有機質36.57 g/kg,全氮2.38 g/kg、全磷0.75 g/kg,全鉀14.15 g/kg,速效氮195.30 mg/kg、速效磷20.25 mg/kg、速效鉀272.80 mg/kg,pH 5.90~6.27。
1.2試驗方法
2014年1月15日~5月30日在云南省農業科學院藥用植物研究所小哨試驗基地進行試驗。1月15日,將大小一致、來源相同、無破損及病害的1年生三七苗,栽人到上口徑21 cm,下口徑14 cm,高19 cm的花盆中,基質為壤土:農家肥=2:1,每盆3.5kg,每盆種4株,農藝管理措施一致。
5月22日采用智能人工氣候箱(寧波江南儀器廠制造)進行試驗。設25(對照)、5、10和35℃共4個溫度處理,光照強度3500 lX,相對濕度60%,每處理3次重復;各溫度梯度持續處理48 h后,測定三七葉片的光合特性,并將測過光合特性的葉片立刻用蒸餾水沖洗干凈后于-80℃保存,進行各項生理指標測定。
1.3測定項目及方法
1.3.1光合特性指標測定 采用LI-6400便攜式光合測定系統(LI-COR,USA)測定光合一光強曲線(Pn-PAR)。在上午9:00~11:30進行測定,連續測定3 d,葉室溫度設為25℃,CO2濃度設為400 μmol/mol,光強(PAR)設為0、20、50、80、100、150、200、400、600、800、1200和1600μmol/m2·s,共12個梯度,由儀器配置的紅藍光源產生。依據Pn-PAR曲線的初始斜率(PAR<250μmol/m2·s)計算出a。參考Bassman和Zwier(1991)的方法計算Pmax、LSP及光補償點(LCP)等。此外測定各處理同一光強(PAR=200μmol/m2·s)下的Pn、Tr、Gs和Ci等參數。
1.3.2生理特性指標測定 SOD活性采用NBT(淡藍四唑)光還原法測定,以抑制NBT光還原反應50%所需酶量為一個酶活性單位(U),活性以u/gFW·min表示,SOD試劑盒購于南京生物工程研究所;POD活性采用愈刨木酚顯色法測定,以每分鐘OD470變化0.01為一個酶活性單位(U),活性以U/gFW·min表示;CAT活性采用紫外吸收法測定,以1 min內OD240降低0.01為一個酶活性單位(U),活性以U/gFW·min表示。MDA含量采用硫代巴比妥酸(TBA)比色法測定;可溶性蛋白含量采用考馬斯亮藍G-250染色法測定;葉綠素含量采用一定體積的95%酒精提取,用分光光度計測665、649和470 nm 3個波長的吸光度,根據高俊鳳(2006)的方法計算葉綠素a(Chla)、葉綠素b(Chlb)、總葉綠素[Chl(a+b)]含量及葉綠素a和葉綠素b的比值[Chl(a/b)]。
1.4統計分析
采用Excel 2007進行數據整理和制圖,利用JMP7進行差異顯著性分析。
2結果與分析
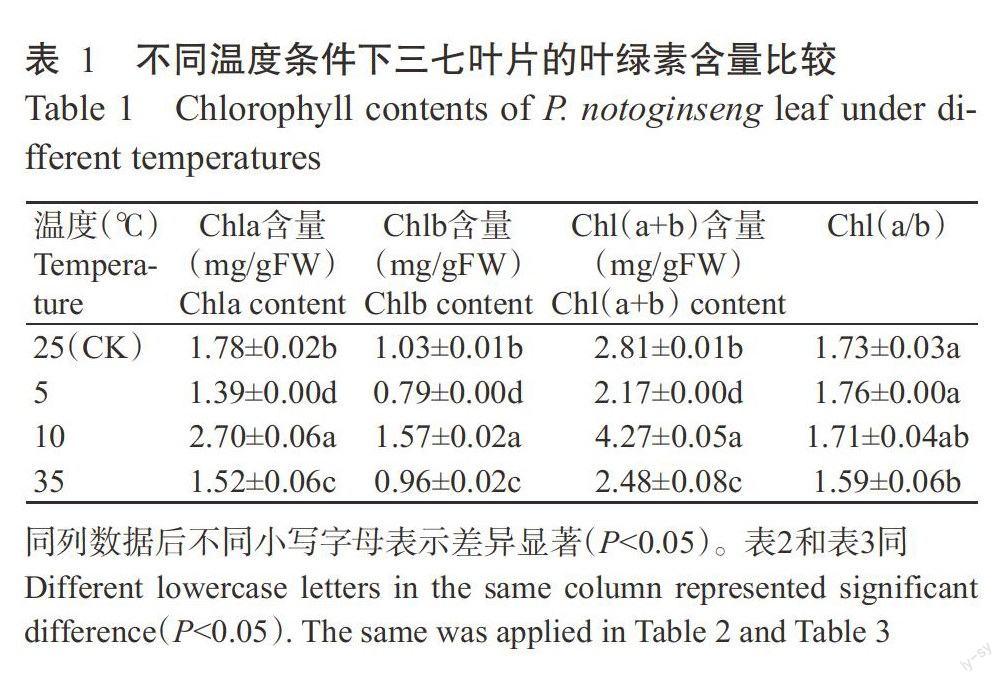
2.1不同溫度條件對三七葉片葉綠素含量的影響
由表1可知,在5~25℃的溫度條件下,三七葉片的Chla、Chlb和Chl(a+b)含量呈先升后降的變化趨勢。與25℃處理的Chla、Chlb和Chl(a+b)含量相比,5℃處理分別降低21.91%、23.30%和22.78%,10℃處理分別提高51.69%、52.43%和51.96%,差異均達顯著水平(P<0.05,下同),而Chl(a/b)變化不顯著(P>0.05,下同)。在35℃高溫條件下,Chla、Chlb、Chl(a+b)含量和Chl(a/b)分別比25℃處理降低14.61%、6.80%、11.74%和8.10%,差異顯著。表明三七葉片的葉綠素含量與其耐寒性有關,在一定的低溫脅迫下三七可通過增加其葉片葉綠素含量以提高光合作用,超過一定低溫脅迫后,葉片葉綠素合成受阻,而高溫脅迫下其葉綠素合成同樣受阻。
2.2不同溫度條件對三七葉片光合特性的影響
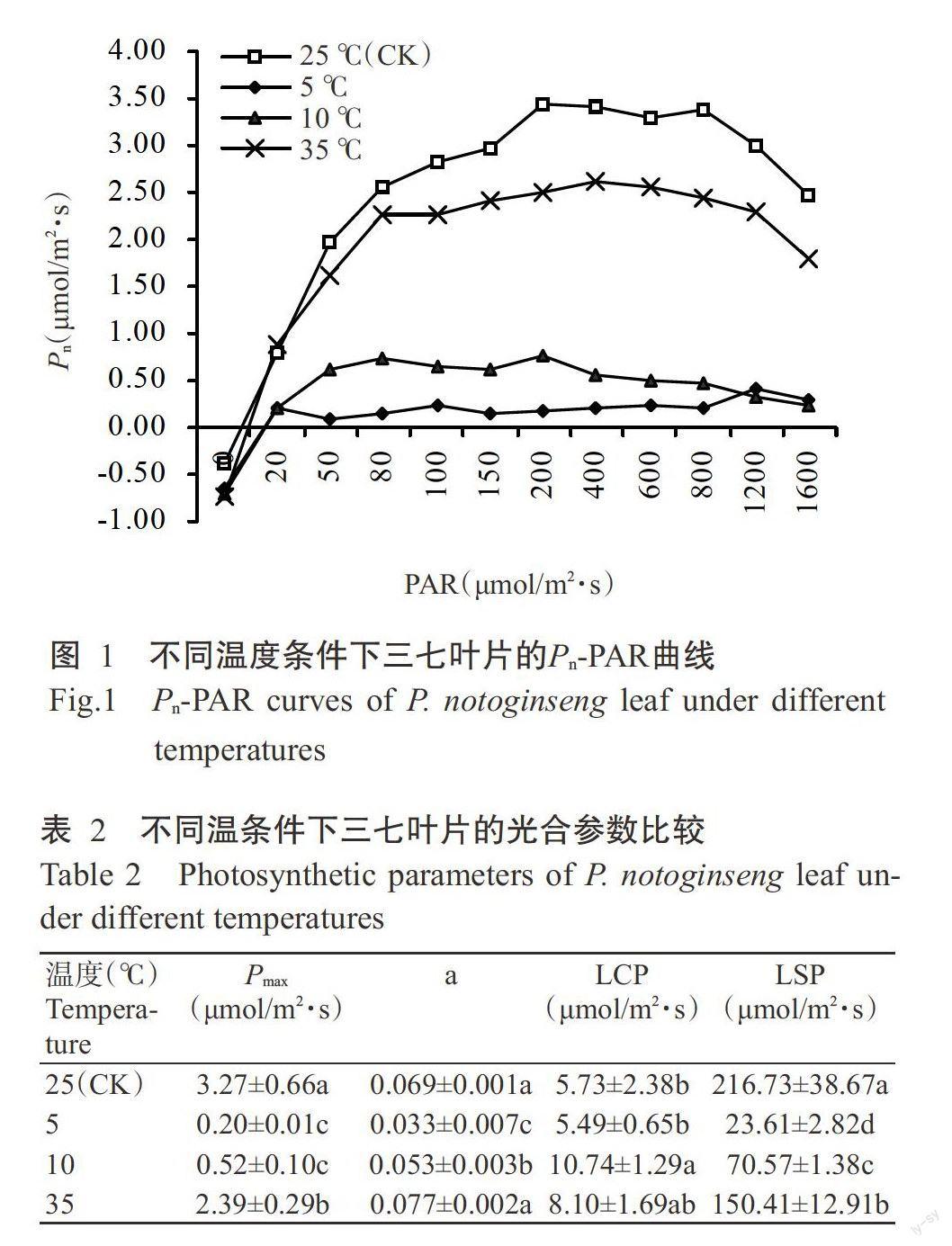
2.2.1光響應曲線和光合參數比較 由圖1可看出,不同溫度條件下,三七葉片的Pn隨PAR增強其變化趨勢并不一致。10和35℃處理下的Pn變化趨勢與25℃基本一致,均隨PAR的增強先增加后下降,且PAR相同時,10和35℃處理下的Pn均低于25℃對應值。5℃處理下的Pn隨PAR增強變化不明顯,表明在該溫度條件下三七的光合作用嚴重受阻。Pn-PAR曲線方程及相應方程的計算結果(表2)表明,溫度脅迫導致三七葉片的Pmax、a、LCP和LSP呈不同程度的變化。低、高溫脅迫均使三七葉片的Pmax和LSP顯著下降,與25℃處理相比,5、10和35℃處理的Pmax分別下降93.88%、84.10%和26.91%,LSP分別下降89.11%、67.44%和30.60%。在低溫脅迫下,a呈顯著下降趨勢,即與25℃處理相比,5和10℃處理的a分別下降52.17%和23.19%。5℃處理的LCP下降4.19%,與25℃處理差異不顯著,10℃處理的LCP則顯著升高87.43%。在35℃高溫條件下,a和LCP的變化均不顯著。說明高、低溫脅迫均抑制了三七的光合作用,在一定條件下低溫脅迫的傷害程度較高溫嚴重;三七在強光下均出現不同程度的光抑制,低、高溫脅迫越嚴重,越容易出現光抑制現象。
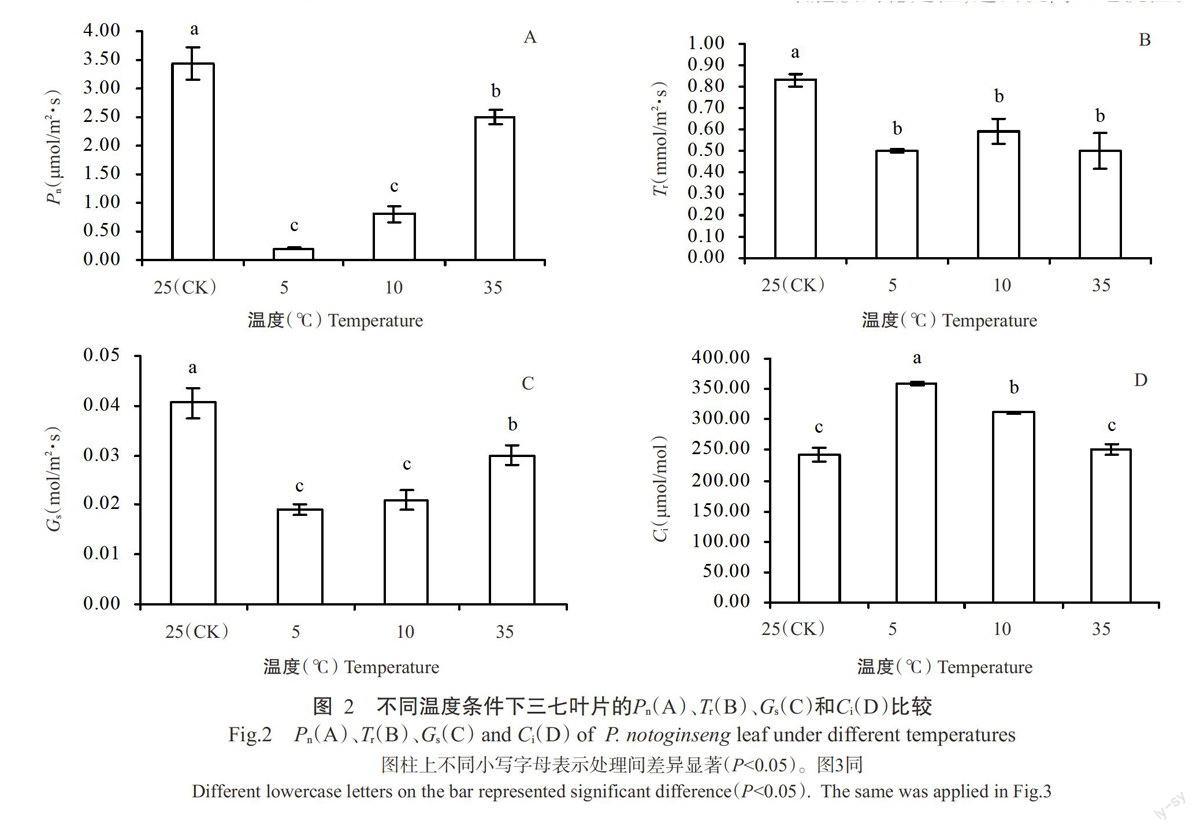
2.2.2 Pn、Tr、Gs和Ci的比較在200μmol/m2·s光照強度下,低、高溫脅迫使三七葉片的Pn(圖2-A)、Tr(圖2-B)和Gs(圖2-C)均顯著下降,與25℃處理相比,5、10和35℃處理的Pn分別下降94.19%、76.74%和27.33%,Tr分別下降39.76%、28.92%和39.76%,Gs分別下降53.09%、48.15%和25.93%。G(圖2-D)則呈相反的變化趨勢,與25℃處理相比,5和10℃處理的Ci分別顯著升高47.74%和27.98%,35℃處理的G升高3.29%,變化不顯著。
2.3不同溫度條件對三七葉片保護酶活性和MDA含量的影響
由表3可知,低溫和高溫脅迫下,三七葉片的CAT活性降低,SOD活性升高。與25℃處理相比,5、10和35℃處理的CAT活性分別顯著降低60.71%、46.83%和68.15%,SOD活性分別顯著升高18.49%、107.54%和22.36%。5和35℃處理的POD活性較25℃處理顯著升高,10℃處理的POD活性較25℃處理略有降低,但差異不顯著。低溫或高溫脅迫下,三七葉片的MDA含量均較25℃處理有所增加,但差異不顯著。說明三七的保護酶系統對溫度脅迫較敏感,可通過增強其葉片的POD和SOD活性,及時清除活性氧,保護細胞膜的穩定性,進而提高三七抗性。
2.4不同溫度條件對三七葉片可溶性蛋白含量的影響
由圖3可看出,不同溫度條件下,三七葉片的可溶性蛋白含量均呈不同程度的升高趨勢。與25℃處理相比,5和10℃處理的可溶性蛋白含量顯著升高22.72%和32.77%;35℃處理的可溶性蛋白含量也有所升高,但與25℃處理差異不顯著。說明三七葉片的可溶性蛋白含量與三七的耐低溫能力密切相關。
3討論
3.1不同溫度條件對三七葉片葉綠素含量的影響
葉綠素是植物細胞內參與光合作用的重要色素,起著接收光能和轉換能量的作用,其含量與光合能力密切相關。目前,關于逆境脅迫下葉綠素含量變化的研究結果尚存在一定差異。任彩虹等(2007)研究指出,高溫脅迫下,馬鈴薯幼苗葉片的葉綠素含量明顯下降。金花等(2011)分析比較了7種常綠闊葉植物越冬期間葉綠素含量的變化,結果顯示,葉綠素含量隨低溫脅迫的加劇先降后升。邵怡若等(2013)研究表明,隨低溫脅迫時間的延長,假連翹幼苗的葉綠素含量先升后降,鹽膚木、老鴨嘴和葛藤幼苗的葉綠素含量則表現為持續下降,葉綠素含量的變化趨勢與植物類型和脅迫時間密切相關。本研究中,在5-25℃的溫度條件下,三七葉片的Chla、Chlb和Chl(a+b)含量呈先升后降的變化趨勢;在35℃高溫條件下,Chla、Chlb、Chl(a+b)含量和Chl(a/b)較25℃處理顯著下降。表明在一定的低溫范圍內,三七可通過增加其葉片葉綠素含量以維持或提高其光合作用,隨著低溫脅迫的加劇,三七葉片的葉綠素合成受阻;高溫脅迫也可導致三七葉片葉綠素合成受阻。低、高溫脅迫使得葉片葉綠素含量明顯下降。這是由于葉綠素合成是在葉綠體或前質體中一系列酶的作用下所形成,低、高溫脅迫可能通過影響葉綠素合成相關的酶類,如葉綠素合成酶等,進而抑制葉片的葉綠素合成(Tewan and Tripathy,1998);低、高溫脅迫引起活性氧積累,加速葉綠素降解,造成葉片葉綠素含量降低(Willekens et al.,1994)。
3.2不同溫度條件對三七葉片光合特性的影響
光合作用是植物體的重要代謝之一,可作為判斷植物生長狀況和抗逆性強弱的重要指標,光補償點和光飽和點的高低直接反映植物對光環境的適應能力(劉悅秋等,2007)。低溫脅迫導致三七葉片對光輻射的利用能力下降,隨著脅迫溫度降低,葉片的Pmax、a和LSP快速下降,LCP則呈先升后降的變化趨勢,與武輝等(2014)在棉花幼苗上的研究結果相似。此外,本研究結果顯示,低溫脅迫下Pn、Gs和Tr呈顯著下降趨勢,而Cli呈顯著上升趨勢,與邵怡若等(2013)在假連翹幼苗上的研究結果相似。據Farqu-har和Sharkey(1982)報道,當Pn和Gi變化方向一致,而Gs下降時,認為Pa的下降主要是由Gs引起。本研究中,隨著三七葉片Gs顯著下降,Ci則顯著上升,說明Pn下降是由非氣孔限制占主導因素。由此可知,低溫脅迫使三七葉片組織受損,導致光合機構的穩定性能降低、暗反應過程所需的ATP和NADPH供應不足、關鍵酶活性下降、反應底物的再生速率減慢,對CO2的固定造成阻礙,引起細胞間隙CO2積累,從而導致葉肉細胞光合性能降低(Yu et al.,2002)。在35℃高溫脅迫下,三七葉片的Rmax和LSP顯著下降,a和LCP則呈上升趨勢。說明高溫脅迫同樣可導致三七葉片對強光環境的適應能力降低。同時,高溫條件下,三七葉片的Pn、Gs和Tr均呈顯著下降趨勢,G呈上升趨勢。與杜堯東等(2012)研究得出的高溫處理后水稻劍葉的Pn、Gs下降,Ci上升的結果相似。
3.3不同溫度條件對三七葉片保護酶活性和膜質過氧化水平的影響
保護酶是植物適應逆境的重要生理基礎(馬成倉等,2004),其活性高低可反映植物對環境條件適應能力的強弱。保護酶系統主要包括CAT、POD和SOD,其協同作用可降低逆境不良信號對細胞膜脂的攻擊(Jebara et al.,2005)。本研究中,低溫和高溫脅迫下,CAT活性顯著降低,POD和SOD活性總體上顯著升高。表明三七的保護酶系統對溫度脅迫的反應十分敏感,一定程度的低溫或高溫脅迫下,三七可通過增強其葉片POD和SOD活性,及時清除因脅迫而積累的H2O2及活性氧,以保持細胞內活性氧產生和清除的動態平衡,保護細胞膜穩定性,進而提高其對低、高溫逆境的抗性。這可能是三七葉片對低、高溫脅迫的一種應激性保護方式,類似結果也表現在其他植物對溫度脅迫后的應激反應中(陳穎等,2012)。POD和SOD可能是三七主要的耐冷、耐熱保護酶系統。逆境脅迫下產生的活性氧若不能及時清除,可造成自由基積累,損害細胞膜系統,引起或加劇膜質過氧化作用,產生MDA破壞細胞內活性氧的代謝平衡(Chen,1991)。本研究結果表明,一定程度的低溫或高溫脅迫下,三七葉片的MDA含量變化不顯著,說明三七具有一定的耐冷和耐熱性能,可能與其具有的較高POD和SOD活性有關。
3.4不同溫度條件對三七葉片可溶性蛋白含量的影響
低溫引起植物細胞中蛋白質發生變化,主要表現為可溶性蛋白和酶類的變化及激發信號產生抗寒性蛋白(王毅等,1994)。可溶性蛋白能增加細胞的持水力,并具有較強的親水性,可通過束縛細胞周圍的水分緩解低溫條件下原生質因結冰而受到傷害,提高植物體抗寒性(沙偉等,2008)。本研究結果表明,低溫脅迫下三七葉片內可溶性蛋白含量顯著升高。表明低溫可誘導產生新的蛋白,增加可溶性蛋白含量,有利于提高三七的抗寒性,與朱政等(2011)在茶樹葉片上的研究結果相似。高溫條件會造成正常蛋白合成受限、蛋白分解加劇、細胞內可溶性蛋白含量下降,因此,高溫脅迫會引起植物體內可溶性蛋白含量下降(Chaitanya et al.,2001)。但本研究中高溫脅迫下三七葉片的可溶性蛋白含量有小幅度上升。說明三七具有一定的耐熱性,與魯歧等(1992)研究的三七起源于第三紀,屬古熱帶殘遺植物的結論吻合。
4結論
低溫或高溫脅迫對三七葉片光合特性及生理特性的影響明顯。低、高溫脅迫均顯著抑制三七的光合能力,與三七“喜冬暖夏涼,畏嚴寒酷熱”的生長習性相符。低溫脅迫下,三七主要通過提高其葉片POD和SOD活性,增加可溶性蛋白和葉綠素含量來增強其抗冷性,從而緩解低溫對其造成的傷害;高溫脅迫下,三七主要通過提高POD和SOD活性來增強其抗熱性,從而緩解高溫對其所造成的傷害。
