山東壽光蔬菜種業(yè)集團(tuán)示范園部分番茄品種褪綠病毒感染分析
2017-04-11 07:38:04吳韻雅楊葳劉敏敏楊碩歐陽波
長江蔬菜·學(xué)術(shù)版 2017年1期
吳韻雅+楊葳+劉敏敏+楊碩+歐陽波
摘 要:番茄褪綠病毒(ToCV)病近幾年來在我國各地相繼暴發(fā),北方產(chǎn)區(qū)染病情況尤為嚴(yán)重。對山東壽光蔬菜種業(yè)集團(tuán)示范園內(nèi)采集回的27個(gè)番茄品種的樣品進(jìn)行RT-PCR檢測,結(jié)果表明,其中23個(gè)品種感染了ToCV;之后對這23個(gè)樣品進(jìn)行實(shí)時(shí)熒光定量PCR檢測,分析病毒的CP基因的相對表達(dá)量,結(jié)果表明,感染品種中ToCV含量存在明顯差異;進(jìn)一步對樣品的葉綠素含量進(jìn)行了檢測,結(jié)果表明,較難通過葉綠素含量高低判斷ToCV的感染程度。
關(guān)鍵詞:番茄褪綠病毒;PCR;實(shí)時(shí)熒光定量PCR;葉綠素
番茄是我國和全球最重要的蔬菜作物之一。近年來,番茄褪綠病毒(Tomato Chlorosis Virus,ToCV)病對我國番茄產(chǎn)業(yè)形成了潛在的威脅。2012年,北京首次出現(xiàn)該病毒病發(fā)生的報(bào)道,隨后在膠東半島、天津、河北等地也相繼暴發(fā)了ToCV[1~3],目前該病毒病已經(jīng)在山東省廣泛蔓延,使得番茄產(chǎn)量大幅度下降,造成了巨大的經(jīng)濟(jì)損失。
ToCV屬長線形病毒科(Closteroviridae)毛形病毒屬(Crinivirus),該病毒巨大,由2條超過8 kb的RNA鏈組成。ToCV不能由種子、汁液等途徑進(jìn)行傳播,只能由銀葉粉虱(Bemisia argentifolii)、煙粉虱(B. tabaci)、紋翅粉虱(Trialeurodes abutilonea)、溫室白粉虱(T. vaporariorum)傳播,是迄今為止發(fā)現(xiàn)的唯一一個(gè)可以由2個(gè)屬的粉虱作為傳播媒介的植物病毒[4,5]。在我國,煙粉虱是ToCV的主要傳播媒介。1998年,美國佛羅里達(dá)州報(bào)道發(fā)現(xiàn)ToCV[6]。之后,ToCV相繼在美洲、歐洲、亞洲以及中東等多個(gè)國家和地區(qū)蔓延。ToCV不但能侵染番茄,還可以侵染馬鈴薯、甜椒等其他的園藝作物[7],為害極大。
ToCV在侵染番茄后有較長的潛伏期,一般情況下3~4周內(nèi)不會(huì)表現(xiàn)出明顯癥狀,因此難以發(fā)現(xiàn)植株已經(jīng)染病[8]。而且發(fā)病初期的典型癥狀與缺素癥極其相似,容易混淆,具體表現(xiàn)為葉脈間失綠,葉片變厚,但葉脈仍為綠色;發(fā)病中期,葉片從邊緣向中間壞死;發(fā)病后期,整個(gè)植株病死,嚴(yán)重影響果實(shí)的
質(zhì)量。
目前,對于ToCV的鑒定和檢測,主要運(yùn)用血清學(xué)方法[9]和分子生物學(xué)方法[10,11]。分子生物學(xué)技術(shù)中,PCR技術(shù)操作簡便、靈敏度高,因此被廣泛使用。在PCR技術(shù)上發(fā)展起來的實(shí)時(shí)熒光定量PCR更是可以高效、快速地鑒別包括ToCV在內(nèi)的任何植物病毒[12]。
山東壽光是“中國蔬菜之鄉(xiāng)”,山東壽光蔬菜種業(yè)集團(tuán)示范園是山東有代表性的示范園區(qū)之一。2015年示范園種植有800多個(gè)品種,當(dāng)年的示范種植中許多番茄品種出現(xiàn)了葉片皺縮卷曲、邊緣黃化等明顯的病毒病癥狀,疑似感染了ToCV。為了確認(rèn)ToCV的感染情況,從該示范園中取樣,通過RT-PCR檢測ToCV的外殼蛋白基因CP,分析ToCV的感染情況,進(jìn)一步通過實(shí)時(shí)熒光定量PCR檢測確定染病品種中CP基因的相對表達(dá)量,與此同時(shí)還測定了樣品中葉綠素含量,以分析ToCV感染程度與葉綠素含量之間的關(guān)系。
1 材料與方法
1.1 試驗(yàn) 材料
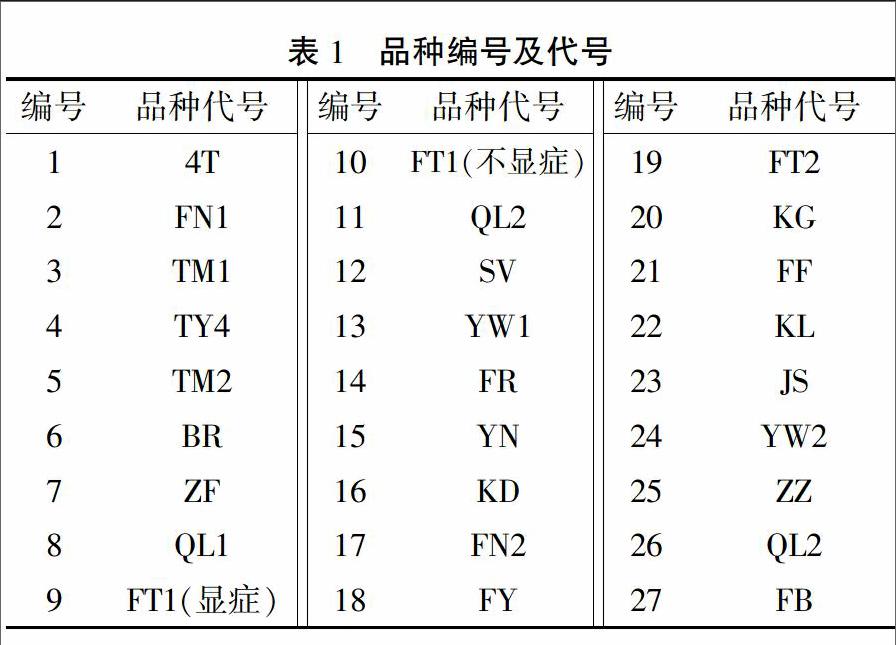
2015年12月6日,從山東壽光市蔬菜種業(yè)集團(tuán)示范園采集27個(gè)番茄品種的樣品,編號(hào)為1~27號(hào)(表1)。其中編號(hào)為2、3、27號(hào)的材料表現(xiàn)較好,無明顯疑似ToCV癥狀,而編號(hào)為1、11、23號(hào)的材料表現(xiàn)較差,其葉片明顯變黃,具有明顯的疑似ToCV癥狀。部分材料癥狀有較為明顯的分離,如編號(hào)為9、10號(hào)的植株。同時(shí)采集了顯癥明顯的9號(hào)和未顯癥狀的樣品10號(hào),全部樣品保存在-80℃冰箱中備用。
1.2 試驗(yàn)方法
①樣品總RNA的提取 采用Trizol法提取樣品的總RNA。樣品在液氮中研磨成粉,用Trizol充分提取之后離心,上清液轉(zhuǎn)移至新離心管后用氯仿充分抽提,再次離心,上清液轉(zhuǎn)移至新離心管后用異丙醇沉淀RNA。得到的RNA沉淀用75%乙醇洗滌,離心、干燥后用無RNA酶的超純水溶解。提取得到的RNA樣品用甲醛變性的1%瓊脂糖凝膠快速電泳,檢測RNA的完整性,用NanoDrop測定RNA濃度。
②RT-PCR擴(kuò)增 用DNase去除RNA樣品中的DNA后,進(jìn)行反轉(zhuǎn)錄(RT)反應(yīng),體系如下(20 μL):RNA樣品2.2 μL、HiScriptII逆轉(zhuǎn)錄酶2 μL、2×緩沖液10 μL、Actin基因的正向和反向引物(10 μmol/L)各0.3 μL、六堿基隨機(jī)引物5.2 μL。RT反應(yīng)在PCR儀上進(jìn)行,程序如下:25℃ 5 min,50℃ 15 min,85℃ 5 min。反轉(zhuǎn)錄完成后得到的cDNA加入40 μL ddH2O稀釋,保存于-20℃?zhèn)溆谩?/p>
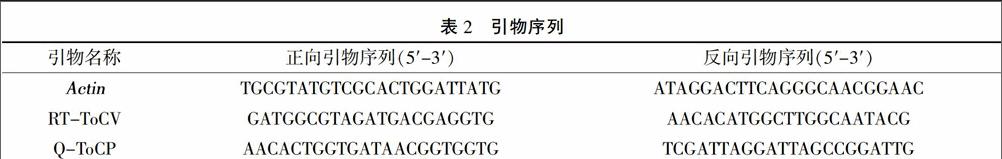
利用Actin基因的引物檢測RT反應(yīng)的質(zhì)量,利用ToCV的CP基因引物RT-ToCV檢測ToCV的存在與否。PCR擴(kuò)增體系如下(15 μL):ddH2O 10.42 μL、10×緩沖液 1.5 μL、dNTPs 1.2 μL、正向和反向引物(10 μmol/L)各 0.4 μL、Taq DNA聚合酶0.08 μL,模板cDNA 1 μL。PCR反應(yīng)程序?yàn)椋?5℃預(yù)變性2 min;95℃變性30 s,65℃退火30 s,72℃延伸 30 s,擴(kuò)增30個(gè)循環(huán);72℃延伸10 min。PCR擴(kuò)增產(chǎn)物進(jìn)行1%瓊脂糖凝膠電泳并利用凝膠成像系統(tǒng)記錄結(jié)果。本研究所用引物序列見表2。
③實(shí)時(shí)熒光定量PCR擴(kuò)增 以上述cDNA作模板,以番茄Actin基因?yàn)閮?nèi)參,利用CP基因的定量PCR引物Q-ToCP進(jìn)行擴(kuò)增,檢測CP基因的相對表達(dá)量。采用羅氏480實(shí)時(shí)熒光定量PCR儀進(jìn)行PCR擴(kuò)增,PCR反應(yīng)體系為(10 μL):2×SYBR Green熒光定量PCR混合液5 μL、正向和反向引物(10 μmol/L)各0.5 μL、cDNA模板 4 μL。定量PCR反應(yīng)程序?yàn)椋?5℃預(yù)變性60 s;95℃ 10 s,57℃ 15 s,72℃ 20 s,共擴(kuò)增40個(gè)循環(huán),每次循環(huán)結(jié)束采集熒光信號(hào);之后進(jìn)行溶解曲線的采集與分析。循環(huán)閾值(Ct)由機(jī)器自動(dòng)讀取。根據(jù)公式REL=Average[power(2,-CtCP)/power(2,-CtActin)]計(jì)算樣品中CP基因的相對表達(dá)量。
④葉綠素含量測定 葉綠素的提取方法如下:取0.1 g經(jīng)液氮研磨的樣品粉末于2 mL的離心管中,加入2 mL 95%丙酮-無水乙醇(2∶1)提取液。將離心管放入25℃生化培養(yǎng)箱中過夜提取直至肉眼觀察到樣品粉末變白為止。取200 μL樣品放入多功能酶標(biāo)儀中,以提取液作為空白對照,測定645、663 nm波長下的吸光度。利用Arnon公式計(jì)算葉綠素含量,葉綠素總含量(mg/g)=(20.29A645+8.05A663)×v/w×1 000。
2 結(jié)果與分析
2.1 RT-PCR檢測ToCV的感染情況
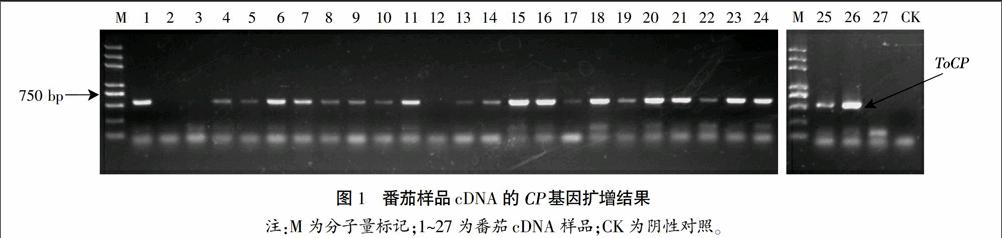
將采集到的27份樣品葉片提取總RNA,反轉(zhuǎn)錄后得到cDNA,利用Actin基因的特異引物擴(kuò)增樣品以檢測cDNA的質(zhì)量,發(fā)現(xiàn)cDNA質(zhì)量符合RT-PCR要求。進(jìn)一步利用CP基因的特異引物進(jìn)行PCR擴(kuò)增,結(jié)果如圖1。
從圖1可以看出,有19個(gè)樣品能擴(kuò)增出CP基因的清晰條帶,4個(gè)樣品的條帶較淺,另外有4個(gè)樣品看不到條帶。從RT-PCR的結(jié)果來看,27個(gè)樣品中有23個(gè)檢測出ToCP,檢出率高達(dá)85%。
2.2 實(shí)時(shí)熒光定量PCR檢測CP基因的相對表達(dá)量
對RT-PCR中擴(kuò)增出條帶的23個(gè)樣品利用實(shí)時(shí)熒光定量PCR進(jìn)一步檢測ToCV病毒的CP基因的相對表達(dá)量,同時(shí)設(shè)置了田間抗性表型良好、RT-PCR中無可見條帶的27號(hào)樣品作為參照系。以Actin為內(nèi)參基因,利用ToCV的CP基因特異引物Q-ToCP-F和Q-ToCP-R對樣品cDNA進(jìn)行定量PCR擴(kuò)增,獲得Ct值,計(jì)算得到CP基因的相對表達(dá)量,結(jié)果見圖2。
從圖2可以看出,CP的相對表達(dá)量在11號(hào)樣品中最高,表明其感染程度最嚴(yán)重。根據(jù)CP基因的表達(dá)量高低排序,其中9、23、20、1、6、18、19、7、21、15、24號(hào)樣品明顯感染了不同程度的ToCV;22號(hào)樣品相對表達(dá)量最低,感染程度最輕;其余樣品CP基因的表達(dá)量均較低,包括作為參照的27號(hào)樣品。
2.3 葉綠素含量
從圖3可以看出,樣品中葉綠素含量差異較大。15號(hào)樣品的葉綠素含量最高,為0.818 mg/g,13號(hào)和4號(hào)樣品次之;1號(hào)樣品的葉綠素含量最低,為0.063 mg/g, 5號(hào)和12號(hào)等樣品的葉綠素含量也很低。結(jié)合圖2、3結(jié)果可看出,僅憑葉綠素含量無法判斷ToCV的感染程度。
3 討論與結(jié)論
ToCV的鑒定技術(shù)主要包括表型鑒定、血清學(xué)鑒定和分子生物學(xué)鑒定,其中實(shí)時(shí)熒光定量PCR技術(shù)[12]和反轉(zhuǎn)錄環(huán)介導(dǎo)等溫?cái)U(kuò)增檢測技術(shù)[3]是目前較為高效、快速、準(zhǔn)確的鑒定方法。本研究從山東壽光蔬菜種業(yè)集團(tuán)示范園采集27個(gè)番茄品種的樣品,通過RT-PCR檢測確定染病品種多達(dá)23個(gè),比率為85%。從這個(gè)結(jié)果可以推測當(dāng)前生產(chǎn)上的絕大部分番茄品種對ToCV是感病的。進(jìn)一步對RT-PCR檢測判斷為明顯染病的品種進(jìn)行實(shí)時(shí)熒光定量PCR,檢測CP基因的相對表達(dá)量,其中有12個(gè)品種CP基因相對表達(dá)量較高,如11、23號(hào)樣品,可以判斷其感染程度較為嚴(yán)重,推斷這些品種對ToCV的抗性較差,在ToCV暴發(fā)的地區(qū)不宜推廣或者要做好嚴(yán)格的防控措施。盡管2、3、12、27號(hào)樣品在RT-PCR檢測中沒有擴(kuò)增出CP基因的條帶,表現(xiàn)為未受ToCV感染,但由于缺乏嚴(yán)格的接種鑒定,尚不能推斷這些品種對ToCV具有抗性。實(shí)際上,以27號(hào)樣品作為參照進(jìn)行實(shí)時(shí)熒光定量PCR檢測時(shí),仍然能夠檢測到熒光信號(hào),即表明仍然有ToCV感染的跡象。從這里也可以看出,實(shí)時(shí)熒光定量PCR比傳統(tǒng)的RT-PCR更為靈敏。總之,對未檢測到明顯ToCV感染的品種,可以進(jìn)一步進(jìn)行多茬多點(diǎn)觀察。
通過測定樣品中葉綠素含量發(fā)現(xiàn),無論是檢測為感染了ToCV還是沒有感染ToCV的品種,葉綠素含量有高有低,表現(xiàn)出明顯的品種差異。同時(shí),感病品種中,較難通過葉綠素含量高低判斷ToCV的感染程度。
在ToCV的防控方面,由于現(xiàn)在沒有抗病商業(yè)品種可以選擇,可以優(yōu)先選擇在發(fā)病條件下表現(xiàn)良好、分子檢測CP基因含量低的耐病品種。同時(shí),對病原的傳播進(jìn)行嚴(yán)格的防控,包括采用防蟲網(wǎng)、粘蟲板和殺蟲劑嚴(yán)防粉虱,嚴(yán)禁從病區(qū)調(diào)運(yùn)種苗等。此外,對疑似感染了ToCV的苗子,及時(shí)進(jìn)行拔除處理,并對生產(chǎn)田進(jìn)行抗病毒防治。
當(dāng)然,防治病蟲害的根本辦法是種植抗性優(yōu)良的品種,并配合正確的田間管理。因此,培育抗番茄褪綠病毒的優(yōu)良品種應(yīng)該作為育種家們未來重點(diǎn)研究的課題。
隨著煙粉虱在全世界范圍內(nèi)的大規(guī)模擴(kuò)散,由煙粉虱傳播的病毒病日趨嚴(yán)重。由煙粉虱傳播的ToCV在世界范圍內(nèi)相繼暴發(fā),也在我國山東省多個(gè)地區(qū)發(fā)生并且造成嚴(yán)重為害。建立嚴(yán)格的防控體系和盡快選育出抗病品種是當(dāng)前重要的任務(wù)。目前,西班牙科學(xué)家已從野生番茄中鑒定出2份抗褪綠病毒的番茄材料[13],盡快將其中的抗性基因轉(zhuǎn)育到栽培品種中是將來一段時(shí)間的重要研發(fā)目標(biāo)。
參考文獻(xiàn)
[1] 高利利,孫國珍,王勇,等.天津地區(qū)番茄褪綠病毒的分子檢測和鑒定[J].華北農(nóng)學(xué)報(bào),2015(3):211-215.
[2] 孫國珍,高利利,陸文利,等.河北省設(shè)施番茄褪綠病毒分子檢測和鑒定研究[J].北方園藝,2015(9):95-98.
[3] 趙黎明,李剛,劉永杰,等.侵染番茄的番茄褪綠病毒山東泰安分離物的分子鑒定和序列分析[J].植物保護(hù),2014
(5):34-39.
[4] Kataya A R A, Stavridou E, Farhan K, et al. Nucleotide sequence analysis and detection of a Greek isolate of tomato chlorosis virus[J]. Plant Pathology, 2008, 57: 819-824.
[5] Wintermantel W M, Wisler G C, Anchieta A G, et al. The complete nucleotide sequence and genome organization of tomato chlorosis virus[J]. Archives of Virology, 2005, 150:
2 287-2 298.
[6] Wisler G C, Li R H, Liu H Y, et al. Tomato chlorosis virus: a new whitefly-transmitted, phloem-limited, bipartite closteroviruses of tomato[J]. Phytopathology, 1998, 88: 402-409.
[7] Freitas D M S, Nardin I, Shimoyama N, et al. First report of tomato chlorosis virus in potato in Brazil[J]. Virology, 2014, 464: 365-374.
[8] Font M I, Juarez M, Martinez O, et al. Current status and newly discovered natural hosts of tomato infectious chlorosis virus and tomato chlorosis virus in Spain[J]. Plant Disease, 2004, 88: 82.
[9] Jacquemond M, Verdin E, Dalmon A, et al. Serological and molecular detection of tomato chlorosis virus and tomato infectious chlorosis virus in tomato[J]. Plant Pathology, 2009, 58: 210-220.
[10] Tiberini A, Tomassoli L, Barba M, et al. Oligonucleotide microarray-based detection and identification of 10 major tomato viruses[J]. Journal of Virological Methods, 2010, 168: 133-140.
[11] Moriones E, Nabas-Castillo J. Tomato yellow leaf curl virus, an emerging virus complex causing epidemics worldwide[J]. Virus Research, 2000, 71: 123-134.
[12] Papayiannis L C, Harkou I S, Markou Y M, et al. Rapid discrimination of tomato chlorosis virus, tomato infectious chlorosis virus and co-amplification of plant internal control using real-time RT-PCR[J]. Journal of Virological Methods, 2011, 176: 53-59.
[13] García-Cano E, Navas-Castillo J, Moriones E, et al. Resistance to tomato chlorosis virus in wild tomato species that impair virus accumulation and disease symptom expression[J]. Phytopathology, 2010, 100: 582-592.
