冷鮮灘羊肉微生物菌群代謝物與其關聯性分析
2017-05-12 04:09:01胡倩倩張同剛尤麗琴羅瑞明李俊麗
食品工業科技 2017年8期
胡倩倩,張同剛,尤麗琴,羅瑞明,李俊麗
(寧夏大學農學院,寧夏銀川 750021)
冷鮮灘羊肉微生物菌群代謝物與其關聯性分析
胡倩倩,張同剛,尤麗琴,羅瑞明*,李俊麗
(寧夏大學農學院,寧夏銀川 750021)
本文以冷鮮灘羊肉為研究對象,采用選擇性培養基得出了寧夏鹽池縣冷鮮灘羊肉表面初始菌相構成為假單胞菌、腸桿菌、乳酸菌及熱死環絲菌。結合課題組成員使用GC-TOF-MS技術所得的微生物差異代謝物,用主成分分析微生物菌群與其關聯性。主成分分析的結果表明:與假單胞菌關聯程度較大的差異代謝物為D-甘油酸和苯丙氨酸,且代謝物D-甘油酸與苯丙氨酸之間、菌群與代謝物之間均呈現出正相關關系(相關系數矩陣值為正)。而1-磷酸葡萄糖、5′-肌苷酸和乳酸菌關聯性較大,且代謝物1-磷酸葡萄糖與5′-肌苷酸之間呈現出正相關關系,而代謝物與菌群呈現出負相關關系(相關系數矩陣值為負)。差異代謝物天冬氨酸、天冬酰胺、乙酰苯胺和核糖與腸桿菌及熱死環絲菌菌群相互聯系與影響的程度較大,且代謝物天冬酰胺、乙酰苯胺和核糖與菌群之間負相關,而代謝物天冬氨酸與菌群之間正相關;代謝物天冬氨酸與代謝物天冬酰胺、核糖負相關,與乙酰苯胺正相關。本文為揭示菌落變化的內在機制提供理論參考,為冷鮮肉保鮮分子生態學理論構建提供借鑒。
冷鮮灘羊肉,微生物菌群,差異代謝物,關聯性
健康動物由于自身具備完善的防御系統,一般情況下其血液和肌肉處于無菌狀態,而肉品變質主要源于外源性微生物污染[1]。在屠宰、加工和貯藏過程中,肉品表面首當其沖的被外環境中的好氧微生物污染[2],隨著微生物不斷的生長繁殖,微生物間開始相互競爭,而最終能抵抗或適應一切不利的生長條件,且對肉品腐敗發揮主要作用的微生物只有一小部分[3-5],深入探究這些優勢菌群的種類、生長狀態及代謝過程能使我們在解決肉類腐敗變質問題時做到對癥下藥[6-7]。由于肉品自身質構及生產加工工藝的不同,其初始菌相存在差異。羅欣等[8]在研究乳酸鈉在冷卻牛肉保鮮中的應用時,鑒定出實驗牛肉材料的初始菌相由假單胞菌屬(Pseudomonasspp.)、乳酸菌(Lactobacillus)、熱死環絲菌(Brchothrixthermosphacta)、酵母菌(Saccharomycetes)等構成;李苗云等[9]發現冷卻豬肉貯藏過程中的優勢菌為假單胞菌屬、熱死環絲菌、氣單孢菌屬(Aeromonas),然而只要初始菌相中存在葡萄球菌(Staphylococcus),它會逐步躍升為優勢菌。林惠珍[10]初步鑒定冰鮮鴨腿肉的主要腐敗菌除假單胞菌、乳酸菌、腸桿菌科(Enterobacter)之外,還有氣單孢菌屬、葡萄球菌、節桿菌(Arthrobacter)、熱死環絲菌。對于羊肉,劉瑩瑩等[11]采用選擇性培養基分析了寧夏吳忠地區冷卻羊肉初始菌相,指出該地區冷卻灘羊肉主要腐敗菌為假單胞菌、乳酸菌、腸桿菌科、葡糖球菌。但已有研究沒有揭示冷鮮肉微生物菌落差異代謝產物與其的關聯性。
本研究以冷鮮灘羊肉表面微生物為研究目標,將其接種于選擇性培養基上鑒定其類別并觀察其生長繁殖的狀況,并結合本課題組成員所得的差異代謝物,分析討論菌落代謝物與其關聯性,為揭示菌落變化的內在機制提供理論參考,為冷鮮肉保鮮分子生態學理論構建提供借鑒。
1 材料與方法
1.1 材料與儀器
灘羊后腿肉 購自寧夏大夏牧場清真食品有限公司(灘羊飼養期間不做任何生物性防疫工作,且定期進行清糞、消毒處理);瓊脂培養基:甘露醇鹽瓊脂(MSA)、假單胞菌選擇培養瓊脂(PSA)、紫紅膽鹽葡萄糖瓊脂(VRBGA)、MRS瓊脂、STA瓊脂平板計數瓊脂(PCA) 均購自上海麗臣商貿有限公司。
YXQ-SG46-280S手提式滅菌鍋 上海博訊醫療生物儀器有限公司;BPH-9052恒溫培養箱 上海一恒科學儀器有限公司;WT-B3003電子天平 杭州萬特衡器有限公司;JJ-2組織搗碎機 常州恒隆儀器有限公司;SW-CJ-1FD無菌工作臺 上海博迅科技有限公司;BCD-251冰箱 青島海爾股份有限公司。
1.2 實驗方法
1.2.1 樣品處理 新鮮采購的灘羊當天屠宰后立刻放入冷藏箱,運回實驗室后迅速放入-20 ℃冰箱1 h,使其中心溫度降至0 ℃,在無菌條件下,將灘羊后腿肉分割成重量為250 g相等的4塊,保鮮膜密封包裝,放在0 ℃冰箱分別貯藏0、4、8、12 d[12]后,在無菌條件下,處理得到后腿肉的表面刮擦物,充分進行破碎。
1.2.2 冷鮮灘羊肉微生物胞內差異代謝物的獲取 本課題組成員采用氣相色譜-飛行時間質譜聯用儀(GC-TOF-MS),且Agilent 7890氣相色譜-飛行時間質譜聯用儀配有Restek Rxi-5Sil MS毛細管柱(30 m×250 μm×0.25 μm),Restek,USA的工作條件下獲得了30種冷鮮灘羊肉微生物胞內差異代謝物。
1.2.3 冷鮮灘羊肉表面腐敗菌的培養 本課題組成員采用宏基因組測序技術對冷鮮灘羊肉微生物多樣性[13]的研究指出,肉品中酵母和霉菌菌落數量的對數值均小于1,而優勢菌集中在假單胞菌屬、腸桿菌科、熱死環絲菌、乳酸菌和葡萄球菌等細菌菌群上,因此本實驗以選擇性培養基為基礎,應用平板計數法對細菌總數、假單胞菌屬、腸桿菌科、熱死環絲菌屬、乳酸菌、葡萄球菌數量進行統計。
取被充分破碎后的冷鮮灘羊肉表面刮擦物25 g,加入225 mL、0.85%滅菌生理鹽水,振蕩均勻,取上清液1 mL進行10倍遞增梯度稀釋,取三個合適的稀釋梯度,平行梯度三次重復,傾注平板計數[14]。表面主要腐敗菌培養條件見表1。
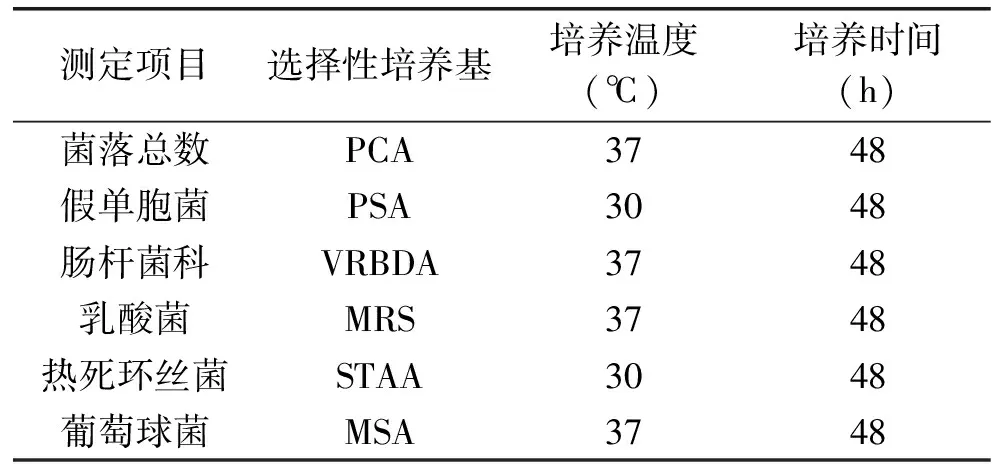
表1 主要腐敗菌培養條件Table 1 Culture conditions for major spoilage organisms
1.2.4 主成分分析 使用SPSS 13.0軟件選擇了12個組份進行了主成分分析,包括D-甘油酸(X1)、1-磷酸葡萄糖(X2)、天冬酰胺(X3)、核糖(X4)、5′-肌苷酸(X5)、乙酰苯胺(X6)、天冬氨酸(X7)和苯丙氨酸(X8)八種差異代謝物間接濃度和假單胞菌(X9)、腸桿菌科(X10)、乳酸菌(X11)及熱死環絲菌(X12)四種主要腐敗菌數量。
1.2.5 數據處理 使用SPSS 13.0統計軟件進行數據分析,測定結果以均數和標準差表示。實驗數據分析載荷圖和相關系數矩陣得出結論。
2 結果與分析
2.1 冷鮮灘羊肉表面的初始菌相
以選擇性培養基為基礎,應用平板計數法對細菌總數、假單胞菌屬、腸桿菌科、熱死環絲菌、乳酸菌、葡萄球菌數量進行統計,結果如表2所示。
從表2中可得出,托盤密封包裝的冷鮮灘羊肉表面的初始菌相主要包含假單胞菌屬、腸桿菌科、乳酸菌、熱死環絲菌四類腐敗菌。其中初始優勢菌群為假單胞菌,其次為腸桿菌科。

表2 冷鮮灘羊肉初始菌相分布Table 2 The initial distribution of bacterial phase in chilled Tan sheep meat
這與劉瑩瑩[15]等報道的寧夏吳忠地區冷卻灘羊肉初始菌相構成存在差異的菌相為熱死環絲菌,可能是由于屠宰分割、胴體冷卻、殺毒滅菌工藝不同或地域區別而引起的。此外,袁東曉等研究將冷鮮肉初始菌數控制在102~103cfu/g范圍內,有助于其貨架期的延長[16]。本實驗樣本初始菌落總數對數值為3.89 lg cfu/g,超出了有助于延長貨架期的控制范圍,因而推定該冷鮮灘羊肉保質期可能不會很長。
2.2 灘羊肉表面主要腐敗菌菌相變化態勢
托盤密封包裝的冷鮮灘羊肉貯藏在0 ℃時,表面主要腐敗菌菌相變化態勢見圖1。
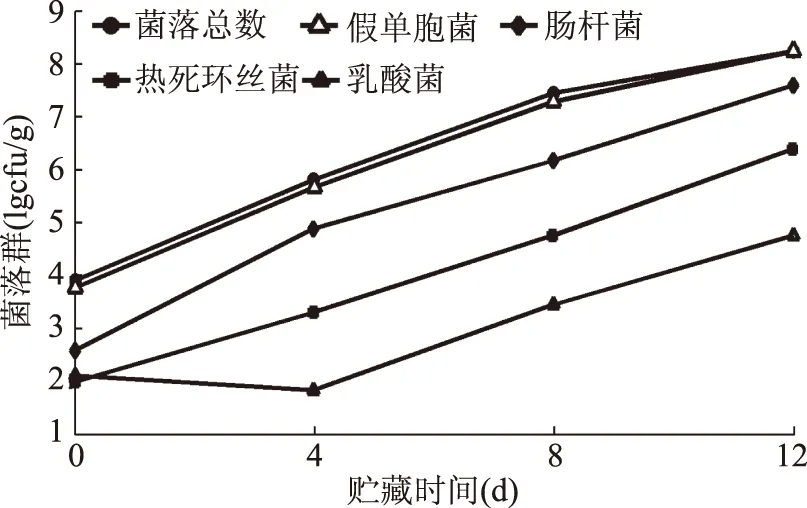
圖1 冷鮮灘羊肉主要腐敗菌菌相變化趨勢圖Fig.1 Bacteria phase change of the main spoilage organisms of chilled Tan sheep meat
在0 ℃冷鮮貯藏過程中,托盤密封包裝灘羊肉主要腐敗菌總體上均呈現出菌落數不斷增長的態勢,但是乳酸菌在0~4 d菌落數有所下降,扈麟等人研究此現象原因可能有兩個[17]:一是乳酸菌為厭氧菌,外部環境中的氧氣組分會阻礙其生長;二是微生物間相互作用,使乳酸菌的生長受到抑制。就平均增長速率而言,假單胞菌屬和腸桿菌科的平均增長率整體上大于乳酸菌和熱死環絲菌,再加上假單胞菌和腸桿菌科的初始污染菌量也遠大于乳酸菌及熱死環絲菌,因此,冷鮮灘羊肉表面優勢腐敗菌為假單胞菌和腸桿菌科。該結果與本課題組成員張赫宇等人采用宏基因組學技術測定的冷鮮灘羊肉表面優勢腐敗菌為假單胞菌和腸桿菌科結果[13]相一致。
通常情況下,106cfu/g的細菌總數被認為是冷鮮肉品質劣變的臨界點,一旦菌落總數突破該臨界點增長到107cfu/g時,冷鮮肉在感官品質方面(氣味、顏色及觸感)有明顯的變質跡象,而當菌落總數飆升到108cfu/g時,其表面會形成一層粘液,失去食用價值。一些國家冷鮮肉的細菌總數合格標準為106cfu/g[18],若以該標準作為評判依據,實驗條件下冷鮮灘羊肉的貨架期可能為在7~8 d;若以出現明顯劣變現象為標準,實驗條件下冷鮮灘羊肉的貨架期可能在9~10 d[19]。考慮到以第一種依據得到的貨架期更為嚴格,基于安全第一原則,建議以國家細菌總數合格標準為評判依據。
2.3 冷鮮灘羊肉菌群代謝物與其關聯性分析
結合本課題成員采用GC-MS測得的30種冷鮮灘羊肉微生物胞內差異代謝物中,D-甘油酸(D-Glyceric acid)、天冬酰胺(Asparagine)、1-磷酸葡萄糖(Glucose-1-phosphate)、5′-肌苷酸(Inosine-5′-monophosphate)、核糖(Ribose)、乙酰苯胺(Acetanilide)是在四個貯藏時間段內呈現出兩次差異的代謝物,而差異代謝物中天冬氨酸(Aspartic acid)是必需氨基酸,異亮氨酸、蘇氨酸和賴氨酸的前體物質及差異代謝物中必需氨基酸苯丙氨酸(Phenylalanine)的Similarity值大于900且OPLS-DA模型變異權重系數VIP值較大,故以D-甘油酸、天冬酰胺、1-磷酸葡萄糖、5′-肌苷酸、乙酰苯胺、核糖、天冬氨酸和苯丙氨酸八種差異物作為分析代謝物與不同時間段菌落差異代謝物,其間接表示相對濃度的峰面積平均值見表3。
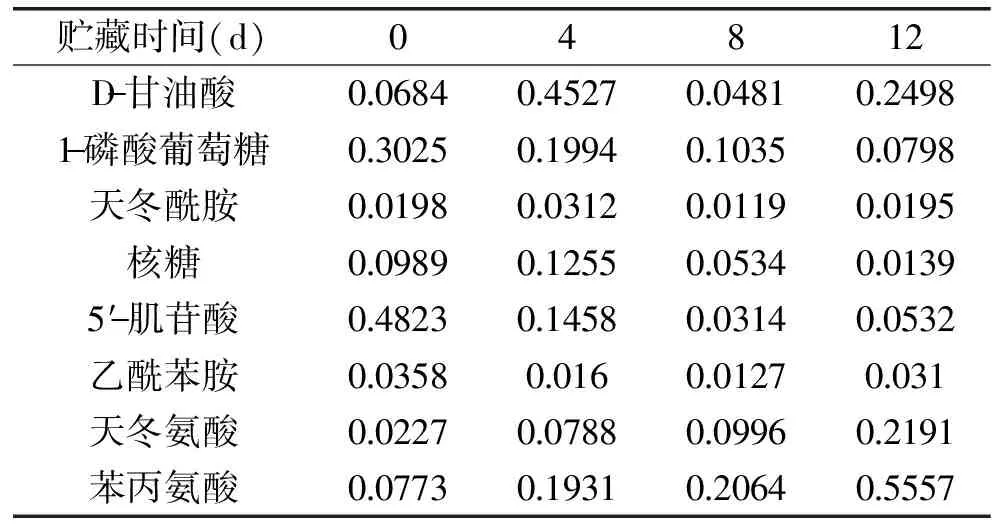
表3 不同貯藏時間差異代謝物的峰面積平均值Table 3 The average peak area of differential metabolites during storage
使用SPSS 13.0統計軟件將隨著貯藏時間變化而變化的差異代謝物峰面積平均值和表面主要腐敗菌數量進行主成分分析,主成分分析的結果見表4~表6。
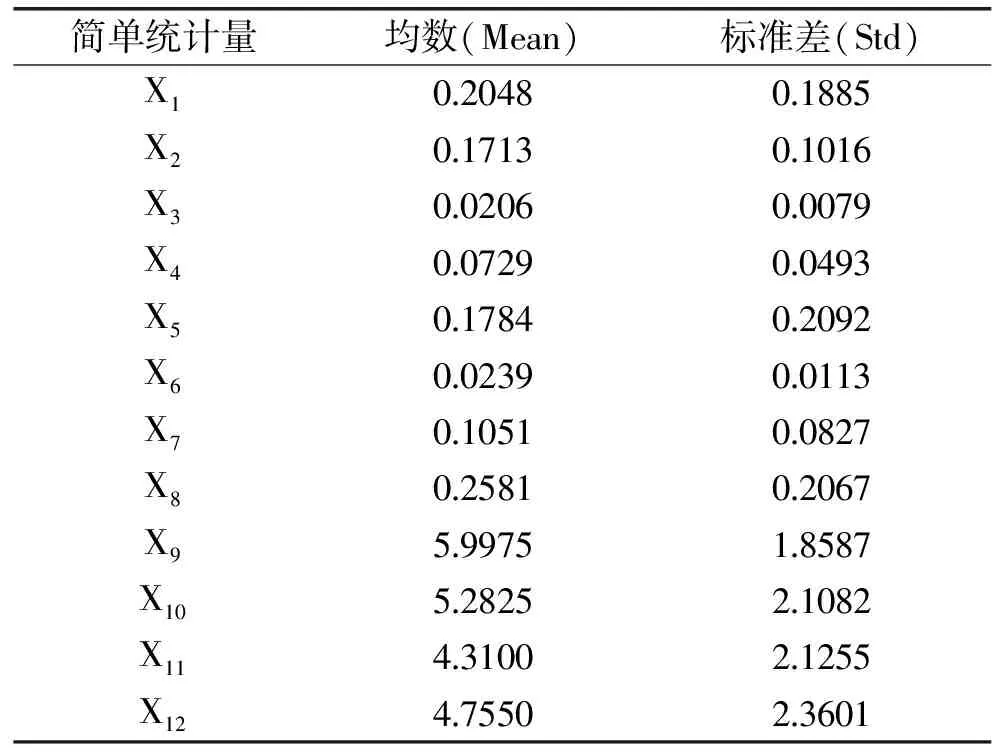
表4 代謝物與四種主要腐敗菌主成分分析數據集信息Table 4 PCA data information of metabolites and spoilage organisms

表5 代謝物與四種主要腐敗菌相關系數矩陣特征值Table 5 Correlation matrix eigenvalues of correlation analysis of metabolites and spoilage organisms
從表5中可得,第一主成分、第二主成分和第三主成分的特征值均大于1,故需再以貢獻率為著手點,進一步對其進行考察,第一主成分的貢獻率0.6892,說明第一主成分能提供原始變量提供的綜合信息的一半以上,但不到80%;第二主成分的貢獻率為0.1889,與第一主成分貢獻率合并的累計貢獻率達到87%,說明前兩個主成分能提供87%以上原始變量的綜合信息,所以在冷鮮灘羊肉菌群代謝物與其關聯性分析中保留前兩個主成分。

表6 代謝物與四種主要腐敗菌相關分析特征向量Table 6 Eigenvector of correlation analysis of metabolites and spoilage organisms

表7 代謝物與菌群關聯性分析相關系數矩陣Table 7 Matrix of the correlation coeffient of metabolites and spoilage organisms
3 結論
冷鮮灘羊肉微生物群落變化是從復雜到簡單的過程。本文確定了寧夏鹽池地區冷鮮灘羊肉的初始菌相為假單胞菌、腸桿菌、熱死環絲菌及乳酸菌;并指出其中的優勢菌群為假單胞菌,其次為腸桿菌科。為了探明差異代謝物與菌群的關聯性,主成分分析及聚類分析相關數據說明與假單胞菌關聯程度較大的差異代謝物為D-甘油酸和苯丙氨酸,且代謝物D-甘油酸與苯丙氨酸之間、菌群與代謝物之間均呈現出正相關關系。而1-磷酸葡萄糖、5′-肌苷酸和乳酸菌關聯性較大,且代謝物1-磷酸葡萄糖與5′-肌苷酸之間呈現出正相關關系,而代謝物與菌群呈現出負相關關系。差異代謝物天冬氨酸、天冬酰胺、乙酰苯胺和核糖與腸桿菌及熱死環絲菌菌群相互聯系與影響的程度較大,且代謝物天冬酰胺、乙酰苯胺和核糖與菌群之間負相關,而代謝物天冬氨酸與菌群之間正相關;代謝物天冬氨酸與代謝物天冬酰胺、核糖負相關,與乙酰苯胺正相關。宏基因組測序技術可被應用于分析探究微生物群體豐度和多樣性,該方法既可將繁瑣的微生物培養分離過程避開,而且可靈敏的將外界環境對微生物群落結構的影響探測出來,為微生物的研究開辟了新的思路與方向。
[1]臧晉,劉鳳霞,李永祥,等.高產醋酸菌的分離選育[J].中國釀造,1999(5):20-22.
[2]孟憲剛,周鴿鴿,張麗珂. 發酵蔬菜漿水中優勢好氧微生物的分離及鑒定[J]. 食品工業科技,2010(3):222-224.
[3]Carlez A,J-P R,Richard N,et al. High Pressure Inactivation of Citrobacter freundii,Pseudomonas fluorescens and Listeria innocua in Inoculated Minced Beef Muscle[J]. LWT-Food Science and Technology,1993,26(4):357-363.
[4]Garriga M,Grèbol N,Aymerich M T,et al. Microbial inactivation after high-pressure processing at 600 MPa in commercial meat products over its shelf life[J]. Innovative Food Science & Emerging Technologies,2004,5(4):451-457.
[5]Chen H. Temperature-assisted pressure inactivation of Listeria monocytogenes in turkey breast meat[J]. International Journal of Food Microbiology,2007,117(1):55-60.
[6]李旻. 人體腸道菌群結構與宿主代謝的相關性研究[D]. 上海:上海交通大學,2009:15-21.
[7]張艷. 醬香型白酒發酵中乳酸菌群結構及功能研究[D]. 無錫:江南大學,2015:17-26.
[8]羅欣,朱燕. 乳酸鈉在牛肉冷卻肉保鮮中的應用研究[J]. 食品與發酵工業,2000,26(3):1-5.
[9]李苗云,周光宏,徐幸蓮. 應用PCR-DGGE研究冷卻豬肉貯藏過程中的優勢菌[J]. 西北農林科技大學學報:自然科學版,2008,36(9):185-189.
[10]林惠珍. 冰鮮鴨優勢腐敗菌的分析及其控制技術的研究[D]. 福州:福建農業大學,2011:12-25.
[11]劉瑩瑩,羅瑞明,盧君逸,等. 冷卻羊肉貯藏中品質變化及假單胞菌生長預測模型的建立[J]. 肉類研究,2013,27(5):5-9.
[12]Wang C,Geng X. Refolding and purification of recombinant human granulocyte colony-stimulating factor using hydrophobic interaction chromatography at a large scale[J]. Process Biochemistry,2012,47(12):2262-2266.
[13]張赫宇,楊波,羅瑞明,等. 高通量測序分析冷鮮灘羊肉儲藏過程中的細菌群落多樣性[J]. 食品工業科技,2016(13):177-182.
[14]杜宗緒. 3種平板計數瓊脂培養基的質量比較[J]. 山西農業科學,2014,42(8):835-837.
[15]劉瑩瑩,羅瑞明,盧君逸,等. 冷卻羊肉貯藏中品質變化及假單胞菌生長預測模型的建立[J]. 肉類研究,2013,27(5):5-9.
[16]袁東曉. 冷卻肉防腐劑的研究[D].上海:華東師范大學,2006:4-8.
[17]扈麟. 冷卻豬肉腐敗的細菌群體感應研究[D].長沙:湖南農業大學,2010:20-24.
[18]中華人民共和國衛生部. GB/T 9959.2-2008分割鮮、凍豬豬瘦肉[S]. 北京:中國標準出版社,2008.
[19]彭佳程. 冷鮮肉品質安全控制技術的研究[D].武漢:武漢輕工大學,2014:37-40.
特征向量是衡量主成分和原變量關聯性大小的,關聯性越大,則特征向量的絕對值越大。表6的結果表明:第一主成分與X12(熱死環絲菌)、X9(假單胞菌)、X11(乳酸菌)、X10(腸桿菌科)、X2(1-磷酸葡萄糖)、X7(天冬氨酸)、X8(苯丙氨酸)和X5(5′-肌苷酸)關系比較密切,第二主成分與X1(D-甘油酸)、X3(天冬酰胺)、X4(核糖)及X6(乙酰苯胺)的關系比較密切。主成分載荷圖見圖2。

圖2 主成分得分圖Fig.2 Plot of principal component analysis(PCA)
由主成分一和主成分二的載荷圖2和表7相關系數矩陣可知,與假單胞菌關聯程度較大的差異代謝物為D-甘油酸和苯丙氨酸,且代謝物D-甘油酸與苯丙氨酸之間、菌群與代謝物D-甘油酸與苯丙氨酸之間均呈現出正相關關系。而1-磷酸葡萄糖、5′-肌苷酸和乳酸菌關聯性較大,且代謝物1-磷酸葡萄糖與5′-肌苷酸之間呈現出正相關關系,而代謝物1-磷酸葡萄糖與5′-肌苷酸與菌群呈現出負相關關系。差異代謝物天冬氨酸、天冬酰胺、乙酰苯胺和核糖與腸桿菌及熱死環絲菌菌群相互聯系與影響的程度較大,且代謝物天冬酰胺、乙酰苯胺和核糖與菌群之間負相關,而代謝物天冬氨酸與菌群之間正相關;代謝物天冬氨酸與代謝物天冬酰胺、核糖負相關,與乙酰苯胺正相關。
Correlation analysis of microbial community and their metabolites in cold fresh Tan lamb
HU Qian-qian,ZHANG Tong-gang,YOU Li-qin,LUO Rui-ming*,LI Jun-li
(College of Agricultural,Ningxia University,Yinchuan 750021,China)
In this paper,based on the cold fresh Tan lamb as the research object,using the selective culture method,the initial phases on the chilled meat surface of Tan sheep in Yanchi,Ningxia were determined asPseudomonas,Enterobacter,BrchothrixthermosphactaandLactobacillus,and the dominant bacteria wasPseudomonasfollowed byEnterobacter. The differential metabolites were obtained by gas chromatography time of flight mass spectrometry(GC-TOF-MS)in combination with the members of the group. Principal component analysis was used to discuss the relationship between the microbial community and their metabolites. The results of principal component analysis showed that it had greater relevance betweenPseudomonasand D-glyceric acid and phenylalanine,and the relationships of D-glyceric acid and phenylalanine as well as flora and metabolite had shown a positive correlation tendency. The correlation coefficient matrix was positive. The relevance between glucose-1-phosphate,Inosine-5′-monophosphate andLactobacilluswas significant. Among the metabolites of glucose 1-phosphate and 5′-inosinic acid showed a positive correlation,and the metabolites and bacteria showed a negative correlation. The correlation coefficient matrix was negative. Meanwhile,aspartic acid,asparagine,ribose and acetanilide had great interrelation withEnterobacterandBrchothrixthermosphacta. The metabolites of asparagine,acetanilide had the negative correlation with and ribose and bacterium group,while the aspartic acid and bacterium group had the positive correlation.The metabolite of aspartic acid had negative correlation with asparagine and ribose,positive correlation with acetanilide. This paper provides a theoretical reference for revealing the inner mechanism of colony change,reference for the construction of cold fresh meat molecular ecology theory.
the cold fresh Tan lamb;microbial community;differential metabolites;relatedness
2016-09-22
胡倩倩(1991-),女,碩士研究生,研究方向:畜產品加工,E-mail:1570687113@qq.com。
*通訊作者:羅瑞明(1964-),男,博士,教授,研究方向:畜產品貯藏與加工,E-mail:ruiming@126.com。
國家自然科學基金地區基金項目(31460431)。
TS251.5
A
1002-0306(2017)08-0322-05
10.13386/j.issn1002-0306.2017.08.054
