氯喹對(duì)戊四氮慢性致癇大鼠的抗癲癇作用及對(duì)腦組織內(nèi)mGluR5、mGluR1表達(dá)的影響
2017-05-04 01:58:55荊麗麗高平解水杉曹慶華濱州醫(yī)學(xué)院煙臺(tái)附屬醫(yī)院山東煙臺(tái)264100
山東醫(yī)藥 2017年13期
關(guān)鍵詞:癲癇
荊麗麗, 高平,解水杉,曹慶華(濱州醫(yī)學(xué)院煙臺(tái)附屬醫(yī)院,山東煙臺(tái) 264100)
氯喹對(duì)戊四氮慢性致癇大鼠的抗癲癇作用及對(duì)腦組織內(nèi)mGluR5、mGluR1表達(dá)的影響
荊麗麗, 高平,解水杉,曹慶華
(濱州醫(yī)學(xué)院煙臺(tái)附屬醫(yī)院,山東煙臺(tái) 264100)
目的 探討氯喹對(duì)戊四氮慢性致癇大鼠的抗癲癇作用及對(duì)腦組織內(nèi)代謝型谷氨酸受體5(mGluR5)及代謝型谷氨酸受體1(mGluR1)表達(dá)的影響。方法 選擇健康雄性SD大鼠36只,隨機(jī)分為對(duì)照組(12只)、戊四氮致癇組(12只)、氯喹干預(yù)組(12只)。對(duì)照組:腹腔注射生理鹽水,40 mg/kg,1次/24 h;戊四氮致癇組與氯喹干預(yù)組:腹腔注射0.5%戊四氮溶液,40 mg/kg,1次/24 h。連續(xù)給藥,直至動(dòng)物模型制備成功。模型制備成功后,氯喹干預(yù)組腹腔注射0.5%氯喹,40 mg/kg,1次/24 h,持續(xù)2周。干預(yù)結(jié)束后,觀察并記錄各組大鼠行為學(xué)表現(xiàn),采用Western blotting法檢測(cè)大鼠腦組織內(nèi)mGluR5蛋白的相對(duì)表達(dá)量,免疫組化法檢測(cè)大鼠腦組織內(nèi)mGluR1蛋白的相對(duì)表達(dá)量。結(jié)果 對(duì)照組未出現(xiàn)癲癇發(fā)作,戊四氮致癇組可見點(diǎn)頭、面部抽動(dòng)、前肢陣攣及全身陣攣、跌倒等表現(xiàn),氯喹干預(yù)組癲癇發(fā)作程度較戊四氮致癇組減輕。與對(duì)照組、氯喹干預(yù)組相比,戊四氮致癇組mGluR5及mGluR1蛋白相對(duì)表達(dá)量增多(P均<0.05);氯喹干預(yù)組與對(duì)照組相比,P均>0.05。結(jié)論 氯喹能夠減輕戊四氮慢性致癇大鼠的癲癇發(fā)作,抑制腦組織內(nèi)mGluR5及mGluR1的表達(dá)。
癲癇;氯喹;戊四氮;代謝型谷氨酸受體5;代謝型谷氨酸受體1;大鼠
癲癇是慢性反復(fù)發(fā)作性短暫腦功能失調(diào)綜合征,是以腦神經(jīng)元異常放電引起反復(fù)癇性發(fā)作為特征的中樞神經(jīng)系統(tǒng)疾病,發(fā)病原因極其復(fù)雜。興奮性神經(jīng)遞質(zhì)與抑制性神經(jīng)遞質(zhì)失衡在癲癇中的作用尤為重要。興奮性神經(jīng)遞質(zhì)包括谷氨酸(Glu)和天門冬氨酸,Glu廣泛分布于中樞神經(jīng)系統(tǒng),通過與Glu受體結(jié)合發(fā)揮作用。Glu受體包括離子型谷氨酸受體、代謝型谷氨酸受體(mGluR)。mGluR可借由間接代謝活動(dòng)進(jìn)行活化,該受體又分為8種不同的亞型(mGluR1~8),分3組,其中第Ⅰ組包括mGluR1及mGluR5,與G蛋白相偶聯(lián)發(fā)揮作用。氯喹作為傳統(tǒng)的抗瘧疾藥物,其具有廣泛的藥理作用,可抑制DNA復(fù)制、蛋白質(zhì)合成等。前期研究[1~3]表明,氯喹可以抑制mGluR及神經(jīng)性一氧化碳合酶等的表達(dá),從而控制戊四氮導(dǎo)致的大鼠癲癇發(fā)作。氯喹是否可以通過抑制mGluR5及mGluR1的表達(dá),從而達(dá)到抑制癲癇發(fā)作的目的,尚未見研究報(bào)道。2015年1月~2016年12月,本研究探討了氯喹對(duì)戊四氮慢性致癇大鼠的抗癲癇作用及對(duì)腦組織內(nèi)mGluR5及mGluR1表達(dá)的影響,以期對(duì)癲癇的治療提供有價(jià)值的資料。
1 材料與方法
1.1 材料 選擇SPF級(jí)健康雄性SD大鼠36只,體質(zhì)量120~150 g,環(huán)境溫度20~25 ℃,濕度40%~70%,單籠飼養(yǎng),自由飲水、進(jìn)食,每天稱重,SD大鼠購(gòu)自山東煙臺(tái)綠葉制藥有限公司,動(dòng)物合格證號(hào):SCXK(魯)20030008;戊四氮、氯喹(美國(guó)Sigma公司);兔抗mGluR5抗體、兔抗mGluR1抗體、兔抗β-actin(美國(guó)Santa Cruz公司);HPLAS-1000高清晰彩色病理圖文分析系統(tǒng)(日本Olympus)。
1.2 戊四氮癲癇大鼠模型制備及分組 將大鼠隨機(jī)分為對(duì)照組(12只)、戊四氮致癇組(12只)、氯喹干預(yù)組(12只)。對(duì)照組:腹腔注射生理鹽水,40 mg/kg,1次/24 h;戊四氮致癇組與氯喹干預(yù)組:腹腔注射0.5%戊四氮溶液,40 mg/kg,1次/24 h。連續(xù)給藥,直至動(dòng)物模型制備成功。觀察藥物注射后實(shí)驗(yàn)動(dòng)物反應(yīng),每次觀察至給藥后2 h,連續(xù)給藥7 d左右,大鼠出現(xiàn)癇樣發(fā)作,持續(xù)給藥,觀察并記錄行為學(xué)表現(xiàn),采用Racine評(píng)分標(biāo)準(zhǔn),至少連續(xù)出現(xiàn)5次Ⅱ級(jí)以上癇樣發(fā)作的大鼠被認(rèn)為造模成功。模型制備成功后,氯喹干預(yù)組腹腔注射0.5%氯喹,40 mg/kg,1次/24 h,持續(xù)2周。Racine評(píng)分標(biāo)準(zhǔn):0級(jí):無發(fā)作;Ⅰ級(jí):面部肌肉、口部或耳部出現(xiàn)節(jié)律性抽動(dòng)、陣攣;Ⅱ級(jí):面部肌肉抽動(dòng)陣攣嚴(yán)重,伴點(diǎn)頭;Ⅲ級(jí):前肢陣攣,不伴直立;Ⅳ級(jí):前肢陣攣并伴直立;Ⅴ級(jí):全身強(qiáng)直陣攣發(fā)作、跌倒。
1.3 腦組織內(nèi)mGluR5蛋白相對(duì)表達(dá)量的檢測(cè) 采用Western blotting法。檢測(cè)大鼠按實(shí)驗(yàn)分組,冰上快速開顱取腦,分離腦組織,加入裂解液裂解約30 min,移液,4 ℃離心機(jī)離心(12 000 r/min,5 min),留上清液,分裝。選用BCA法測(cè)定蛋白濃度,確定上樣體積(含20 μg蛋白)。采用8% SDS-PAGE電泳分離,根據(jù)預(yù)染Marker確定mGluR-5及β-actin的位置,并剪下相應(yīng)位置蛋白凝膠條帶,轉(zhuǎn)至PVDF膜上,免疫雜交反應(yīng)。于37 ℃封閉液中封閉2 h,加入山羊抗mGluR5抗體(1∶100)、兔抗β-actin(1∶100),4 ℃過夜,加入相應(yīng)二抗37 ℃孵育1 h,曝光。PVDF膜曬干,條帶掃描,分析灰度值,以mGluR5與β-actin的灰度值之比作為目的蛋白的表達(dá)量。
1.4 腦組織內(nèi)mGluR1蛋白相對(duì)表達(dá)量的檢測(cè) 采用免疫組化法。按照試驗(yàn)分組,應(yīng)用4%水合氯醛麻醉,冰上快速開顱取腦,分離提取腦組織,4%多聚甲醛固定,脫水、透明、浸蠟、包埋,切片,厚度3 μm,烤片,浸入二甲苯、梯度乙醇脫蠟至水。3% H2O2作用20 min以消除內(nèi)源性過氧化物酶的影響,檸檬酸高壓修復(fù),羊血清封閉,加入兔抗mGluR5抗體(1∶200)4 ℃過夜,加入即用型二抗檢測(cè)試劑盒,37 ℃水浴鍋孵育,DAB顯色。復(fù)染、鹽酸分化、乙醇脫水、透明、封片。各步驟間用PBS洗滌5 min/次,共3次。細(xì)胞質(zhì)呈棕褐色為陽(yáng)性結(jié)果。采用HPLAS-1000高清晰彩色病理圖文分析系統(tǒng)進(jìn)行圖像分析。在400倍視野下選擇圖像,分析平均光密度值(AOD)。
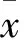
2 結(jié)果
2.1 各組行為學(xué)表現(xiàn) 對(duì)照組未出現(xiàn)癲癇發(fā)作,戊四氮致癇組可見點(diǎn)頭、面部抽動(dòng)、前肢陣攣及全身陣攣、跌倒等表現(xiàn),氯喹干預(yù)組癲癇發(fā)作程度較戊四氮致癇組減輕。
2.2 各組腦組織內(nèi)mGluR5蛋白相對(duì)表達(dá)量的比較 對(duì)照組、戊四氮致癇組、氯喹干預(yù)組mGluR5相對(duì)表達(dá)量分別為1.367±0.076、1.895±0.043、1.564±0.056,三組比較,P<0.05;與戊四氮致癇組比較,對(duì)照組及氯喹干預(yù)組mGluR5水平低(P均<0.05),對(duì)照組與氯喹干預(yù)組比較,P﹥0.05。見圖1。

注:N:對(duì)照組;P:戊四氮致癇組;C:氯喹干預(yù)組。
圖1 各組腦組織內(nèi)mGluR5蛋白的相對(duì)表達(dá)量(Western blotting法)
2.3 各組腦組織內(nèi)mGluR1蛋白的相對(duì)表達(dá)量比較 mGluR1以細(xì)胞質(zhì)染成棕褐色為陽(yáng)性結(jié)果。圖像分析結(jié)果顯示mGluR1陽(yáng)性細(xì)胞主要分布在海馬區(qū)。對(duì)照組、戊四氮致癇組、氯喹干預(yù)組mGluR1相對(duì)表達(dá)量分別為0.082 7±0.012 1、0.110 6±0.017 2、0.091 7±0.014 7。三組比較,P=0.000;戊四氮組高于對(duì)照組及氯喹干預(yù)組(P均<0.05)。對(duì)照組與氯喹干預(yù)組比較,P>0.05。
3 討論
Glu受體可借由間接代謝過程進(jìn)行活化,該受體可與Glu結(jié)合,具有興奮性神經(jīng)傳遞作用。Glu是中樞神經(jīng)系統(tǒng)重要的興奮性神經(jīng)遞質(zhì),癲癇狀態(tài)下Glu蓄積,由于再攝取障礙,激活Glu受體,神經(jīng)元異常放電,導(dǎo)致癲癇發(fā)作[4~6]。Ⅰ組mGluR主要通過與興奮性G蛋白偶聯(lián),激活磷脂酶,經(jīng)過一系列生化反應(yīng),刺激細(xì)胞內(nèi)儲(chǔ)備的Ca2+釋放,I組mGluR激活后,影響離子通道的離子轉(zhuǎn)運(yùn)而發(fā)揮作用[7]。在癲癇動(dòng)物模型及癲癇患者海馬區(qū)會(huì)出現(xiàn)Ⅰ組mGluR的表達(dá)增高[8],提示其與癲癇的發(fā)生有密切關(guān)系。mGluR5及mGluR1是Ⅰ組mGluR的亞型,廣泛分布于中樞神經(jīng)系統(tǒng),mGluR5及mGluR1的激活可提高神經(jīng)元興奮性,增強(qiáng)離子型Glu受體的功能。在中樞神經(jīng)系統(tǒng)的多種神經(jīng)紊亂病變中均存在mGluR5的功能障礙,如焦慮癥、抑郁癥、癲癇、神經(jīng)性疼痛、脆性X染色體綜合征、帕金森病及藥物成癮所引起的中樞神經(jīng)系統(tǒng)紊亂[9~11],在抗藥的顳葉癲癇患者海馬神經(jīng)元中mGluR5的表達(dá)明顯增高,分析認(rèn)為mGluR5表達(dá)的增高可能是癲癇抽搐的結(jié)果,并且mGluR5的高表達(dá)可能有利于維持其海馬的興奮性。Deutsch等[12]研究發(fā)現(xiàn),選擇mGluR5的拮抗劑用于一些神經(jīng)功能紊亂的病變中,結(jié)果證實(shí),mGluR5的拮抗劑可緩解患者病情。人類顳葉癲癇海馬mGluR1表達(dá)增高,能夠增加海馬癲癇發(fā)作易感性,遺傳性癲癇與mGluR1基因異常表達(dá)有關(guān),抗藥性癲癇患者海馬神經(jīng)元mGluR1表達(dá)明顯增加[13~15]。綜上認(rèn)為,Ⅰ組mGluR可作為癲癇的治療靶點(diǎn)。
氯喹最初用于瘧疾的治療,隨著醫(yī)學(xué)的發(fā)展,對(duì)氯喹的認(rèn)識(shí)不斷提升,目前,氯喹可用于多種疾病的治療,主要用于艾滋病病毒復(fù)制的防治。其通過干擾DNA復(fù)制、抑制蛋白質(zhì)合成發(fā)揮作用,還可抑制腫瘤壞死因子及白細(xì)胞介素發(fā)揮免疫調(diào)節(jié)作用,從而抑制一些病毒介導(dǎo)的炎癥反應(yīng)[16~18],且價(jià)格低廉,相對(duì)耐藥性好。本研究結(jié)果顯示,戊四氮可誘導(dǎo)試驗(yàn)動(dòng)物癲癇發(fā)作,使腦組織內(nèi)mGluR5及mGluR1興奮性增高,表達(dá)增強(qiáng),應(yīng)用氯喹干預(yù)后,mGluR5及mGluR1表達(dá)降低,大鼠癲癇發(fā)作程度減弱。提示氯喹可能通過抑制癲癇大鼠腦內(nèi)mGluR5及mGluR1的表達(dá)發(fā)揮抗癲癇作用。氯喹可作為一種神經(jīng)保護(hù)劑,但是其具體作用機(jī)制還有待于進(jìn)一步研究。
[1] 吳淑華,荊麗麗,朱曉晶,等.不同劑量氯喹對(duì)戊四氮慢性致癇大鼠腦內(nèi)nNOS表達(dá)的影響[J].神經(jīng)解剖學(xué)雜志,2012,28(4):392-396.
[2] 荊麗麗,吳淑華,楊東霞,等.氯喹對(duì)戊四氮慢性致癇大鼠還買GluR2及nNOS表達(dá)的影響[J].神經(jīng)解剖學(xué)雜志,2011,27(4):428-432.
[3] 荊麗麗,譚宏偉,于靜,等.不同劑量氯喹對(duì)戊四氮慢性致癇大鼠腦內(nèi)谷氨酸受體2表達(dá)的影響[J].中國(guó)醫(yī)藥導(dǎo)報(bào),2014,11(15):10-12.
[4] Campisi A, Gulino M, Acquaviva R, et al. Reactive oxygen species levels and DNA fragmentation on astrocytes in primary culture after acute exposure to low intensity microwave electromagnetic field [J]. Neurosci Lett, 2010,473(1):52-55.
[5] Juge N, Gray JA, Omote H, et al. Metabolic control of vesicular glutamate transport and release [J]. Neuron, 2010,68(1):99-112.
[6] Ramos-Mandujano G, Vazquez-Juarez E, Hernandez-Benítez R, et al. Thrombin potently enhances swelling-sensitive glutamate efflux from cultured astrocytes [J]. Glia, 2007,55(9):917-925.
[7] Neyman S, Manahan-Vaughan D. Metabotropic glutamate receptor 1 (mGluR1) and 5 (mGluR5) regulate late phases of LTP and LTD in hippocampal CA1 region in vitro[J]. Neurosci, 2008,27(6):1345-1352.
[8] Blumcke I, Becker AJ, Klein C, et al. Temporal lobe epilepsy associated up-regulation of metabotropic glutamate receptors: correlated changes in mGluR1 mRNA and protein expression in experimental animals and human patients [J]. Neuropathol Exp Neurol, 2000,59(1):1-10.
[9] Olive MF. Cognitive effects of Group Ⅰ metabotropic glutamate receptor ligands in the context of drug addiction [J]. Eur J Pharmacol, 2010,639(1-3):47-58.
[10] Borowicz KK, Luszczki JJ, Czuczwar SJ. 2-Methyl-6-phenylethynyl-pyridine (MPSP), a non-competitive mGluR5 antagonist, differentially affects the anticonvulsant activity of four conventional antiepileptic drugs against amygdale-kindled seizures in rats[J]. Pharmacol Rep, 2009,61(4):621-630.
[11] Tang AH, Alger BE. Homer protein-metabotropic glutamate receptor bingding regulates endocannabinoid signaling and affects hyperexcitability in mouse model of fragile X syndrome [J]. J Neurosci, 2015,35(9):3938-3945.
[12] Deutsch SI, Burket JA, Cannon WR, et al. Selective mGluR5 antagonism attenuates the stress-induced reduction of MK-801′s antiseizure potency in the genetically inbred Balb/c mouse[J]. Epilepsy Behav, 2011,21(4):352-355.
[13] Bianchi R, Chuang SC, Zhao W, et al. Cellular plasticity for group-I mGluR-mediated epileptogenesis [J]. J Neurosci, 2009,29(11):3497-3507.
[14] Guo F, Sun F, Yu JL, et al. Abnormal expressions of glutamate transporters and metabotropic glutamate receptor 1 in the spontaneously epileptic rat hippocampus[J]. Brain Res Bull, 2010,81(4-5):510-516.
[15] Sukigara S, Dai H, Nabatame S, et al. Expression of astrocyte-related receptors in cortical dysplasia with intractable epilepsy [J]. J Neuropathol Exp Neurol, 2014,73(8):798-806.
[16] Vijaykumar TS, Nath A, Chauhan A. Chloroquine mediated molecular turning of astrocytes for enhanced permissiveness to HIV infection [J]. Virology, 2008,381(1):1-5.
[17] Bhattacharyya S, Solakyildirim K, Zhang Z, et al. Chloroquine reduces arylsulphatase B activity and increases chondroitin-4-sulphate: implications for mechanisms of action and resistance[J]. Malar J, 2009,8:303.
[18] 張振春,李紅,于守杰,等. 羥基氯喹對(duì)系統(tǒng)性紅斑狼瘡患者血脂水平的影響[J].山東醫(yī)藥,2008,48(20):5.
高平(E-mail: wxh_2005@163.com)
10.3969/j.issn.1002-266X.2017.13.014
R544.1
A
1002-266X(2017)13-0049-03
2017-02-09)
猜你喜歡
中國(guó)民間療法(2021年5期)2021-06-09 09:21:04
中華養(yǎng)生保健(2020年2期)2020-11-16 00:49:00
解放軍醫(yī)學(xué)院學(xué)報(bào)(2020年12期)2020-03-29 05:11:46
中成藥(2017年6期)2017-06-13 07:30:35
飲食科學(xué)(2017年5期)2017-05-20 17:11:53
臨床醫(yī)藥文獻(xiàn)雜志(電子版)(2017年11期)2017-05-17 04:48:10
安徽醫(yī)科大學(xué)學(xué)報(bào)(2015年9期)2015-12-16 11:09:44
中國(guó)當(dāng)代醫(yī)藥(2015年7期)2015-03-01 02:01:13
西南軍醫(yī)(2015年4期)2015-01-23 01:19:30
西部中醫(yī)藥(2014年6期)2014-03-11 16:07:47

- 山東醫(yī)藥的其它文章
- 小劑量硝酸甘油對(duì)老年股骨頸骨折患者全髖關(guān)節(jié)置換術(shù)后血流動(dòng)力學(xué)及血清乳酸水平的影響
- 補(bǔ)硒對(duì)自身免疫性甲狀腺炎患者甲狀腺抗體及T淋巴細(xì)胞亞群的影響
- 經(jīng)腹膜后或經(jīng)腹腔入路腹腔鏡下腎上腺腫瘤切除術(shù)對(duì)單側(cè)腎上腺腫瘤患者腎功能的影響
- 慢性髓性白血病患者骨髓單個(gè)核細(xì)胞SDF-1水平與病情及臨床參數(shù)的關(guān)系
- 女性盆底功能障礙患者診療行為主動(dòng)性的影響因素
- 沙棘水提物對(duì)UVC輻射HaCaT細(xì)胞損傷的保護(hù)作用及其機(jī)制