水稻黃嘌呤脫氫酶基因OsXDH克隆及表達分析
2017-05-30 10:48:04
南方農業學報 2017年12期
關鍵詞:水稻

摘要:【目的】克隆水稻黃嘌呤脫氫酶(Xanthine dehydrogenase,XDH)基因(OsXDH),分析其生物信息學特性及表達特性,為研究XDH在水稻生長發育和響應逆境脅迫中的調控機制提供理論依據。【方法】以粳稻品種日本晴為材料,采用同源克隆技術克隆OsXDH基因,應用生物信息學方法對其氨基酸序列進行分析。利用實時熒光定量PCR(qPCR)檢測OsXD14基因的組織表達特性及逆境脅迫下的表達情況,并對不同轉基因株系乳熟期劍葉OsXDH基因表達量、XDH活性和葉綠素含量進行比較分析。【結果】克隆獲得OsXDH基因的開放閱讀框序列(ORF)(GenBankg登錄號LOC4333171),其長度為4110 bp,編碼1369個氨基酸。OsXDH蛋白分子量大小為150.23 kD,理論等電點(pI)為6.54,與小麥、高粱、玉米、谷子和油菜等作物XDH蛋白氨基酸序列的相似性分別為84.54%、84.07%、81.52%、76.35%和69.22%,表明XDH蛋白氨基酸序列具有高度保守性。OsXDH基因在水稻不同組織部位均有表達,灌漿期的表達量顯著高于苗期和分蘗盛期(P<0.05),且受干旱、黑暗、高溫和鹽脅迫誘導高效表達。OsXDH過表達水稻轉基因株系乳熟期劍葉的XDH活性和葉綠素含量高于野生型,OsXDH干擾轉基因株系的XDH活性和葉綠素含量低于野生型。【結論】OsXDH基因受水稻生長發育和逆境脅迫因子誘導表達,推測其是調控水稻生長發育和響應逆境脅迫的關鍵基因。
關鍵詞:水稻;OsXDH基因;基因克隆;生物信息學;表達分析;脅迫
中圖分類號:S511.03 文獻標志碼:A 文章編號:2095-1191(2017)12-2113-09
0引言
【研究意義】由于生態環境不斷惡化,干旱、鹽堿、極端溫度及重金屬污染等嚴重影響糧食作物的產量和品質。作物抗性是由多基因控制的數量性狀,抗性基因克隆和功能分析已成為研究熱點。黃嘌呤脫氫酶(Xanthine dehvdrogenase,XDH)作為調控嘌呤代謝的中間酶類,能將黃嘌呤和次黃嘌呤催化生成尿酸,經一系列代謝反應,最終生成酰脲類物質(Ureides),在應答生物脅迫與非生物脅迫中發揮重要作用(Montalbini,2000;Aguey-Zinsou et al.,2003;Zdunek-zastocka and Lips,2003;Taylor and Cowan,2004;Zrenner et al.,2006;Brychkova et al.,2008a)。因此,克隆水稻XDH基因(OsXDH),分析其序列特征,可為闡明植物XDH的抗逆境脅迫調控機制提供理論參考。【前人研究進展】目前,已對不同植物XDH的抗逆境脅迫調控機制開展了大量研究,結果均表明XDH對逆境脅迫有應答響應,與植物體內的多個生理代謝相關(Aguey-Zinsou et al.,2003;Taylorand Cowan,2004;Nakagawa et al.,2007),如Montal-bini(1991)研究發現,感染銹病的豆科植物葉片中嘌呤分解代謝加快,酰脲類物質含量快速積累,對非共生病原的抗性增強。植物抗性增強的原因是嘌呤代謝產物酰脲類物質具有較高的碳氮比,易于在植物體內轉運,是氮素貯存和低耗轉運載體,可降低氮素轉運過程中的能量損耗,以提高植物在逆境脅迫下的生存能力(Buchanan-Wollaston,1997;Gepstein,2004),因此,植物體內的XDH活性隨著鹽和NH4濃度的增加而升高,且氨態氮處理下XDH活性高于硝態氮處理(Barabfis et al.,2000;Zrenner et al.,2006;Zdunek-zastocka and Lips,2003)。也有研究表明,酰脲類物質可清除植物體內H2O2、O2等活性氧自由基(Becker et al.,1989;Pastori and del Rio,1997),增加尿囊素和尿囊酸含量,從而降低氧化性損傷和減少幼苗死亡(Brychkova et al.,2008b)。當植物受干旱、鹽脅迫和黑暗脅迫時,XDH活性升高,尿酸和酰脲類物質含量增加,體內活性氧含量降低,促使植株對環境脅迫的耐受能力增強,其病死率明顯降低(Brychkova et al.,2008b;Watanabe et al.,2010;Maet al.,2016;Hofinann,2016;You et al.,2017)。但鉬素缺失會阻礙黃嘌呤分解代謝,不利于氮素在植株體內的運輸,導致植株長勢變弱(孫學成和胡承孝,2005)。此外,有研究表明XDH蛋白結構的發生改變可介導細胞分裂素、脫落酸和生長素等植物激素的動態平衡,發揮對植物性狀及抗逆境脅迫的調控作用(Leydecker et al.,1995;Taylor and Cowan,2001;Smith and Atkins,2004;Watanabe et al.,2014)。【本研究切入點】至今鮮見有關水稻OsXDH基因克隆并分析其在非生物脅迫下表達特性的研究報道。【擬解決的關鍵問題】利用同源克隆技術克隆水稻OsXDH基因的開放閱讀框序列(ORF),采用生物信息學軟件分析其序列結構特征,并用實時熒光定量PCR(qPCR)檢測其組織表達特性及逆境脅迫下的表達情況,同時比較不同OsXDH轉基因水稻株系乳熟期劍葉OsXDH基因表達量、XDH活性及葉綠素含量的差異,為研究XDH對水稻品種生長發育及響應逆境脅迫的調控機理提供理論參考。
1材料與方法
1.1試驗材料
供試的粳稻品種為日本晴(Oryza sativa L.spp.japonica cv.Nipponbare),其野生型、OsXDH干擾轉基因株系(Ri3、Ri5和Ri6)及過表達轉基因株系(OEl、OE3和OE9)均由江西省作物生理生態與遺傳育種重點實驗室提供。主要試劑:KOD FX高保真PCR酶購自東洋紡(上海)生物科技有限公司,Mini-BEST Plant RNA Extraction Kit試劑盒、PrimeScriptTMRT Master Mix試劑盒和SYBR Premix Ex TaqTMⅡ試劑盒購自寶生物工程(大連)有限公司,其他生化試劑均購自生工生物工程(上海)股份有限公司。主要設備儀器:人工氣候箱(GXZ智能型,寧波江南儀器廠)、ChemiDoc XRS+凝膠成像系統(BIO-RAD,美國)、NanoDrop ND2000超微量核酸蛋白分析儀(Thermo,美國)、CFX96 Real-Time PCR Detection System(B10-RAD,美國)、TU-1810紫外可見分光光度計(普析通用,北京)和電泳轉印系統(BIO-RAD,美國)。
1.2脅迫處理及樣品采集
干旱脅迫處理采用土培法,高溫、黑暗和鹽脅迫處理均采用水培法。所有處理均在人工氣候箱中進行,正常生長條件參數:12 h/12 h(白天/黑夜)、28℃/25℃(白天/黑夜)、光照強度為20000 lx/0 lX(白天/黑夜)和75%相對濕度。水稻幼苗在正常條件下生長N4.5葉期,選取長勢基本一致的植株進行脅迫處理。干旱脅迫處理:施用30%PEG-6000溶液進行模擬干旱脅迫處理,脅迫時間為4 d。高溫脅迫處理:處理期間將人工氣候箱溫度調至42℃/42℃(白天/黑夜),其他參數不變,脅迫時間為5 d。黑暗脅迫處理:處理期間將人工氣候箱光照調至0 lx/0 lX(白天/黑夜),其他參數不變,脅迫時間為6 d;鹽脅迫處理:在營養液中添加200 mmol/L NaCl,脅迫時間為3 d,脅迫處理后,更換為正常的水培營養液。每處理均在脅迫處理前、脅迫處理后及恢復生長7 d后采集主莖最上部完全展開葉,液氮速凍后,于80℃保存,用于不同逆境脅迫下OsXDH基因的表達特性分析。
將日本晴的野生型、OsXDHtz擾轉基因株系及過表達轉基因株系種植于江西農業大學實驗田,采集野生型苗期(4.5葉期)的葉(主莖最上部葉)、莖、根及分蘗盛期和灌漿期主莖最上部葉,液氮速凍后,于-80℃保存,用于qPCR檢測不同組織及不同生育期OsXDH~因的表達情況;采集野生型、OsXDH干擾轉基因株系及過表達轉基因株系乳熟期劍葉,液氮速凍后,于-80℃保存,用于OsXDH基因表達量、XDH活性和葉綠素含量測定。
1.3總RNA提取及cDNA第一鏈合成
按照RNAprep Pure植物總RNA提取試劑盒說明提取水稻葉片總RNA,利用NanoDrop ND2000超微量核酸蛋白分析儀對其濃度和質量進行檢測,利用1%瓊脂糖凝膠電泳檢測其完整性。以提取的總RNA為模板,按照PrimeScriptTM1stStand cDNA Syn-thesis Kit說明反轉錄合成cDNA第一鏈。
1.4引物設計及合成
根據擬南芥AtXDH1基因序列在NCBI數據庫中進行BLAST檢索,發現在水稻基因組數據庫中存在相似度較高的轉錄本(OSJNBa0091822.11),被詮釋為Xanthine dehydrogenase。根據NCBI數據庫中AtXDH1基因的cDNA序列,利用Primer 5.0設計引物(表1),引物由生工生物工程(上海)股份有限公司合成。
1.5基因克隆
以cDNA為模板進行PCR擴增,反應體系50.0μL:2xPCR Buffer for KOD FX 25.0μL,2 mmol/LdNTPs 10.0μL,10μmol/L正、反向引物(ORF-F和ORF-R)各1.5μL,cDNA模板2.0μL,1.0 U/μL KODFX 1.0μL,ddH2O補足N50.0μL。擴增程序:94℃預變性2.0 min;98℃10 s,58℃30 s,68℃5 min,進行35個循環;68℃延伸10.0 min,4℃保存。取10.0μL PCR產物用于1%瓊脂糖凝膠電泳檢測,其余送至生工生物工程(上海)股份有限公司測序。
1.6生物信息學分析
利用NCBI數據庫進行BLAST同源比對分析,利用ORF Finder尋找基因序列的編碼框,利用Prot-Param和Smart進行蛋白的理論性質和保守結構域預測,利用NetPhos 3.1 Server和DictyOGlvc 1.1 Server預測氨基酸序列磷酸化及O-糖基化修飾情況,利用ProtScale分析蛋白質的疏水性,利用SignalP1.1Server、TMHMM Sever 2.0和PSORT II預測蛋白的信號肽和亞細胞定位預測,運用SOPMA預測蛋白的二級結構,使用DNAMAN進行氨基酸序列比對及聚類分析。
1.7 qPCR檢測
利用SYBR*Premix Ex TaqTMⅡ試劑盒在CFX96Real-Time PCR Detection System上進行qPCR檢測,內參基因為Ubq-real,其引物序列(Ubq-real-F和Ubq-real-R)見表1。反應體系25.0μL:SYBR Pre-mix Ex Taq Ⅱ 12.5μL,正、反向引物(qPCR-F和qPCR-R)各1.0μL,DNA模板(<100 ng)2.0μL,ddI-120補足至25.0μL。擴增程序:95℃預變性30 s;95℃5 s,60℃30 s,進行40個循環。利用Bio-Rad CFX Manager 2.1數據分析模塊分析OsXDH基因的表達豐度。
1.8 XDH活性測定
稱取0.2 g去除葉脈的水稻劍葉碎片,液氮研磨2~3次后加入800.0μL蛋白提取液研磨至糊狀;將研磨液移入2 mL離心管,4℃下14000 r/min離心20 min后取上清液,65℃水浴90 s變性,再離心取上清液;取10μL上清液放入5 mL離心管中,加入490.0μLddH2O和4.5 mL考馬斯亮藍溶液混勻,于595 nm波長下測定OD值。利用牛血清蛋白制作標準曲線,測定可溶性蛋白含量,并進行非變性聚丙烯酰胺凝膠電泳,上樣量為50 ug,時長為4 h,將膠片放至以次黃嘌呤為底物和以氮藍四唑為顯色劑的混合溶液中,暗處20 min,用冰醋酸清洗后成像(Sagiet al.,1998)。
1.9葉綠素含量測定
參照Lichtenthaler和Wellbuen(1983)的方法,用80%丙酮溶液提取水稻葉片葉綠素,并用分光光度法測定其含量。
1.10統計分析
試驗數據采用Excel 2007進行整理作圖;利用SPSS 22.0進行顯著性分析(One-way ANOVA)。
2結果與分析
2.1 OsXDH基因cDNA編碼區的克隆結果
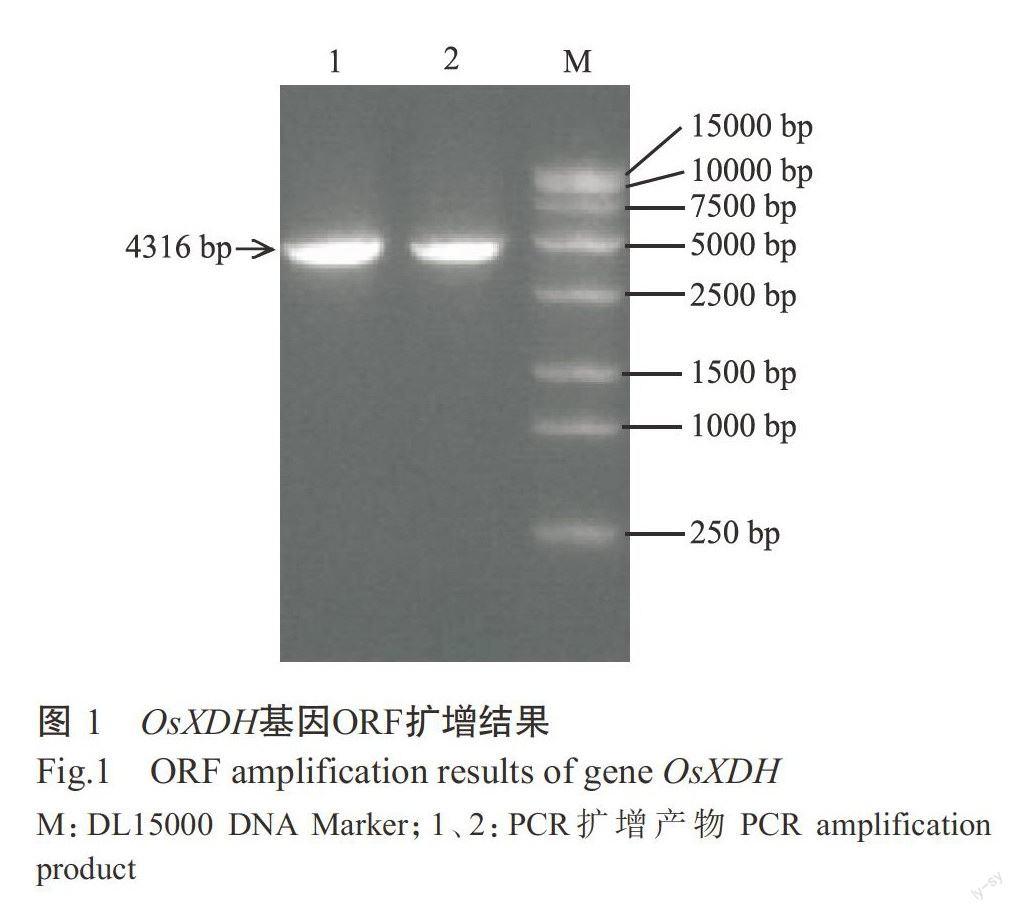
以反轉錄合成的cDNA為模板,以CDS-F和CDS-R為引物,PCR擴增獲得目的片段。結果如圖1所示,目的片段約4500 bp,與預期結果相符。測序結果顯示,克隆獲得的OsXDH基因序列長度為4316bp,其開放閱讀框(ORF)長度為4110 bp,編碼1369個氨基酸(圖2)。將OsXDH基因序列提交至Gen-Bank,登錄號LOC4333171。OsXDH基因與基因組DNA序列的比對結果顯示,該基因含有14個外顯子和13個內含子。
2.2 0sXDH蛋白的生物信息學分析結果
2.2.1理化性質 OsXDH蛋白的理化性質預測結果顯示,其分子式為C6713H10509N182501976857,分子量為150.23 kD,理論等電點(pI)為6.54;丙氨酸、甘氨酸、亮氨酸和纈氨酸個數分別占0sXDH蛋白氨基酸總數的8.0%、8.2%、8.6%和7.7%,不含蛋氨酸和含硒半胱氨酸;不穩定指數為38.87,表明其為穩定蛋白。
2.2.2保守結構域 OsXDH蛋白不僅存在1個CO_deh_flav_C結構域(455~561 aa),可與鉬結合形成鉬輔因子,構成XDH底物的結合部位和氧化還原部位,還存在1個進化保守的蛋白結構域Aid Xandh C(620~727 aa),此結構域僅存在于醛氧化酶和黃嘌呤脫氫酶中,由此推測黃嘌呤脫氫酶與醛氧化酶具有相似的功能,均可參與植物對非生物脅迫的響應。
2.2.3蛋白磷酸化和糖基化位點 氨基酸殘基的磷酸化和糖基化可調節蛋白的活性和功能。0sXDH蛋白氨基酸序列中發生磷酸化修飾的位點有131個,其中色氨酸、蘇氨酸和酪氨酸磷酸化修飾位點分別有71、45和15個(圖3);發生O-糖基化修飾的位點僅1個(圖4)。通過預測磷酸化與O-糖基化修飾位點,為黃嘌呤脫氫酶翻譯后修飾調控研究提供參考。
2.2.4信號肽及亞細胞定位 信號肽預測結果顯示,OsXDH蛋白信號肽平均值為0.106(<0.500),推測其為非分泌蛋白,在細胞質中合成后不被轉運。利用TargetP 1.1 Server預測發現OsXDH蛋白廣泛位于細胞內(除葉綠體和線粒體外),而pSORTⅡ預測其位于葉綠體中,存在明顯分歧,因此,其實際位置需利用免疫熒光、GFP融合表達等技術進一步確認。
2.2.5二級結構 OsXDH蛋白的二級結構中,無規則卷曲(Random coil,C)占35.43%,α-螺旋(α-he-liX,H)占31.56%,延伸鏈(Extended strand,E)占22.86%,β-轉角(β-turn,t)占10.15%。如圖5所示,4種二級結構分布較均勻。
2.2.6 親/疏水性 蛋白的親/疏水性有助于提高其穩定性及多樣性,與其結構和功能密切相關。由圖6可知,0sXDH蛋白疏水性最強的位點為第1314位,分值2.389;親水性最強的位點為第1029位,分值-3.589,親水性平均值為-0.098,由此推測0sXDH為親水性蛋白。
2.2.7同源比對及系統發育進化樹分析 從NCBI數據庫篩選獲得5種作物的XDH蛋白氨基酸序列,并與OsXDH蛋白的氨基酸序列進行比對,結果顯示OsXDH蛋白的氨基酸序列與高粱(Sorghum bico-lot)、谷子(Setaria italica)、小麥(Aegilops tauschii)、玉米(Zea mays)和油菜(Brassica napus)XDH氨基酸序列相似性分別為84.54%、84.07%、81.52%、76.35%和69.22%(圖7)。系統發育進化樹分析結果表明,單子葉植物和雙子葉植物XDH蛋白同系物分別聚在兩個不同的分支,表明XDH蛋白在單子葉植物和雙子葉植物間存在一定的分化,但XDH蛋白在同種屬間保守性較高,如水稻與小麥、二穗短柄草(Brachypodium distachyon)、谷子和高粱同屬一個小分支,均為禾本科植物,XDH蛋白具有較高的同源性;雖然玉米同屬禾本科植物,但與蘆筍、油棕、芭蕉和蝴蝶蘭分屬于單子葉植物的另一分支群(圖8),其原因可能與物種起源地環境有關,玉米、蘆筍、油棕、芭蕉和蝴蝶蘭均起源于熱帶或亞熱帶地區。綜上所述,植物中XDH蛋白的氨基酸序列具有高度保守性,水稻OsXDH蛋白與小麥、二穗短柄草的XDH蛋白分子進化距離最小,親緣關系最近。
2.3 OsXDH基因表達特性分析結果
2.3.1 OsXDH基因組織表達特性 從圖9-A可知,苗期OsXDH基因在水稻根、莖和葉(主莖最上部葉)中的表達量無顯著差異(P<0.05,下同)。從圖9-B可知,苗期和分蘗盛期OsXDH基因在水稻主莖最上部葉片的表達量無顯著差異,但灌漿期OsXDH基因的表達量顯著高于苗期和分蘗盛期,由此推測OsXDH基因為特異性表達,參與水稻生長發育相關的分子調控。
2.3.2不同脅迫下OsXDH基因的表達情況 由圖10可知,不同脅迫處理均可誘導水稻OsXDH基因上調表達,其中干旱、黑暗和高溫脅迫處理后OsXDH基因的表達量均顯著高于脅迫處理前,恢復生長7 d后其表達量顯著降低,但仍顯著高于脅迫處理前,尤其是干旱和高溫脅迫處理后OsXDH基因的表達量增至脅迫處理前的2.5倍。鹽脅迫處理后OsXDH基因的表達量顯著高于脅迫處理前,恢復生長7 d后其表達量顯著降低,但與脅迫處理前無顯著差異。可見,OsXDH基因的表達受到逆境脅迫因子誘導高效表達,參與逆境脅迫響應。
2.4 OsXDH基因表達量對水稻乳熟期生理指標的影響
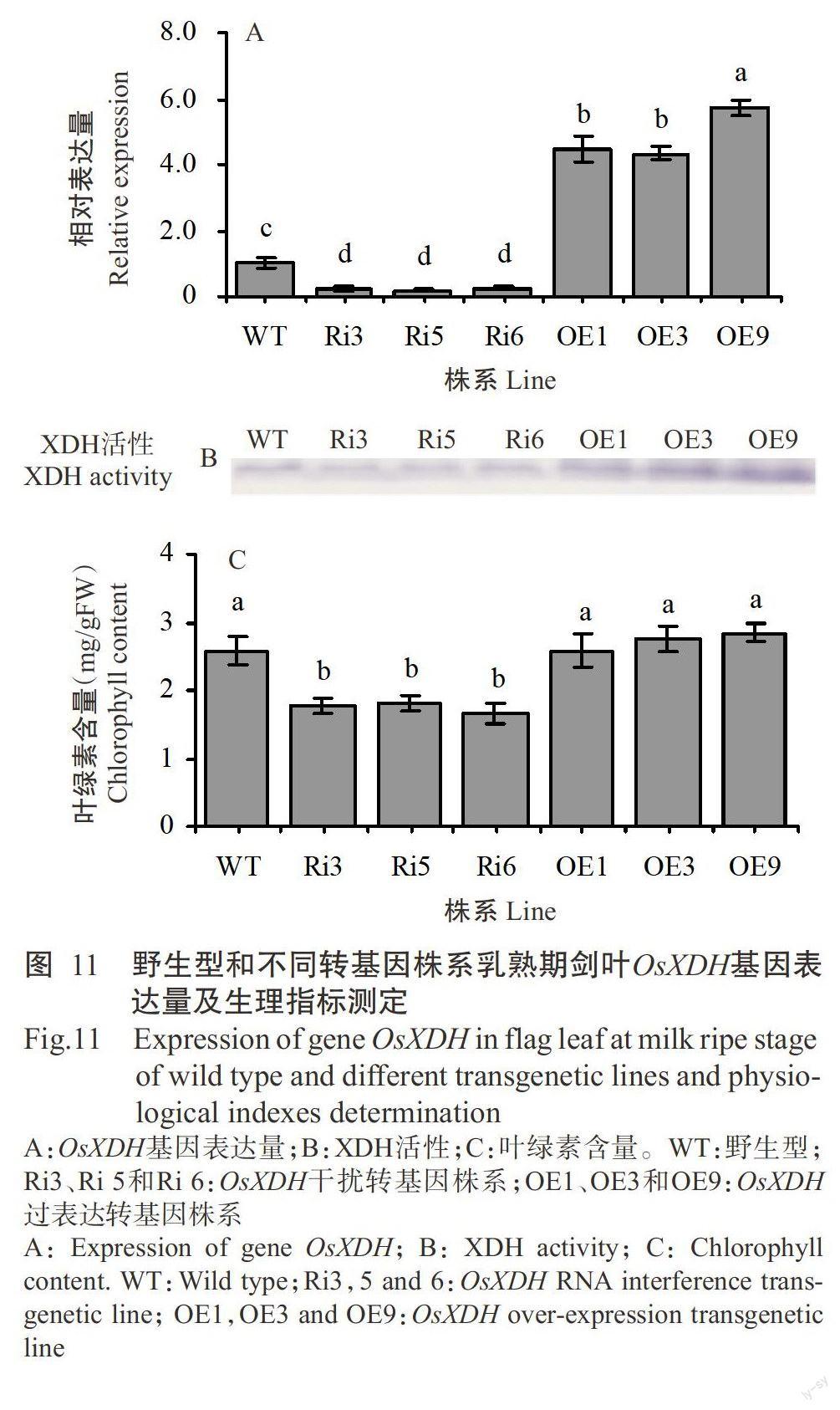
對野生型、OsXDH干擾轉基因株系及過表達轉基因株系乳熟期劍葉的OsXDH基因表達量、XDH活性和葉綠素含量進行比較分析,結果表明,OsXDH干擾轉基因株系的OsXDH基因表達量均顯著低于野生型和OsXDH過表達轉基因株系,OsXDH過表達轉基因株系的OsXDH基因表達量均顯著高于野生型(圖11-A);OsXDH干擾轉基因株系XDH活性低于野生型,而OsXDH過表達轉基因株系高于野生型(圖11-B);OsXDH干擾轉基因株系的葉綠素含量顯著低于野生型和OsXDH過表達轉基因株系,OsXDH過表達轉基因株系的葉綠素含量高于野生型,但未達顯著水平(圖11-C)。表明OsXDH基因表達量、XDH活性與葉綠素含量呈正相關,結合水稻不同生育時期OsXDH基因的表達特性,推測OsXDH基因參與調控水稻生育后期的生長發育。
3討論
本研究從粳稻品種日本晴中克隆獲得OsXDH基因ORF序列,長度為4110 bp,編碼1369個氨基酸。已有研究表明,擬南芥和蘭科植物的XDH基因均屬于核基因(Xiong et al.,2001;Hesberg et al.,2004;G6miak et al.,2010),但本研究中不同生物信息學軟件對OsXDH蛋白亞細胞定位結果不同,故未能明確其位置,需利用免疫熒光、GFP融合表達等技術進行蛋白亞細胞定位。此外,本研究通過同源比對及聚類分析,發現水稻OsXDH蛋白的氨基酸序列與單子葉作物高粱、小米和小麥的同源性較高,均在80.00%以上,雖然單子葉與雙子葉植物分屬不同分支,但其同源性仍在70.00%以上,證明不同植物中XDH蛋白的氨基酸序列具有高度的保守性,推測其在不同植物中發揮相似的生理功能。
本研究發現OsXDH基因在不同組織及不同生育期均有表達,但其在水稻灌漿期的表達量顯著高于苗期和分蘗盛期,表明OsXDH基因為特異性表達。同時本研究發現,在干旱、黑暗、高溫和鹽脅迫下,OsXDH基因表達量顯著上調,尤其在干旱和高溫脅迫下其增至脅迫處理前的2.5倍,推測水稻OsX-DH基因參與逆境脅迫響應。這與Hesberg等(2004)研究發現擬南芥中AtXDH1基因受不同逆境脅迫因子誘導均表達上調的結論一致。說明不同植物中的XDH基因具有相似表達調控功能。
本研究利用OsXDH干擾轉基因株系和過表達轉基因株系,對OsXDH基因的功能進行初步鑒定,結果顯示,OsXDH干擾轉基因株系中的XDH活性和葉綠素含量低于野生型,而OsXDH過表達轉基因株系的XDH活性和葉綠素含量均高于野生型,表明通過調控OsXDH基因的表達水平可調節水稻XDH活性,從而調控水稻的生長發育,與前人在擬南芥中的研究結果(Nakagawa et al.,2007;Watanabe et al.,2010;Zarepour et al.,2010;Watanabe et al.,2014)一致。由此推測OsXDH基因參與調控水稻生育后期的生長發育。在今后的研究中,可利用OsXDH~-擾轉基因株系和過表達轉基因株系進一步研究OsX-DH基因水稻在生長發育及逆境脅迫應答過程中的調控機制。
4結論
OsXDH基因受水稻生長發育和逆境脅迫因子誘導表達,推測其是調控水稻生長發育和響應逆境脅迫的關鍵基因。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
